Article Text

Abstract
Objective: To assess longitudinal expression of a proliferation-inducing ligand (APRIL) in patients with systemic lupus erythematosus (SLE) and its correlation with B lymphocyte stimulator (BLyS) expression, serum anti-dsDNA titres, and clinical disease activity.
Methods: Sixty eight patients with SLE were longitudinally followed up for a median of 369 days. At each visit the physician assessed disease activity by SLEDAI, and blood was collected for determination of serum APRIL and BLyS levels and of blood APRIL and BLyS mRNA levels. Fifteen normal control subjects underwent similar laboratory evaluation.
Results: Dysregulation of APRIL was not as great as that of BLyS. Changes in serum levels of APRIL and BLyS over time were usually discordant, whereas blood levels of APRIL and BLyS mRNA strongly paralleled each other. Serum APRIL levels modestly, but significantly, inversely correlated with serum anti-dsDNA titres in anti-dsDNA positive patients analysed in aggregate. Moreover, serum APRIL levels modestly, but significantly, inversely correlated with clinical disease activity in all patients analysed in aggregate.
Conclusion: Serum levels of APRIL and BLyS are differentially regulated. APRIL may serve as a down modulator of serological and/or clinical autoimmunity in patients with SLE. This may have important ramifications for BLyS targeted treatment, and it remains to be determined whether agents which neutralise only BLyS will be preferable to agents which neutralise both BLyS and APRIL.
- APRIL, a proliferation-inducing ligand
- BLyS, B lymphocyte stimulator
- ELISA, enzyme linked immunosorbent assay
- IFN, interferon
- SLE, systemic lupus erythematosus
- SLEDAI, SLE Disease Activity Index
- TNF, tumour necrosis factor
- B lymphocyte stimulator (BLyS)
- APRIL
- systemic lupus erythematosus
- anti-dsDNA antibodies
- disease activity
Statistics from Altmetric.com
- APRIL, a proliferation-inducing ligand
- BLyS, B lymphocyte stimulator
- ELISA, enzyme linked immunosorbent assay
- IFN, interferon
- SLE, systemic lupus erythematosus
- SLEDAI, SLE Disease Activity Index
- TNF, tumour necrosis factor
- B lymphocyte stimulator (BLyS)
- APRIL
- systemic lupus erythematosus
- anti-dsDNA antibodies
- disease activity
Several members of the tumour necrosis factor (TNF) ligand superfamily have emerged as candidate targets of biological agents for the management of patients with systemic lupus erythematosus (SLE). One such TNF ligand superfamily member is B lymphocyte stimulator (BLyS; trademark of Human Genome Sciences, Inc (HGS), Rockville, MD; also known as BAFF, TALL-1, THANK, TNFSF13B, and zTNF4),1–,6, a potent B cell survival factor.7–,10
Substantial evidence has accumulated in support of BLyS as a therapeutic target in SLE. Constitutive overproduction of BLyS in some (albeit not all) mice transgenic for the human or murine blys gene leads to SLE-like features, including raised titres of multiple autoantibodies (including anti-dsDNA), circulating immune complexes, and renal immunoglobulin deposits.6,11,12 Of great importance, cross sectional and longitudinal studies have documented overexpression of BLyS in >50% of human patients with SLE,13–,15 pointing to a role for BLyS in human disease as well. Moreover, treatment of SLE prone (NZB×NZW)F1 and MRL-lpr/lpr mice with a BLyS antagonist ameliorates progression of disease and improves survival.6,16 Although the efficacy of BLyS antagonism in human patients with SLE remains to be formally proved, a fully human anti-BLyS monoclonal antibody 17 has undergone phase I testing in patients with SLE and has been shown to be safe and biologically active.18 A phase II clinical trial with this anti-BLyS monoclonal antibody in SLE has recently begun.
Another member of the TNF ligand superfamily related to, but distinct from, BLyS is a proliferation-inducing ligand (APRIL, also known as TNFSF13A), a 250 amino acid protein that binds to two of the three BLyS receptors (BCMA and TACI)19–,23 but not to the third BLyS receptor, BAFFR.24,25 The role, if any, for APRIL in SLE pathogenesis is uncertain. In contrast with the development of SLE-like features in blys transgenic mice, april transgenic mice (which constitutively overexpress APRIL) develop neither overt B cell abnormalities nor serological and/or clinical autoimmunity.26 Because APRIL does not bind to BAFFR, the receptor through which BLyS predominantly mediates its agonist effects on B cells,16,24,25,27 it had been felt that APRIL was relatively inert in its effect on B cell biology. However, this belief needs to be modified, and APRIL may importantly contribute to B cell homoeostasis. On the one hand, APRIL and BLyS can each independently promote T cell independent class switching of IgD+ B cells in vitro,28 and surface expression of BCMA by in vitro generated plasma blasts renders them sensitive to agonist effects of both APRIL and BLyS as indicated by their survival and immunoglobulin secretion.29 Accordingly, APRIL could potentiate and/or supplement BLyS activity. On the other hand, TACI (which is bound by both BLyS and APRIL) can transmit to the B cell a negative signal that is vital for the prevention of B cell based autoimmunity.30 Accordingly, APRIL could serve as a homoeostatic down modulator of B cell hyperactivity.
Given the potential for APRIL to up modulate or down modulate the “autoimmunogenic” effects of BLyS, it stands to reason that changes in circulating APRIL levels over time could have important ramifications for serological and/or clinical autoimmunity in patients with SLE. We previously characterised BLyS overexpression in a cohort of 68 patients with SLE followed up longitudinally over 12 months.15 In this report, we examine APRIL expression in these patients.
PATIENTS AND METHODS
Subjects
The selection criteria, characteristics, and medical management of the 68 patients with SLE and 15 healthy control volunteers who served as the subjects of this report have been previously detailed.15 Clinical disease activity was assessed by SLEDAI (SLE Disease Activity Index).31 Drugs being taken were recorded, and venous blood samples were collected for laboratory based studies (see below). Doses of corticosteroids other than prednisone were converted to and reported as daily prednisone equivalent dosages. Each of the patients with SLE who were longitudinally studied for 147–420 (median 369) days was seen 3–12 (median 8) times. Day 0 was defined as the day that a given recruited patient or control underwent his/her first clinical and/or laboratory evaluation.
Serum BLyS determination
Serum BLyS levels for the patients and controls were determined by enzyme linked immunosorbent assay (ELISA)14,32 and have recently been reported.15
Serum APRIL determination
Serum APRIL levels were determined by ELISA.33 To eliminate day to day interassay variability, all serum samples from a given subject were stored at −70°C until the subject’s entire observation period was complete. All serum samples collected over the entire observation period from a given subject were assayed on the same day.
The “normal” range for serum APRIL levels was defined as the geometric mean of all the values (n = 86) ±2SD obtained over time from the 15 control subjects. Each patient with SLE was assigned a serum APRIL phenotype based on his/her serum APRIL levels over time. A “persistently raised” serum APRIL phenotype was assigned when all or all but one of the patient’s serum APRIL levels were greater than the upper limit of normal. An “intermittently raised” serum APRIL phenotype was assigned when at least two of the patient’s serum APRIL levels were greater than the upper limit of normal and at least two were in the normal range. A “persistently normal” serum APRIL phenotype was assigned when no more than one of the patient’s serum APRIL levels was greater than the upper limit of normal. A “persistently low” serum APRIL phenotype was assigned when all or all but one of the patient’s serum APRIL levels were less than the lower limit of normal.
Blood BLyS mRNA determination
Levels of BLyS mRNA in whole blood were determined by real time quantitative polymerase chain reaction and have recently been reported.15
Blood APRIL mRNA determination
Levels of APRIL mRNA were determined by real time quantitative polymerase chain reaction in a manner identical to that used for BLyS mRNA determination, using the oligonucleotide 5′-AAGCAGCACTCTGTCCTGCACCTGGTT-3′ as probe and the primers 5′-GCAGTGCTCACCCAAAAACA-3′ and 5′-CCTTGGAGGTGGCGTTAATG-3′. The APRIL amplicon was designed to span the region containing nucleotides 313–385 of the APRIL coding sequence. The comparative Delta Ct method (Perkin-Elmer user bulletin No 2 1997) was used for measurement of mRNA using an 18S ribosomal RNA probe as the endogenous reference. As with serum APRIL determination, all RNA samples from a given subject were stored at −70°C until the subject’s entire observation period was complete, at which time all the collected samples for that subject were processed in parallel. Results are presented as APRIL mRNA: 18S mRNA ratios.
The “normal” range for blood APRIL mRNA levels and assignment of blood APRIL mRNA phenotypes were determined in a manner analogous to that described above for serum APRIL.
Statistical analysis
All analyses were performed using SigmaStat software (SPSS, Chicago, IL). Serum BLyS and serum APRIL levels were log transformed to achieve normality, permitting parametric testing. Even after log transformation, blood BLyS and APRIL mRNA levels were not normally distributed, so non-parametric testing was performed. Parametric testing between two matched groups was performed by the paired t test. Nominal data were analysed by χ2 analysis of contingency tables or by the Fisher exact test. Correlations were determined by Spearman rank order correlation for interval data that did not display normality and for ordinal data.
RESULTS
Serum APRIL levels and relation to serum BLyS levels
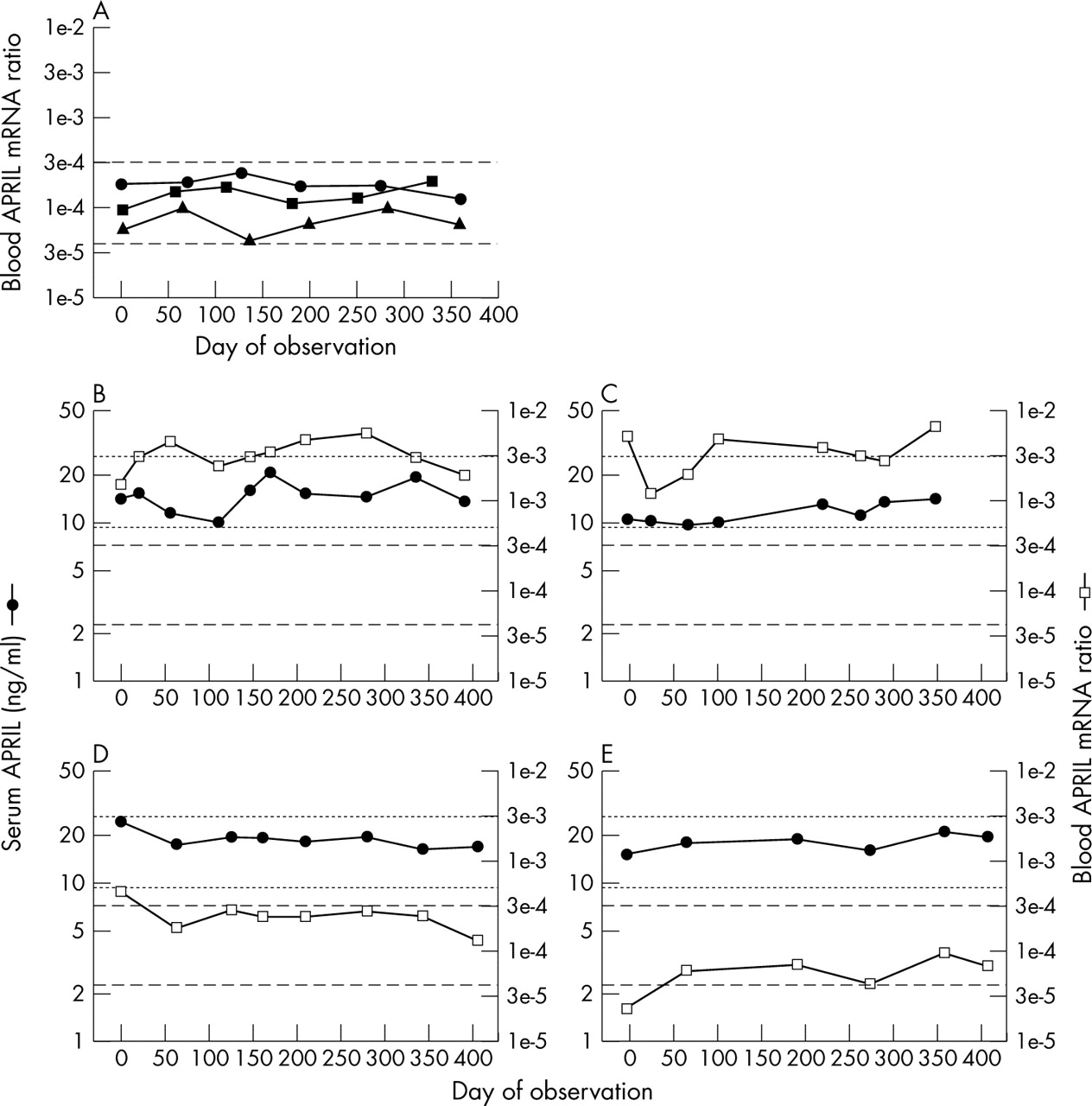
Among normal healthy controls, serum APRIL levels fell within about a threefold range and remained stable over time for any individual subject (fig 1A⇓). This narrow range and stability of serum APRIL levels among normal subjects qualitatively paralleled that of serum BLyS levels, but APRIL levels routinely exceeded those of BLyS by 5- to 10-fold (figs 2A⇓ and B). All but one of the control subjects exhibited a “persistently normal” serum APRIL phenotype, with the remaining control subject having a “persistently raised” serum APRIL phenotype (table 1⇓).
APRIL protein and mRNA phenotypes

Heterogeneity in serum APRIL phenotypes among patients with SLE and relation to disease activity. (A) Serum samples collected from three representative normal controls (circles, triangles, squares) were assayed for APRIL levels. By definition, the first serum samples were collected on day 0. The dashed lines indicate 2SD above and below the geometric mean of all the values (n = 86) obtained from the 15 control subjects and arbitrarily define the “normal” range. (B-E) Serum samples collected from individual patients with SLE were assayed for APRIL levels (circles). Disease activity was measured by SLEDAI (bars). Results for representative patients exhibiting the “persistently normal” serum APRIL phenotype (B), the “persistently low” serum APRIL phenotype (C), the “intermittently raised” serum APRIL phenotype (D), or the “persistently raised” serum APRIL phenotype (E) are illustrated. (F) Serum APRIL levels plotted against the corresponding SLEDAI scores for the 523 individual serum samples from the 68 patients with SLE for whom serum APRIL levels and SLEDAI scores were both determined.

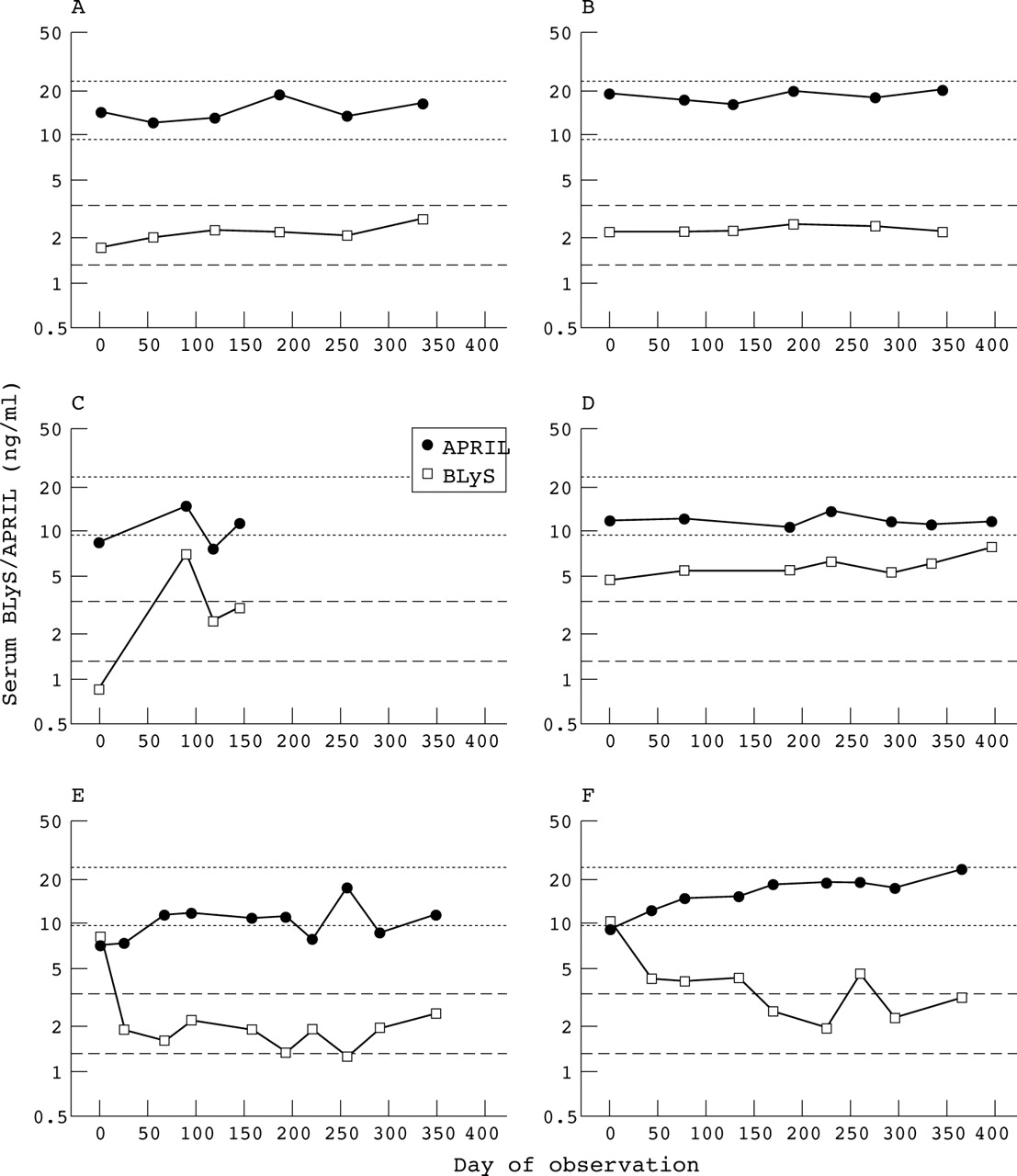
Relation between serum APRIL and serum BLyS levels. Serum samples collected from two representative normal controls (A, B) and four representative patients with SLE (C–F) were assayed for APRIL and BLyS levels. The dotted lines delineate the normal range for serum APRIL, and the dashed lines delineate the normal range for serum BLyS.15
Among patients with SLE, there was some heterogeneity in the serum APRIL phenotype, with “persistently normal”, “persistently low”, “intermittently raised”, and “persistently raised” phenotypes all being represented (figs 1B⇑–E). However, the distribution of serum APRIL phenotypes among patients with SLE did not significantly differ from that among the normal controls (p = 0.415, table 1⇑). In aggregate, APRIL levels were raised in only 28 (5.3%) of the 529 individual serum samples collected from the 68 patients with SLE in comparison with 5 (5.8%) of the 86 individual sera collected from the 15 control subjects (p = 0.953). These “normal” results for serum APRIL levels in the patients with SLE are in sharp contrast with the common increase in serum BLyS levels in these same patients.15
As a rule, there was non-concordance in the patients with SLE between serum APRIL levels and serum BLyS levels. Although serum levels of BLyS and APRIL paralleled each other to a considerable degree in a minority of patients with SLE (figs 2C⇑ and D), changes in serum BLyS levels were discordant with those in serum APRIL levels in the majority of patients (figs 2E⇑ and F). This resulted in a lack of overall correlation between serum levels of APRIL and BLyS (data not shown). Further highlighting the discordance between serum APRIL levels and serum BLyS levels are the responses in the nine patients with SLE treated with short courses of high dose corticosteroids.15 Whereas the decline in serum BLyS levels was dramatic (geometric mean change from 6.9 to 2.6 ng/ml (62% decrease), p<0.001), serum APRIL levels were not appreciably affected in these same patients (geometric mean change from 11.1 to 11.9 ng/ml (−7% decrease), p = 0.412; fig 3⇓).

Insensitivity of serum APRIL levels to treatment with high dose corticosteroids. Nine patients with SLE were treated with short courses of high dose corticosteroids.15 BLyS levels in pretreatment (B-pre) and post-treatment (B-post) serum samples and APRIL levels in pretreatment (A-pre) and post-treatment (A-post) sera are plotted.
Blood APRIL mRNA levels and relation to blood BLyS mRNA levels
The serological results suggested that dysregulation of APRIL among patients with SLE was, at most, modest. To independently assess APRIL expression in patients with SLE by non-serological means, peripheral blood APRIL mRNA levels were measured. In normal controls, blood APRIL mRNA levels showed somewhat greater variations over time, with a broader range of values in comparison with those seen for serum APRIL levels (compare fig 4A⇓ with fig 1A⇑). None of the 88 APRIL mRNA determinations in control blood samples were raised, and all control subjects exhibited a “persistently normal” blood APRIL mRNA phenotype (table 1⇑).

Heterogeneity in blood APRIL mRNA phenotypes among patients with SLE. (A) Peripheral blood samples collected from the same three representative normal controls illustrated in fig 1A⇑ (circles, triangles, squares) were assayed for APRIL mRNA levels. By definition, the first blood samples were collected on day 0. The dashed lines indicate 2SD above and below the geometric mean of all the values (n = 88) obtained from the 15 control subjects and arbitrarily define the “normal” range. (B–E) Serum APRIL levels (filled circles) and blood APRIL mRNA levels (open squares) were measured. The dotted lines delineate the normal range for serum APRIL, and the dashed lines delineate the normal range for blood APRIL mRNA. In four patients, blood APRIL mRNA levels were markedly raised despite normal serum APRIL levels (B, C). In most patients, blood APRIL mRNA levels remained normal throughout the entire observation periods (D, E).
In the patients with SLE, blood APRIL mRNA levels were sometimes raised despite normal serum APRIL levels (figs 4B⇑ and C), although blood APRIL mRNA levels were persistently normal in the majority of patients (figs 4D⇑ and E). Of the 526 individual SLE blood samples assayed for APRIL mRNA, 92 (17.5%) had raised levels (p<0.001 compared with normal controls). This is reflected in the moderate prevalence of “intermittently raised” and “persistently raised” blood APRIL mRNA phenotypes among patients with SLE (p = 0.018 compared with normal controls; table 1⇑).
In contrast with the frequent discordance between serum levels of APRIL and BLyS in patients with SLE (fig 2⇑), there routinely was a strong concordance between blood levels of APRIL and BLyS mRNA not just in normal controls (figs 5A⇓ and B) but in patients with SLE as well (figs 5C⇓ and D). This resulted in a very strong and highly significant correlation between blood APRIL mRNA levels and blood BLyS mRNA levels when taking all the SLE blood samples into account (fig 5E⇓).

Relation between blood APRIL mRNA levels and blood BLyS mRNA levels. (A–D) Blood collected from two representative normal controls (A, B) and two representative patients with SLE (C, D) were assayed for APRIL mRNA and BLyS mRNA. The dotted lines delineate the normal range for blood APRIL mRNA, and the dashed lines delineate the normal range for blood BLyS mRNA.15 (E) Blood BLyS mRNA levels plotted against blood APRIL mRNA levels for the 526 individual blood samples obtained from the 68 patients with SLE for whom both blood APRIL and BLyS mRNA levels were determined.
Inverse correlation between serum APRIL levels and serum anti-dsDNA titres or disease activity
Although circulating BLyS levels do not correlate well with disease activity in individual patients with SLE,15 they do correlate with disease activity and with circulating anti-dsDNA levels among patients with SLE analysed in aggregate.13–15,34 Thus, the prevalent discordance between serum BLyS levels and serum APRIL levels (fig 2⇑) raises the possibility of an inverse relation between serum APRIL levels and BLyS driven biological and/or clinical sequelae.
This was indeed the case. Thirty four patients with SLE harboured a serum anti-dsDNA titre of ⩾1/20 on ⩾2 observational time points. Although serum APRIL levels and serum anti-dsDNA titres substantially paralleled each other in 10 of these patients (figs 6A⇓ and B), an anti-parallel relation was found in the remaining 24 patients (figs 6C⇓ and D). When serum samples from all times from all patients were considered, a modest but significant inverse correlation between serum APRIL levels and serum anti-dsDNA titres was found (fig 6E⇓).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Relation between serum APRIL levels and serum anti-dsDNA titres. (A, B) Serum APRIL levels (filled circles) and serum anti-dsDNA titres (open squares) were determined in two representative patients with SLE in whom changes in serum APRIL levels largely paralleled those in serum anti-dsDNA titres. (C, D) Serum APRIL levels and serum anti-dsDNA titres were determined in two representative patients with SLE in whom changes in serum APRIL levels were largely anti-parallel to those in serum anti-dsDNA titres. (E) Serum anti-dsDNA titres plotted against serum APRIL levels for the 275 individual serum samples obtained from the 34 anti-dsDNA positive patients with SLE.
A similar inverse correlation was found between serum APRIL levels and disease activity (as measured by SLEDAI). Although serum APRIL levels did not overtly correlate with disease activity over time in individual patients with SLE regardless of their serum APRIL phenotype (figs 1B⇑–E), disease activity was modestly, but significantly, inversely correlated with serum APRIL levels when all patient serum samples at all times were collectively analysed (fig 1F⇑). No significant correlations between blood APRIL mRNA levels and disease activity or serum anti-dsDNA titres were found (data not shown).
DISCUSSION
Little information has hitherto been available about circulating APRIL levels in patients with SLE. We now demonstrate that in comparison with the great stability of serum APRIL levels over time in normal donors, serum APRIL levels manifest a limited degree of fluctuation over time in patients with SLE (figs 1A⇑–E). This fluctuation in serum APRIL levels in patients with SLE largely occurs within the normal range and results in a spectrum of serum APRIL phenotypes that is not significantly different from that in normal subjects (table 1⇑). Nevertheless, two facets of the APRIL response are striking.
Firstly, the usual non-concordance between serum levels of APRIL and BLyS (figs 2E⇑ and F), highlighted by the insensitivity of serum APRIL levels to short courses of high dose corticosteroids despite the exquisite sensitivity of serum BLyS levels to such treatment (fig 3⇑), strongly suggests that APRIL and BLyS are differentially regulated. Although in vitro treatment of dendritic cells (and, probably, other myeloid lineage cells) with interferon (IFN)α, IFNγ, or CD40 ligand (CD40L) results in up regulation of both APRIL and BLyS,28 and although blood APRIL and BLyS mRNA levels did strongly correlate with each other in the patients with SLE (fig 5⇑), APRIL is expressed in vivo by other cell types, including cells from prostate, colon, and pancreas.19 It is entirely plausible that the bulk of APRIL produced in vivo arises from sources other than myeloid lineage cells. The “non-myeloid APRIL” may be regulated in a manner different from that of “myeloid APRIL”. Alternatively, it is possible that factors other than IFNα, IFNγ, or CD40L are major post-transcriptional regulators of APRIL and/or BLyS in vivo. These as yet unidentified factors may have differential effects on APRIL and BLyS. Either possibility could explain why the degree of dysregulation in patients with SLE of APRIL is quantitatively much more limited than that of BLyS (compare table 1⇑ with our previous study15).
Secondly, changes in serum APRIL levels in patients with SLE, despite often remaining in the “normal” range, appear to be biologically and clinically important. APRIL binds to TACI,20–,23 which (presumably) delivers a negative signal to the B cell.30 APRIL, however, does not bind to BAFFR, so it does not deliver a BAFFR mediated positive signal to the B cell.16,24,25,27 Because circulating levels of APRIL are considerably greater than those of BLyS (fig 2⇑), even small changes in serum APRIL levels may have substantial ramifications for B cell biology. Serum APRIL levels and serum anti-dsDNA titres usually demonstrated an anti-parallel relation (figs 6C⇑ and D), and a modest inverse correlation between the two was found when all samples from all the anti-dsDNA positive patients were considered (fig 6E⇑). Importantly, despite serum APRIL levels not overtly correlating with disease activity over time for any individual patient with SLE (figs 1B⇑–E), a modest inverse correlation was seen when all patients at all times were considered (fig 1F⇑). Although purely speculative, it may be that the polymorphism of the april gene associated with human SLE35 imparts upon APRIL a reduced biological half life and/or decreased binding affinity to TACI, either of which would reduce the negative regulatory input to B cells. The fact that the inverse correlations between serum APRIL levels and serum anti-dsDNA titres or disease activity are only modest demonstrates that the precise role for APRIL in regulation of autoantibody production and/or disease activity requires further elucidation.
Circulating soluble APRIL may not be the only form of APRIL relevant to B cell biology and/or B cell based autoimmunity. A biologically active, naturally occurring hybrid protein composed of the cytoplasmic and transmembrane domains of TWEAK (another member of the TNF ligand superfamily) fused to the APRIL C-terminal domain is expressed by T cells and monocytes.36 What regulatory role, if any, this cell surface APRIL hybrid plays in vivo remains speculative. There is no a priori reason to exclude the possibility of the APRIL moiety interacting with B cell surface TACI,20–,23 which could transmit a negative signal to the B cell,30 and/or with B cell surface BCMA,20,21,23 which could transmit a positive signal to plasma blasts.29 In addition, alternate splice forms of APRIL have been identified (37 and HGS, unpublished observations). Whether these alternate splice forms result in variants of APRIL and what, if any, biological activities these putative APRIL variants have, remains to be determined. Further investigation into these issues is warranted.
The apparent down regulatory property of APRIL may have important ramifications for BLyS targeted treatment in SLE. It is tempting to speculate that agents which can neutralise only BLyS while having no effect on APRIL may be preferable to those agents which can neutralise both BLyS and APRIL. Ultimately, clinical experience will determine which BLyS antagonists are most efficacious in the treatment of SLE and other B cell based autoimmune disorders.
Acknowledgments
The authors thank Dong Xu for handling and cataloguing the patient samples and all the subjects for their participation in the study.
This work was supported in part by grants from the NIH (AR41006) and the Alliance for Lupus Research and a gift from Lupus International.