Article Text

Abstract
Objectives: The aim of the current study was to determine the contribution of interleukin (IL)1 gene cluster polymorphisms previously implicated in susceptibility for ankylosing spondylitis (AS) to AS susceptibility in different populations worldwide.
Methods: Nine polymorphisms in the IL1 gene cluster members IL1A (rs2856836, rs17561 and rs1894399), IL1B (rs16944), IL1F10 (rs3811058) and IL1RN (rs419598, the IL1RA VNTR, rs315952 and rs315951) were genotyped in 2675 AS cases and 2592 healthy controls recruited in 12 different centres in 10 countries. Association of variants with AS was tested by Mantel–Haenszel random effects analysis.
Results: Strong association was observed with three single nucleotide polymorphisms (SNPs) in the IL1A gene (rs2856836, rs17561, rs1894399, p = 0.0036, 0.000019 and 0.0003, respectively). There was no evidence of significant heterogeneity of effects between centres, and no evidence of non-combinability of findings. The population attributable risk fraction of these variants in Caucasians is estimated at 4–6%.
Conclusions: This study confirms that IL1A is associated with susceptibility to AS. Association of the other IL1 gene complex members could not be excluded in specific populations. Prospective meta-analysis is a useful tool in confirmation studies of genes associated with complex genetic disorders such as AS, providing sufficiently large sample sizes to produce robust findings often not achieved in smaller individual cohorts.
Statistics from Altmetric.com
Association of members of the interleukin (IL)1 gene cluster on chromosome 2q13 has been reported in several studies in Caucasian and Asian populations. The IL1 gene cluster is a 360-kb region containing nine genes with sequence homology either to the pro-inflammatory IL1 agonists IL1A and IL1B, or the anti-inflammatory IL1 antagonist IL1RN (encoding for the protein IL1Ra). Six genes with structural homology to IL1A/B or IL1RN lie between IL1A and IL1RN. They are named IL1F5, IL1F6, IL1F7, IL1F8, IL1F9 and IL1F10. Initial reports focused on association of alleles of the IL1RN variable number tandem repeat (VNTR) with ankylosing spondylitis (AS).1 2 These findings were extended by Maksymowych et al, who demonstrated association between single nucleotide polymorphisms (SNPs) and SNP haplotypes within IL1RN, and AS.3 The same haplotypic associations have also been replicated in Taiwanese AS.4 A study of all IL1 gene cluster members in British Caucasians revealed more widespread association, with evidence of two associated haplotypes tagged by SNPs in IL1B and IL1F10.5 The attributable risk fraction from the locus was 18%, compared with the major histocompatability complex (MHC) that contributes 37–50% of the heritability of AS.6 7 Further mapping in an extended cohort of British Caucasian samples suggested that the strongest associated gene was IL1A.8 The IL1A association has been confirmed in a multi-centre Canadian AS study,9 and also in a Canadian psoriatic arthritis cohort.10 In the current study we sought to determine the contribution of this cluster to AS susceptibility in different populations worldwide by planned meta-analysis.
MATERIALS AND METHODS
A total of 2675 AS cases and 2592 ethnically-matched controls from 12 discreet cohorts in 10 countries were genotyped in this study. The population or city of origin of the cohorts includes The Azores, Canada (Alberta, Toronto, Newfoundland), China (Shanghai), Finland, Hungary, Mexico, Sardinia, Taiwan, UK and USA. All AS cases fulfilled the modified New York criteria,11 as assessed by a local qualified rheumatologist. All cohorts had been recruited with the approval of their local ethics committees.
Nine markers were chosen to genotype across the IL1 gene cluster in all populations (table 1). The markers were chosen based on previous known associations with susceptibility to AS,5 12 and ease of genotyping by PCR and restriction fragment length polymorphism (RFLP). A standard genotyping protocol was generated and distributed to all interested centres utilising PCR-RFLP. Each centre genotyped the markers either using this protocol, or using protocols they had developed prior to the meta-analysis (table 1). Known genotype control samples were provided to each genotyping centre with which to validate the genotyping protocol employed. In the case of the Canadian and Taiwanese cohorts, all the genotypes reported here have previously been reported elsewhere.4 12 In the case of the families used in UK cohort, only the findings for SNPs rs419598, rs315952 and the IL1RN-VNTR have previously been published.5
Two centres (Finland and UK) genotyped AS family populations. In order to combine this data in the case–control meta-analysis, affected family-based controls (AFBAC) were generated; these provide an unbiased estimate of the overall population marker alleles using the non-transmitted parental allele.13 For ease of analysis of the rare alleles of IL1RN-VNTR, this marker was coded as a bi-allelic polymorphism, with allele 1 carrying four repeats, and the combination of the remaining alleles pooled as allele 2.
Marker genotypes in case–control cohorts were checked for Hardy–Weinberg equilibrium by χ2 1 degree of freedom tests, and in families for marker-typing inconsistencies using the “Pedcheck” software.14 Within-population associations were assessed using standard χ2 methods. Meta-analysis of association was assessed using the Mantel–Haenszel method to investigate the combination or interaction of a group of independent studies, providing a random-effects pooled odds ratio and related χ2 and p values.15 The I2 statistic was used to measure the percentage of variation across cohorts due to heterogeneity rather than chance,16 and the Cochrane Q statistic used to test non-combinability of studies. Population attributable risk fraction was calculated using the method of Miettinen.17
RESULTS
The findings in individual populations are presented in table 2 along with the combined Mantel–Haenszel meta-analysis. Not all centres were able to provide genotype data for all polymorphisms, and therefore the number of alleles available for individual markers varies, with between 2195–5349 case alleles available (mean 3709) and between 2186–5183 control alleles (mean 3572). All markers were in Hardy–Weinberg equilibrium in all cohorts, and marker-typing inconsistencies were excluded in the Finnish and UK family data.
Association was observed with each polymorphism in at least one centre, but no variant was invariably associated in all centres. Mantel–Haenszel analysis demonstrated strong association of three SNPs in IL1A with AS (rs2856836, rs17561, rs1894399, p = 0.0036, 0.000019 and 0.0003, respectively). These findings remain statistically significant following correction for the number of variants genotyped (9). These associations were mainly driven by findings from the two largest cohorts (UK affected family-based controls and North American Spondylarthritis Consortium samples). However, most datasets showed at least some support for the association of these variants with disease (fig 1). Further, the I2 statistic was 0% and the Cochrane Q statistic was non-significant for each SNP, indicating that a low proportion of the association was due to heterogeneity, and there was no evidence of non-combinability of findings. The case and control allele frequencies in the US and UK cohorts were strikingly similar for these three SNPs, with no more than 1% difference in minor allele frequencies with the exception of the case allele frequencies for rs2856836. No other marker achieved nominal statistical significance (p<0.05), although marker rs3811058 in the gene IL1F10 nearly achieved significance (p = 0.08).

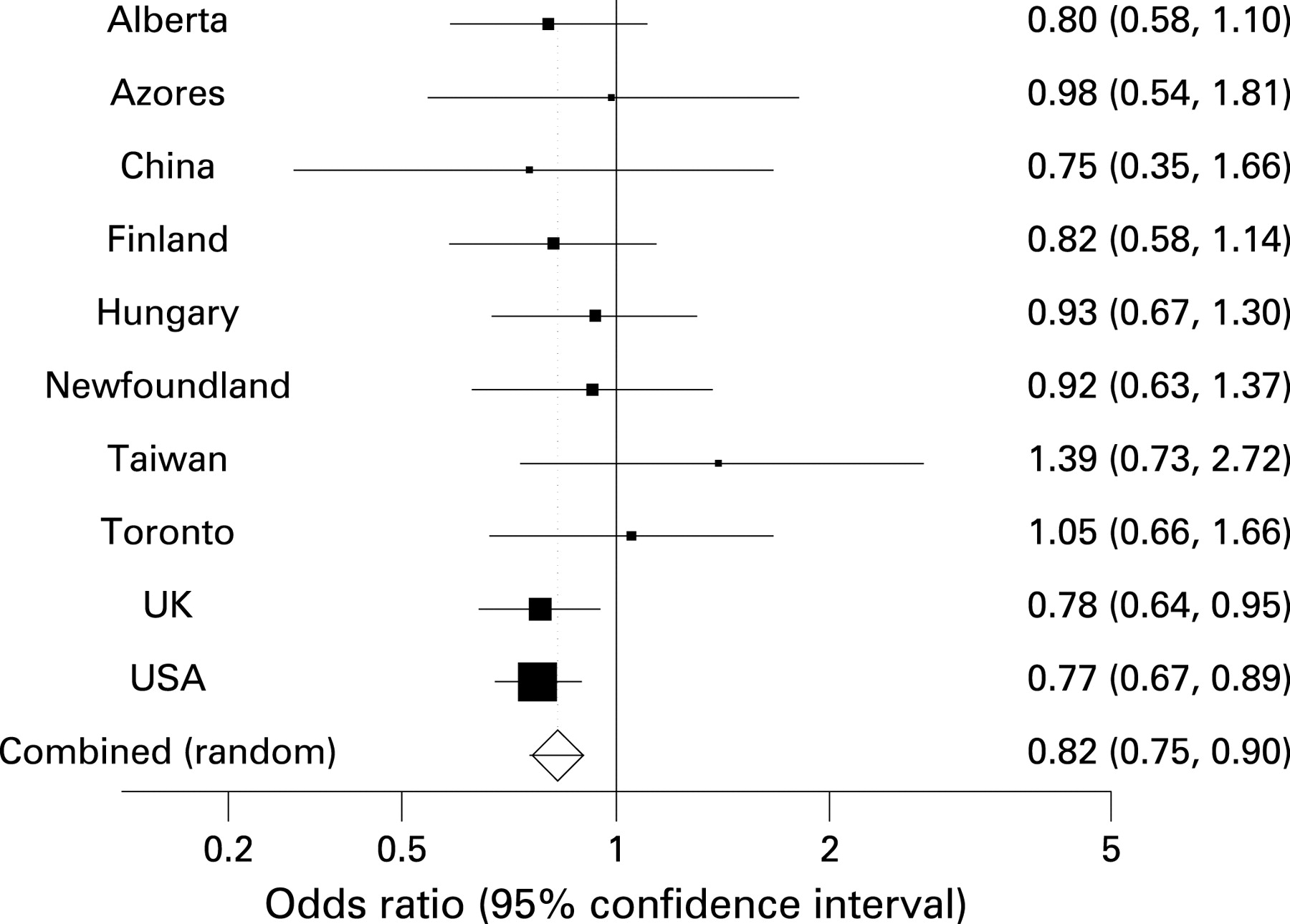{kind=link}
Odds ratios for each polymorphism were quite modest. For the three associated IL1A SNPs, the overall odds ratios were for rs2856836 1.14, rs17561 0.82 and rs1894399 1.2. Assuming a minor allele frequency of 0.3 (roughly the frequency for these three SNPs in Caucasians), the attributable risk fractions for these three markers are 4%, 6% and 6% respectively. The study power to detect an association with AS for a marker with minor allele frequency of 0.3 at a p value of <0.05 and linkage disequilibrium with the true associated variant of D′ = 0.8 for the mean number of case and control samples available is 80% for an odds ratio of >1.2, assuming homogeneity between cohorts.
DISCUSSION
This study confirms previous reports that IL1A genetic variants are associated with AS. The strength of association is considerable (p values for IL1A variants 0.0036 to 0.000019), but the magnitude of the genetic risk associated with each variant is small (attributable risk estimated at 4–6%). The true attributable risk fraction may be greater than this because the markers studied may only be tagging the true disease-associated variant(s), because the odds ratios used in the estimate were derived from diverse ethnic groups, and the association of IL1A with AS may not be homogenous across all ethnicities.
Whilst at least one group showed association with each of the other polymorphisms studied, no other marker showed association in the overall dataset. The power of the individual cohorts was not sufficient to exclude private associations in particular ethnic groups, and it is possible that IL1 gene cluster members other than IL1A are associated with AS in individual ethnic groups. The study did not screen the IL1 gene cluster comprehensively, and thus any associated variants not in significant linkage disequilibrium with the markers genotyped will not have been detected.
This study has the largest sample size of any genetic study yet performed in AS. Its power overall is good to detect effects of the size observed with IL1A, assuming a homogenous effect in different cohorts. In other autoimmune diseases, such as type 1 diabetes, rheumatoid arthritis and Crohn disease, the effect size of associated genes has been similar to those observed with IL1A in AS. Thus, meaningful studies aiming to replicate such findings will require sample sizes similar to this study. Inadequate sample size is a likely reason for the variability of findings in studies of IL1 gene cluster members and AS reported to date, and will also lead to confusion in replication studies of other genetic findings, such as those that will appear from genome-wide association studies in the disease. Prospective meta-analysis is one way of maximising sample size and thus providing more robust, believable findings. Unlike retrospective meta-analysis, this form of study is not subject to publication or inclusion bias. A disadvantage is that in the presence of heterogeneity of association between centres, false negative findings may occur. However, although replication studies should ideally analyse large, ethnically homogenous cohorts, prospective international meta-analyses such as this study can be valuable when such cohorts are not available.
IL1A is a proinflammatory cytokine that is mainly produced by activated macrophages. Its actions include augmentation of activation of T and B lymphocytes and monocyte/macrophages; induction of fibroblast proliferation leading to synovial pannus formation; activation of chondrocytes promoting calcium crystal deposition; production of matrix metalloproteinase, prostaglandin E2 and nitric oxide,18 all factors involved in cartilage break-down; and activation of osteoclasts, promoting bone resorption. In contrast with IL1B, IL1A is rarely found in the extracellular compartment, and is primarily present either as a membrane bound or intracellular protein. We are not aware of any data regarding tissue expression of IL1A in human AS. In mice, IL1A over-expression leads to a severe chronic proliferative inflammatory arthritis,19 but the role of IL1 has not been studied in animal models of AS. The locations of the IL1A SNPs genotyped are: rs2856836 the 3’UTR, rs17561 in exon 5 producing the substitution Ser114Ala and rs1894399 in intron 3. There is little data regarding the effect of the SNPs rs2856836 and rs1894399 on expression or function of IL1A. SNP rs17561 has been shown to be more resistant to calpain cleavage, an essential activation step, thought to allow IL1A to avoid nuclear translocation, and instead be released by cells to exert autocrine and paracrine effects.20 Two studies of IL1 inhibition with anakinra have been reported in AS. Haibel et al reported an open label, observational study of anakinra treatment in 19 patients with severe NSAID resistant disease.21 Of these patients, 11 had ASAS 20 improvements or greater. Tan et al studied nine severe patients with AS, and reported significant improvements in all clinical, laboratory and MRI measures of disease severity, with six cases achieving ASAS 20% responses.22 These studies support a role for IL1 in AS, although the response to anakinra blockade was modest by comparison with that observed with TNF antagonists. This may relate to pharmacodynamic or pharmacokinetic properties of anakinra, such as its short half-life. An alternative hypothesis is that IL1A may induce inflammation in AS through intracellular effects that may not be inhibited by anakinra.23 It is also possible that IL1A is involved in the initiation of disease, but has a less significant role in established disease.
In conclusion, this study indicates that IL1A variants are significantly associated with AS, and that IL1A joins the short list of genes with strong, independently replicated, association with the disease. It provides a model for a form of study that has the required power to replicate the magnitude of genetic effects likely to operate in AS. Further research to explain the mechanism of association of IL1A is clearly warranted.
Acknowledgments
We are grateful to the many patients who contributed samples to these studies and to their doctors for allowing us to study their patients.
REFERENCES
Footnotes
Funding: This study was funded by the Arthritis Research Campaign (UK) and by National Institute of Arthritis and Musculoskeletal and Skin Diseases grants 1PO1-052915-01, RO1 AR046208 and RO1-AR048465. MAB was funded by the National Health and Medical Research Council (Australia).
Competing interests: None declared.
Ethics approval: All cohorts had been recruited with the approval of their local ethics committees.