
Abstract
Objective. To verify the hypothesis that blockade of CD28 costimulation by treatment with abatacept in patients with rheumatoid arthritis (RA) might induce a reduction in the number of CD28– T cells, as well as other effector T cell populations. We evaluated whether these variations correlate with clinical response.
Methods. Peripheral blood T cell subsets were longitudinally evaluated by flow cytometry through the analysis of CD28, CD45RA, and CCR7 expression in 16 patients with RA who were treated with abatacept.
Results. After 48 weeks of treatment, the proportion and the absolute number of circulating CD8+CD28– T cells decreased (p = 0.008, p = 0.055, respectively, compared with baseline), as well as the proportion of the CD8+CD45RA+CCR7– cells, thought to represent terminally differentiated effector T cells (p = 0.03). Reductions of percentages of circulating CD4+CD28– and CD8+CD28– T cells, and (CCR7–) CD8+ total effector T cells were directly correlated with the reduction of Disease Activity Score 28 C-reactive protein (r = 0.58, p = 0.014; r = 0.47, p = 0.059; r = 0.59, p = 0.012, respectively).
Conclusion. After therapy with abatacept, circulating CD28– T cells and other effector populations decrease in patients with RA. This decrease is correlated with clinical response.
- ABATACEPT
- RHEUMATOID ARTHRITIS
- CD28-NEGATIVE T CELLS
- DISEASE ACTIVITY SCORE
- T LYMPHOCYTES
The glycoprotein CD28 is the receptor of costimulatory signals delivered by antigen-presenting cells through the CD80/CD86 family during T cell activation, and thereby delivers the strongest known “second signal” in the activation and survival of T cells by T cell receptor/CD31. Moreover, costimulation through CD28, stabilizing interleukin 2 (IL-2) gene transcription and translation, is critical for IL-2 production by T cells2.
In healthy individuals, CD28 is constitutively expressed by almost all CD4+ and more than 50% of CD8+ T cells. The number of CD28– T cells is extremely low in normal newborns3, and increases gradually with age4.
Many studies have demonstrated a defective expression of CD28 on T cells, as a consequence of the expansion of CD8+CD28– and sometimes also of a small CD4+CD28– population, in clinical conditions characterized by immune activation such as human immunodeficiency virus infection3–8, organ transplant9, acute viral infections10, or autoimmune diseases such as systemic lupus erythematosus11 and rheumatoid arthritis (RA)12. In RA, the expansion of CD28– T cells is associated with aggressive disease, extraarticular manifestations13, and preclinical atherosclerotic changes, including arterial endothelial dysfunction, suggesting that they may contribute to early atherosclerotic damage in these patients14.
It has been shown that downmodulation of CD28 cellular surface marker expression can be obtained with engagement with its ligand15, prolonged stimulation with specific peptide antigens16, or even with cytokines such as IL-417 or IL-218. The CD28– T cell population displays some functional properties of differentiated effector cells18,19. For a phenotypic identification of these populations, it is currently accepted that memory CD4+ and CD8+ T cells are generally CD45RA-negative and comprise at least 2 functionally distinct subsets: (1) nonpolarized “central memory” T cells (TCM), which express the chemokine receptor CCR7 and home to the T cell areas of secondary lymphoid organs; and (2) polarized “effector memory” T cells (TEM), which have lost the expression of CCR7 and have acquired the capacity to migrate to nonlymphoid tissues20–22. Therefore, the evaluation of CCR7 together with CD45RA is now considered the best available way to enumerate TCM and TEM. A third subset of memory T cells expresses CD45RA as do naive cells, but not CCR7. Variously termed, these are highly differentiated cells, with persisting effector functions (“terminally differentiated effector memory”, TTDEM)23.
Abatacept is a fusion protein (CTLA4-Ig) that can act as a costimulation blocker binding to CD80 and CD86 on antigen-presenting cells, blocking the engagement of CD28 on T cells and preventing T cell activation, and it has proven useful in the treatment of patients with RA, including those refractory to tumor necrosis factor-α (TNF-α) blocking agents24,25.
Our aim was to verify that blockade of CD28 costimulation by treatment with abatacept in patients with RA refractory to TNF-α inhibition might induce a reduction in the number of circulating CD28– T cells, as well as of other effector T cell populations, and to evaluate whether these variations are correlated with clinical response.
MATERIALS AND METHODS
Patients
Twenty consecutive patients with a diagnosis of RA, as defined by the 1987 American College of Rheumatology criteria26, who had previously failed 1 or more TNF-blocking therapies, were treated with abatacept by intravenous infusion according to baseline weight (< 60 kg, 500 mg; 60–100 kg, 750 mg; and > 100 kg, 1000 mg) on days 1, 15, 29, and then monthly. The study was conducted in accord with the recommendations of the Helsinki Declaration, all investigations were those routinely required to evaluate the patients, and all patients gave informed consent for all procedures. Clinical assessment of patients was performed using the Disease Activity Score 28-joint count based on C-reactive protein (DAS28), and clinical response was evaluated with the European League Against Rheumatism (EULAR) response criteria27. Main clinical and laboratory data of these patients are summarized in Table 1.
Patient characteristics at baseline. Medians, except for HAQ score, are interquartile range (25th–75th percentile).
Twenty-seven healthy volunteers (median age 49 yrs) were recruited among laboratory personnel as controls.
Leukocyte phenotype analysis
Peripheral blood lymphocyte surface markers were evaluated by 4-color flow cytometry (Cytomics FC-500, Beckman Coulter Inc., Fullerton, CA, USA) at the moment of the first infusion with abatacept (t = 0), and at weeks 4, 12, 24, and 48, using various combinations of monoclonal antibodies (CD3, CD4, CD8, CD45RA, CD45R0, CD28, CCR7; Beckman Coulter or R&D Systems Inc., Minneapolis, MN, USA), to identify naive, TCM, TEM, and TTDEM lymphocyte subpopulations. Absolute cell count was determined by single-platform analysis using Flow-Count beads (Beckman Coulter).
Statistical analysis
Data are expressed as the median (25th–75th percentile, interquartile range). The comparisons were made using the Mann-Whitney test. The variations of cell populations over time were evaluated by standard one-way analysis of variance. Correlations were evaluated by simple linear regression analysis.
RESULTS
Main demographic and clinical data of 20 patients with RA who previously failed TNF-blocking therapy are shown in Table 1. This was a cohort of patients particularly resistant to treatment as demonstrated by the high number of disease-modifying antirheumatic drugs used and because most of them previously received more than 1 TNF blocker, and in 6 cases also rituximab, without success.
One patient discontinued abatacept for elective surgery to treat an osteoporotic fracture after 1 month. Twelve of 19 evaluable patients (62%) achieved a clinical response according to the EULAR criteria (in 6 cases moderate and in 6 cases good response). Three patients discontinued abatacept at 3 months because of inefficacy and were not evaluated in the longitudinal immunophenotypical study. However, their baseline levels of CD28– T cells were not significantly different from other patients.
In this heavily pretreated cohort of 16 evaluable patients, the proportion of CD8+CD28– cells at baseline was higher than in healthy controls of comparable age [38.7% (24.9–62.3) vs 24.7% (15.4–28.7); p = 0.048]. Also, the proportion and the absolute number of CD4+CD28– T cells were higher in the patient group [4.8% (4.1–15.1) vs 3.6% (2.2–4.2); p = 0.029, and 49 cells/μl (32–75) vs 21 (14–35); p = 0.004, respectively].
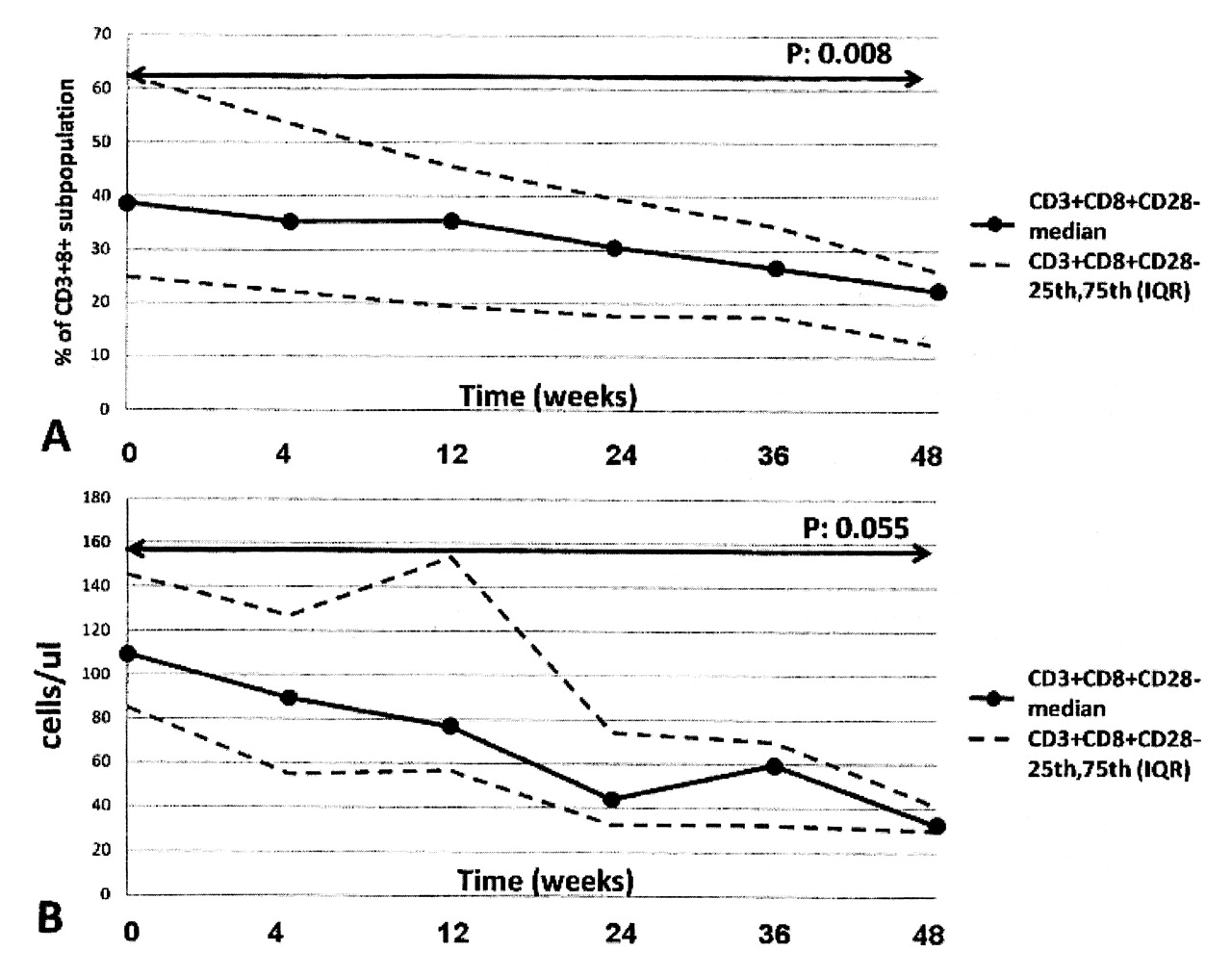
The variations of T cell subsets after treatment with abatacept are shown in Table 2. The proportion of CD8+CD28– T cells progressively decreased with time (p = 0.008 comparing baseline and week 48; Figure 1). Despite a moderate rise of the absolute number of total CD8+ T cells, the absolute number of the CD8+CD28– population also significantly decreased from baseline to week 48 (p = 0.055). Accordingly, the proportion of the CD8+CD45RA+CCR7–, a phenotype thought to represent TTDEM cells, decreased (p = 0.03). Conversely, the proportions of CD8+CD28+ and that of TCM CD8+ cells increased (p = 0.002) from baseline to week 48. There was also a moderate, although not statistically significant, reduction of CD4+CD28– T cells.
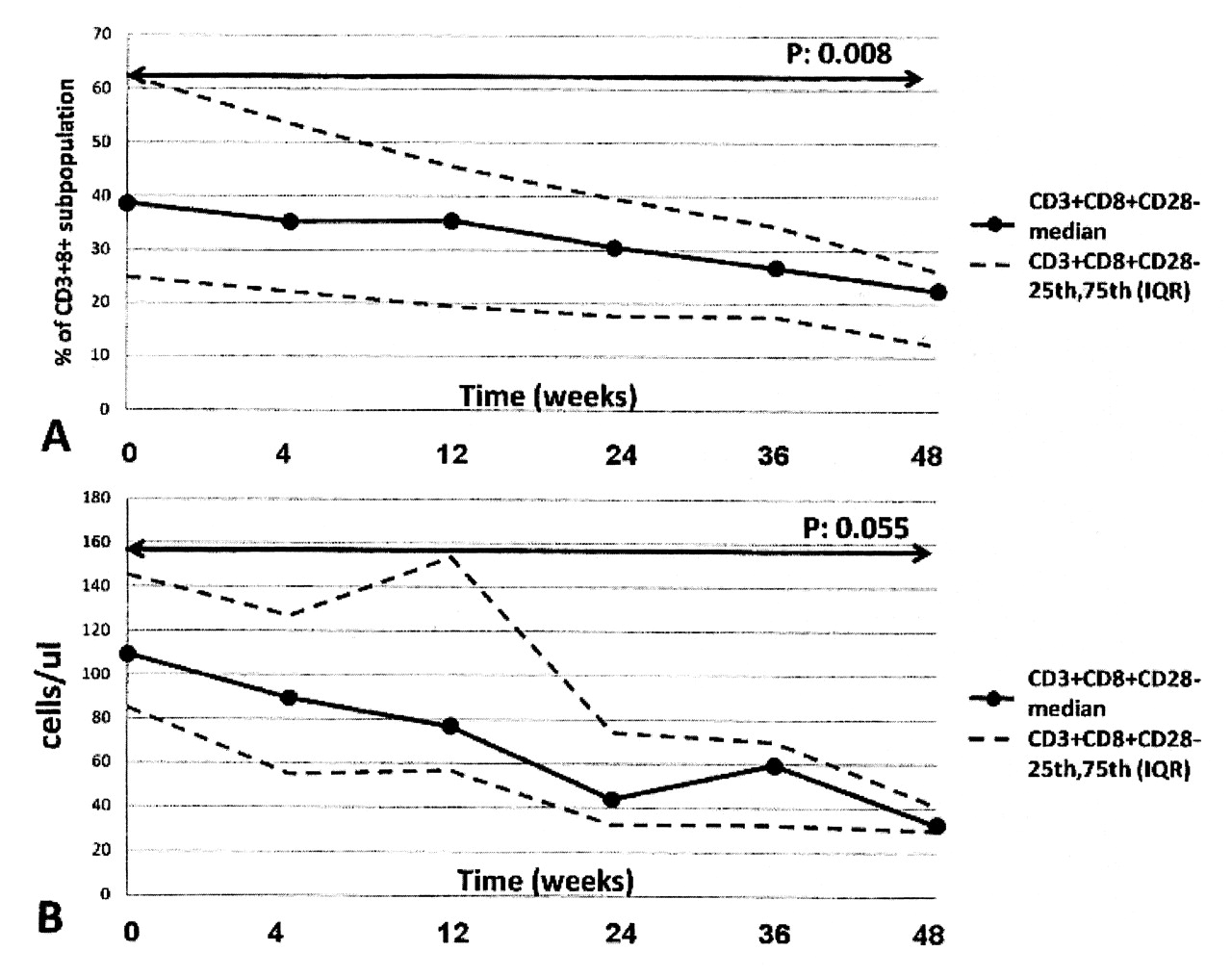
Evolution of percentage and absolute number of CD8+CD28– T cells during therapy with abatacept.
Variations of percentages and absolute numbers of T cell subsets in patients with rheumatoid arthritis after treatment with abatacept. Numbers in 2nd line of each box are absolute numbers (cell/μl).
Interestingly, variations of the T cell subpopulation after therapy with abatacept correlated with the clinical response [variation of the DAS28 (CRP); Figure 2]: in particular, direct correlations were found between DAS28 (CRP) decrease and the reductions of the percentages of circulating CD4+CD28– and CD8+CD28– T cells (r = 0.58, p = 0.014, and r = 0.47, p = 0.059, respectively), and total effector (CCR7–) CD8+ T cells (r = 0.59, p = 0.012).

{kind=link}
{kind=link}
Correlations of variations of T cell subpopulation percentages with variation of the Disease Activity Score (DAS28) during therapy with abatacept.
DISCUSSION
Our main finding was the progressive reduction after treatment with abatacept of the number of circulating CD8+CD28– T cells, a population of differentiated effector lymphocytes that is characteristically raised in patients with RA. Since downmodulation of the costimulatory receptor CD28 follows its ligation by CD80/CD8615, it is conceivable that abatacept, a CTLA4-Ig construct preventing CD28 binding to CD80/CD86, also prevents generation of the CD28– population. The reduction of CD8+ cells with TTDEM phenotype can be easily explained by the fact that this subset largely overlaps with the CD8+CD28– population. The CD4+CD28– population was much smaller than the CD8+ counterpart at baseline, as described by others12–14. A trend toward a reduction of this subset after abatacept therapy was also observed, but it did not reach statistical significance.
Importantly, however, the decreases of CD28– cells among both CD4+ and CD8+ T lymphocyte populations after abatacept therapy were correlated with clinical response as measured by the decrease of the DAS28 (CRP).
To our knowledge, there are no similar data in the literature. It is possible that other therapies in addition to abatacept can induce changes of CD28– T cell populations, particularly TNF-blocking agents. Indeed, downmodulation of CD28 on CD28+ T cells can be obtained in vitro by cultivating them in the presence of TNF-α28, and it has been shown in a small series of patients with RA that treatment with infliximab, a TNF-blocking agent, but not with methotrexate, may decrease the number of circulating CD4+CD28– lymphocytes29, but this decrease was not correlated with clinical response. However, this study evaluated a limited number of patients (n = 7) and reported results concerning only the CD4+CD28– population.
Nevertheless, it is interesting that a study of synovial biopsies after abatacept therapy showed a particular reduction of interferon-γ (IFN-γ) expression that correlated well with improvements of a synovial magnetic resonance index30. IFN-γ is mainly produced by effector CD8+ T cells, and the large majority of this population has a CD28– phenotype. This observation is therefore in keeping with our results, and suggests that one of the mechanisms by which abatacept might obtain its clinical effect is modulation of effector (CD28–) T cell function.
Because studies on chemokine receptor expression on CD4+CD28– T cells in patients with RA showed that these cells have an unusual tissue-trafficking pattern and, despite being end-differentiated effector cells, may be able to survive in lymphoid tissues31, it may be suggested that these cells have a role in manifestations elsewhere than in the joints. Future studies are therefore warranted to evaluate the effect of abatacept in secondary lymphoid organs. Moreover, the influence of this therapy on cardiovascular events in patients with RA should be addressed, because a strict correlation of CD4+CD28– lymphocytes with subclinical atherosclerosis has been observed in these patients14,32. Indeed, the observation that these effector T cells are expanded in patients with unstable angina, where they may invade the atherosclerotic plaque, suggested their direct involvement in plaque disruption33, where they may produce IFN-γ capable of destabilizing the atherosclerotic plaque14,33.
The main limitations of our study, besides the lack of functional data and of tissue analysis, are the small number of patients evaluated and the relatively short period of observation (1 year). However, the observation of a decreased number of circulating CD28– T cells after treatment with abatacept in patients with RA and correlation with clinical response may draw attention to this area and to further research. Research should aim at confirming our observations, and to extending them with longer followup, considering the high longterm safety and efficacy profile of abatacept34.
Acknowledgments
We thank Dr. Fabio Malacarne for advice and suggestions. The skillful technical assistance of Dora Rossi and Carla Bosio is gratefully acknowledged.
Footnotes
- Accepted for publication December 3, 2009.
REFERENCES
- 1.
- 2.
- 3.
- 4.
- 5.
- 6.
- 7.
- 8.
- 9.
- 10.
- 11.
- 12.
- 13.
- 14.
- 15.
- 16.
- 17.
- 18.
- 19.
- 20.
- 21.
- 22.
- 23.
- 24.
- 25.
- 26.
- 27.
- 28.
- 29.
- 30.
- 31.
- 32.
- 33.
- 34.