Article Text

Abstract
Objectives To define in patients affected by familial Mediterranean fever (FMF) whether or not interleukin (IL)-1β secretion (1) is enhanced, (2) correlates with the type of MEFV mutation and (3) is mediated by NLRP3.
Methods Freshly isolated monocytes from 21 patients with FMF (12 homozygous and 9 heterozygous), 14 MEFV healthy carriers and 30 healthy donors (HDs), unstimulated or after lipopolysaccharide (LPS)-induced activation, were analysed for redox state (production of reactive oxygen species (ROS) and antioxidant responses) and IL-1β and IL-1 receptor antagonist (IL-1Ra) secretion. NLRP3 down-modulation was induced by in vitro silencing of the NLRP3 gene.
Results LPS-stimulated monocytes from patients with FMF displayed enhanced IL-1β secretion, which correlated with number and penetrance of MEFV mutations. Silencing of NLRP3 consistently inhibited IL-1β secretion. As in other autoinflammatory diseases, FMF monocytes produced more ROS than genetically negative cells from HDs. Unlike in cryopyrin-associated periodic fever syndromes (CAPS), however, they were characterised by a conserved and sustained antioxidant response. Consistent with this finding, activated MEFV-mutated monocytes did not exhibit the functional indicators of oxidative stress observed in CAPS, including accelerated IL-1β secretion and deficient production of IL-1Ra.
Conclusions MEFV-mutated monocytes display enhanced IL-1β secretion, which correlates with number of high-penetrance mutations and level of endogenous ROS. Unlike NLRP3-mutated cells, monocytes carrying MEFV mutations withstand oxidative stress and preserve IL-1Ra production, thereby limiting inflammation. Finally, in contrast with that found in the animal model, the increased secretion of IL-1β by LPS-stimulated FMF monocytes is NLRP3-dependent.
Statistics from Altmetric.com
Introduction
Familial Mediterranean fever (FMF)1 ,2 is the most common of the hereditary autoinflammatory disorders, and displays a clinical course characterised by bouts of painful inflammation.2 The effectiveness of interleukin (IL)-1β blockers in colchicine-resistant patients3 has suggested that IL-1β plays a role in the pathophysiology of the disease. However, direct evidence of dysregulated IL-1β secretion in patients with FMF is so far missing.
IL-1β is a major proinflammatory mediator mainly produced by monocytic cells, triggering inflammatory responses in most tissues.4 IL-1β requires processing by the inflammasome to become biologically active.5 ,6 Inflammasomes are cytoplasmic multiprotein complexes consisting of NOD-like receptor family members, such as the NLRP proteins, adaptor proteins and the effector subunit caspase-1.5 ,6 One of the best characterised inflammasomes comprises the NOD-like receptor, NLRP3, and it is involved in IL-1β maturation in response to microbial and tissue factors.5 ,6 Aberrant secretion of IL-1β is the hallmark of a number of autoinflammatory diseases, including cryopyrin-associated periodic fever syndromes (CAPS), that are due to mutations in NLRP3 (or cryopyrin).7 The IL-1 dependence of CAPS has been confirmed by the dramatic responsiveness to anti-IL-1 treatment.8 ,9
FMF is caused by mutations of the pyrin-encoding MEFV gene,10 ,11 which is predominantly expressed in innate immune cells.12 The function of both wild-type and mutated pyrin is highly debated. Both pro- and anti-inflammatory roles have been suggested for pyrin. Namely, the wild-type pyrin has been proposed to bind pro-caspase-1 and other components of inflammasomes, including NLRP3, preventing caspase-1 activation.13–15 The subsequent suppressive effect on IL-1β secretion would be lost in the mutated protein. In contrast, increased IL-1β release by Mefv−/− macrophages in response to a spectrum of inflammatory stimuli has been recently shown in a mouse line lacking the Mefv gene.16 Other studies showed that, under particular conditions, pyrin mediates the activation of caspase-1 independently of NLRP3, by forming the ‘pyrin inflammasome’, composed of pyrin, apoptosis-associated speck-like protein containing a caspase recruitment domain (ASC) and caspase-1.17–19 In agreement with this mechanism, homozygous knock-in mice harbouring the mouse pyrin protein fused to the human B30.2 domain, which contains mutations associated with FMF, were recently shown to develop an autoinflammatory phenotype through the formation of the pyrin inflammasome in response to Toll-like receptor agonists.20 This model suggests that pyrin variants are gain-of-function mutations causing an IL-1β-mediated autoinflammatory disease independently of NLRP3.
Increasing observations have drawn attention to the role of redox signalling in NLRP3 inflammasome activation.21–25 Accordingly, conditions of redox distress in inflammatory cells influence IL-1β production.26 ,27 We have recently reported that, compared with healthy cells, NLRP3-mutated monocytes from patients with CAPS display higher levels of both reactive oxygen species (ROS) and antioxidants.28 The precarious redox equilibrium characterised in CAPS monocytes is further deranged by inflammatory stimuli, causing collapse of the antioxidant systems.29 ,30 Functional effects of this redox distress are accelerated IL-1β secretion28 and decreased production of cytokines downstream of IL-1, such as IL-1 receptor antagonist (IL-1Ra) and IL-6.29 ,30 The impaired released of these cytokines coupled with increased IL-1β secretion explains the predominant role of IL-1β in CAPS. Interestingly, monocytes from patients affected by other genetic autoinflammatory diseases, such as NLRP12-associated autoinflammatory disorders31 and tumour necrosis factor receptor-associated periodic syndrome (TRAPS),32 also exhibit higher levels of ROS.
We investigated in monocytes from patients with FMF whether secretion of IL-1β and downstream cytokines (1) is affected, (2) correlates with the type of MEFV mutation, and (3) is mediated by NLRP3.
Patients and methods
Subjects
Twenty-one patients with FMF were evaluated and compared with 14 MEFV-mutated healthy carriers (HCs), 30 healthy donors (HDs) and one patient with CAPS (i.e. carrying an NLRP3 mutation)29 as a control. Disease activity was evaluated at the time of sampling by assessing clinical manifestations (fever and/or other FMF-related symptoms) and laboratory tests, including erythrocyte sedimentation rate (ERS), C-reactive protein (CRP) and serum amyloid A (SAA) (table 1). All the patients were analysed in a symptom-free period. At the time of sample collection, 16 patients were under colchicine treatment with complete control of clinical manifestations. In five patients, concomitant elevation of acute phase reactants was observed. The study was approved by the ethics board of G Gaslini Institute.
Genotype and clinical characterisation of FMF patient cohort
Monocyte isolation
Monocytes were isolated by adherence23 ,26 ,28 ,29 from peripheral blood mononuclear cells (PBMCs) obtained from patients, their parents and/or siblings (i.e. HCs) and HDs.
In vitro activation of monocytes
Monocytes enriched aforementioned were cultured in RPMI 1640 supplemented with 10% fetal calf serum in the presence or absence of lipopolysaccharide (LPS; 1 μg/ml).23 ,26 ,28 ,29 Supernatants and cell pellets were collected at the end of each experimental time point.
NLRP3 mRNA silencing
PBMCs were isolated from 23 subjects including 7 patients with genetically confirmed FMF (i.e. carrying two mutations in trans), 9 MEFV HCs and seven HDs. Three million cells were nucleofected with 5 μg of mock or NLRP3 small interfering (si)RNA (Invitrogen) using Nucleofector Technology (Amaxa) and following manufacturers’ instructions. Transfected cells were seeded in 96-well plates (7.5×105/well) and incubated for 24 h in nucleofection medium enriched with 10% fetal calf serum, 10% l-glutamine and interferon γ (20 ng/ml).26 Cells were then stimulated with 1 μg/ml LPS for an additional 18 h. In these experimental conditions, levels of secreted IL-1β are lower than those detected in supernatants of monocytes stimulated immediately after isolation.26
Cytokine detection
Levels of IL-1β and IL-1Ra in monocyte supernatants were quantified by ELISA (R&D Systems).29
Intracellular ROS detection
Monocyte ROS levels were measured as described.23 ,26 ,28 ,29 Namely, adherent monocytes were incubated with 10 μM fluorescent dye (2,7-dichlorodihydrofluorescein diacetate (H2DCF-DA) Molecular Probes) for 30 min. Fluorescence was then measured in cell lysates, and data were normalised to protein content.23 ,26 ,28 ,29
Real-time PCR
Total mRNA was isolated from cells using TriPure Isolation Reagent (Roche Applied Science) and reverse-transcribed with the QuantiTect Reverse Transcription Kit (Qiagen). Real-time PCR was performed using Platinum SYBR green qPCR SuperMix-UDG (Invitrogen). The specific primers for NLRP3, thioredoxin, cystine transporter (xCT) and glyceraldehyde-3-phosphate dehydrogenase (GAPDH) have been described previously.23 ,26 ,28 Target gene level was normalised to that of GAPDH mRNA, and relative expression was determined using the ΔCt method. 23 ,26 ,28
Measurement of extracellular reduced cysteine
Extracellular cysteine was detected by the DNTB (5,5′-dithiobis-(2-nitrobenzoic acid)) assay as described.23 ,26 ,28 ,29 Namely, 0.1 ml supernatant or cysteine standard was allowed to react with 10 nM DTNB, and absorbance at 412 nm was measured.
Determination of protein synthesis by endogenous labelling and trichloroacetic acid precipitation
Monocytes were loaded with [35S]methionine/cysteine (100 mCi/ml, Easy Tag Express Protein Labelling Mix, [35S]; Perkin–Elmer) in methionine/cysteine-free RPMI 1640 medium at various times after LPS exposure. After 1 h, cells were lysed and aliquots of cell lysates were precipitated in 25% unlabelled trichloroacetic acid. Insoluble radioactivity was measured in a β-counter after addition of scintillation liquid.29
Statistical analysis
Data were displayed in dot-plots, whisker box plots or as median values, and differences among groups were evaluated using the non-parametric Kruskal–Wallis test followed by appropriate post hoc analyses.
Results
Monocytes from patients carrying pyrin mutations secreted higher levels of IL-1β
Secretion of IL-1β was assessed in primary monocytes freshly isolated from patients with FMF (12 homozygous; eight heterozygous or with two mutations in cis) or genetically negative HDs (n=30). Resting monocytes from both patients and healthy subjects did not secrete IL-1β when cultured in vitro without stimuli (data not shown). However, after LPS stimulation, higher levels of IL-1β were found in supernatants from FMF monocytes compared with healthy monocytes (figure 1A). Heterogeneity analysis using the Kruskal–Wallis test revealed that significant differences existed among the groups (p=0.0003). Post hoc tests then showed that both genetically confirmed (p=0.0001) and heterozygous patients with FMF (p=0.005) displayed significantly enhanced IL-1β secretion (figure 1A). Interestingly, the four patients with FMF displaying increased acute phase reactants at the time of monocyte isolation (figure 1A, white symbols) were distributed in the group secreting higher levels of IL-1β.

Pyrin-mutated human monocytes display enhanced lipopolysaccharide (LPS)-induced interleukin (IL)-1β secretion. Primary monocytes from patients with familial Mediterranean fever (FMF) (n=20; 12 genetically confirmed homozygous (+/+) and 8 heterozygous (+/−) for MEFV mutations) or genetically negative healthy donors (HDs) (n=30) were stimulated with LPS for 18 h. Supernatants were then collected, and IL-1β production was assessed by ELISA. Results are expressed as ng/ml. (A) Dot-plot displaying levels of IL-1β secreted by each HD subject (circles) and patients with FMF (triangles); group median values (horizontal black lines) are shown. White triangles correspond to patients with increased levels of acute phase reactants at the time of sampling. Heterogeneity analysis was performed with the Kruskal–Wallis test. Post hoc analyses revealed significant increases in IL-1β (***p=0.0001; **p=0.005) (B) Pedigree of three selected FMF families and the corresponding analysis of LPS-induced IL-1β secretion are depicted. Results are expressed in ng/ml as in (A), and compared with HD median value. (C) LPS-induced IL-1β secretion by HD, FMF and healthy carrier (HC) cohorts, divided into subsets according to the presence of number and penetrance of the mutations carried. Grey-scaled coloured whisker box plot displaying median values of IL-1β production in HDs, patients with FMF and MEFV HCs are shown. Heterogeneity analysis was performed with the Kruskal–Wallis test. Post hoc analyses revealed significant increases in IL-1β in the presence of two high-penetrance mutations (**p=0.005) and one high-penetrance mutations (*p=0.05) in patients with FMF and in HCs, respectively.
IL-1β secretion correlated with number and penetrance of MEFV mutations
We have previously reported that the frequency of FMF-like symptoms decreases from patients carrying two high-penetrance mutations to patients with a single low-penetrance mutation.33 Analyses performed in single families in the present study support this concept. Namely, data derived from representative families and depicted in figure 1B revealed that an increased number or penetrance of MEFV mutations correlated with higher levels of secreted IL-1β. Moreover, increased secretion was observed not only in patients but also in the MEFV-carrying HCs (figure 1B,C). In particular, in HCs carrying one mutated allele (e.g. M694V or K695R in heterozygosis), IL-1β release was higher than in HDs but lower than in the patients homozygous for either M694V or K695R (figure 1B). In addition, in the family displayed at the top right of figure 1B, the parent carrying the high-penetrance mutation (i.e. M680I) secreted levels of cytokine that were intermediate between the parent carrying the low-penetrance mutation (i.e. R761H) and the patient carrying both (M680I/R761H). To explore further whether or not differences in number and penetrance in MEFV mutations consistently lead to variations in IL-1β release, we further divided the FMF cohort and 12 HCs into subsets on the basis of penetrance and number of MEFV mutations carried. Levels of LPS-induced IL-1β were thus compared among genetically negative subjects (HDs) and patients and/or HCs carrying two high-penetrance mutations, one high-penetrance and one low-penetrance mutation, two low-penetrance mutations, one high-penetrance mutation and one low-penetrance mutation.32 As shown in figure 1C, IL-1β secretion correlated with the number and penetrance of MEFV mutations. In spite of the low numbers of subjects in each group, the heterogeneity test was positive (p=0.0020), and post hoc analyses revealed significant increases in IL-1β in the presence of two high-penetrance mutations and one high-penetrance mutation in patients with FMF (p=0.005) and HCs (p=0.05), respectively (figure 1C).
Silencing of NLRP3 in monocytes from patients and controls inhibited LPS-induced IL-1β secretion
As mentioned above, the actual involvement of the NLRP3 inflammasome in the pathophysiology of pyrin-related disease is controversial.13–17 ,20 Thus, we investigated the role of NLRP3 in freshly isolated human monocytes using the silencing technique. First, we studied a selected FMF family comprising two HCs, one with high-penetrance (ie, M680I) and one with low-penetrance (i.e. R761H) pyrin mutation, and one patient carrying both mutations. As shown in figure 2A, LPS-induced IL-1β release by monocytes from both the patient and the two parents was strongly inhibited by transfection with NLRP3 siRNA (white bars), but not with scrambled siRNA (grey bars).

Lipopolysaccharide (LPS)-induced interleukin (IL)-1β secretion in familial Mediterranean fever (FMF) monocytes is NLRP3-dependent. Peripheral blood mononuclear cells isolated from 23 subjects including patients with genetically confirmed FMF (n=7), MEFV healthy carriers (HCs) (n=9) and healthy donors (HDs) (n=7) were nucleofected with 5 μg NLRP3 small interfering (si)RNA (white bars) or appropriate mock control (grey bars), incubated for 24 h in fresh enriched nucleofection medium and then activated with 1 μg/ml LPS for an additional 18 h. Supernatants were then collected for IL-1β detection by ELISA. Results are expressed as pg/ml. (A) Pedigree of a representative FMF family. Seven wild-type pyrin HDs, one M680I/R761H(+) FMF patient, M680I(+) and R761H(+) HC parents, after nucleofection were treated with LPS (1 μg/ml). Comparison of IL-1β secretion is displayed as median values in HDs and absolute level in patients and HCs. (B) Comparison of IL-1β production by monocytes from HDs and a larger cohort of patients with FMF and HCs treated with LPS (1 μg/ml) after nucleofection. Heterogeneity analysis was performed using the Kruskal–Wallis test (p=0.0001). Downregulation of NLRP3 expression by NLRP3 siRNA (white bars) significantly inhibited LPS-induced secretion of IL-1β in both HD and FMF (**p=0.005, *p=0.05) subjects. (C) Total mRNA was isolated from monocytes nucleofected with NLRP3 siRNA from selected HDs, patients with FMF and HCs as indicated. Gene expression of NLRP3 (black bars) and MEFV (white bars) was determined by real-time PCR. Target gene levels were normalised to that of glyceraldehyde-3-phosphate dehydrogenase, and results expressed relative to levels in monocytes nucleofected with control siRNA according to ΔCt methods.
We then extended the analysis to a group of seven HDs, seven patients with FMF and nine HCs (figure 2B). In all cases, siRNA nucleofection downregulated NLRP3 mRNA by 50–80% in both wild-type and pyrin-mutated monocytes, whereas scrambled RNA did not affect NLRP3 mRNA expression (not shown). NLRP3 down-modulation resulted in a dramatic inhibition of LPS-induced secretion of IL-1β in both HD (p=0.05) and FMF (p=0.0001) subjects (figure 2B, white bars), indicating that the IL-1β hypersecretion observed in LPS-stimulated monocytes from patients with FMF was actually dependent on NLRP3 activity. Real-time PCR analyses of the MEFV gene in NLRP3-silenced monocytes from patients, HCs and HDs revealed that the expression of MEFV was not modulated by NLRP3 siRNA (figure 2C), ruling out the possibility that the inhibition of IL-1β secretion by NLRP3-silenced monocytes was due to concomitant dysregulation of the MEFV gene.
FMF monocytes produced more ROS but displayed an effective antioxidant response
We have previously shown that monocytes obtained from patients with CAPS, and thus expressing mutated NLRP3, displayed elevated levels of ROS, which were further induced by LPS.28 ,29 The LPS-induced antioxidant response aimed at detoxifying ROS is more rapid in CAPS monocytes than in healthy monocytes but eventually collapses, causing oxidative stress. Two main functional consequences of these redox alterations in CAPS are: (1) prompt activation of the NLRP3 inflammasome with earlier appearance of secreted IL-1β28; and (2) deficient production of cytokines downstream of IL-1β such as IL-1Ra.28 Thus, we investigated whether or not FMF monocytes were characterised by redox alterations and functional consequences similar to those occurring in NLRP3-mutated cells. Analyses of ROS production in freshly isolated, unstimulated cells revealed that, like monocytes carrying NLRP3 gene variants, MEFV-mutated monocytes had significantly higher ROS levels at baseline (figure 3A, black bars), further induced by LPS (white bars). However, unlike CAPS monocytes, the antioxidant response to LPS was sustained. Namely, the antioxidant genes (e.g. thioredoxin and the cystine transporter xCT) induced by LPS followed a physiological pattern of induction in LPS-stimulated FMF monocytes, with kinetics similar to HDs and different from CAPS (figure 3B,C). xCT and thioredoxin are components of the cystine/cysteine redox cycle, which mediates uptake of oxidised cystine and extracellular release of reduced cysteine.30 Consistent with the sustained increase in xCT and thioredoxin mRNA expression, the release of cysteine, a reliable reflection of the antioxidant activity of activated monocytes,23 ,26 ,28 ,29 was high 18 h after LPS exposure, as in HDs and contrary to what occurs in NLRP3-mutated monocytes (figure 3D).28 ,29

Pyrin-mutated human monocytes are characterised by a physiological pattern of interleukin (IL)-1β secretion in the presence of an overall conserved antioxidant machinery. (A) Monocytes from healthy donors (HDs) (n=8), homozygous (+/+) (n=6) and heterozygous (+/−) (n=4) patients with familial Mediterranean fever (FMF), and healthy carriers (HCs) (n=6) were activated in vitro for 1 h with lipopolysaccharide (LPS), and intracellular reactive oxygen species (ROS) production was detected using 2,7-dichlorodihydrofluorescein diacetate. Data are expressed as relative fluorescence units (RFU) and displayed as median values. Heterogeneity analysis was performed among untreated (-, black bars) groups using the Kruskal–Wallis test (p=0.0056). Post hoc test revealed a significant difference between ROS produced by unstimulated HDs versus unstimulated +/+ FMF and HCs (black bars, *p<0.05). (B, C) Monocyte total mRNA was isolated from four HDs, two representative +/+ patients with FMF (Nos 4 and 21) and one patient with cryopyrin-associated periodic fever syndrome (CAPS), as indicated, at time 0 or after 3 and 6 h of activation with LPS. Gene expression of thioredoxin (B) and xCT (C) was determined by real-time PCR. Target gene levels were normalised to that of glyceraldehyde-3-phosphate dehydrogenase, and results expressed relative to levels at time 0 according to ΔCt methods. (D) Cysteine levels in supernatants of monocytes from HDs and patients with FMF or CAPS stimulated for 18 h with LPS were measured by 5,5′-dithiobis-(2-nitrobenzoic acid) assay. Data are expressed as μM/18 h and displayed as median values±error bars. (E) Primary monocytes from patients with FMF (n=17) or HDs (n=28) were exposed to LPS, and supernatants were analysed for IL-1β content by ELISA after 3, 6 and 18 h of culture. Data are displayed as median values.
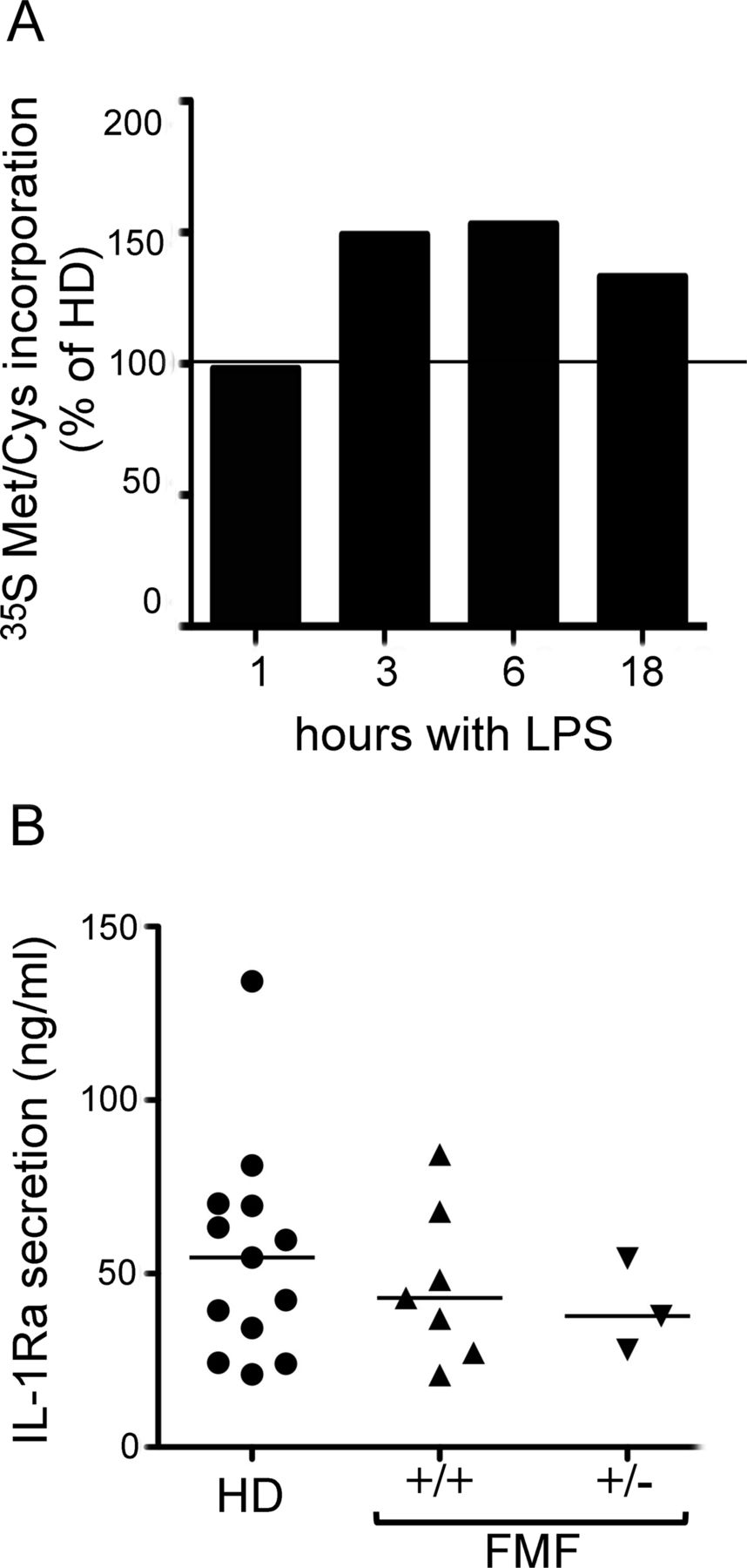
These data suggest that, in spite of the higher ROS level, FMF monocytes have a conserved and efficient antioxidant activity capable of restraining the oxidative stress due to MEFV mutations. Actually, unlike in CAPS monocytes, IL-1β secretion by FMF monocytes was unexpectedly enhanced compared with HDs (figure 3E). Likewise, patients with FMF did not show any stress-related defects in protein synthesis (figure 4A) nor the down-modulation of IL-1Ra production (figure 4B) that occurs in CAPS.29
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Pyrin mutations do not result in any stress-related defects in protein synthesis and interleukin 1 receptor antagonist (lL-1Ra) production. (A) Acid-insoluble [35S]methionine/cysteine incorporation by monocytes from six healthy donors (HDs) and one representative patient with +/+ familial Mediterranean fever (FMF) (No 18), exposed to lipopolysaccharide (LPS) for 1, 3, 6 or 18 h and incubated for 1 h with [35S]Met/Cys at each time point. Data are expressed as percentage of [35S]Met/Cys incorporation by healthy monocytes (n=6) labelled in parallel (taken as 100%). (B) Monocytes isolated from 7 homozygous (+/+) and 3 heterozygous (+/−) patients with FMF and 13 HDs were stimulated for 18 h with LPS, and IL-1Ra content in supernatants was assessed by ELISA. Dot-plot displaying levels of IL-1Ra secreted by each HD (circles) or patients with FMF (triangles), and group median values (horizontal black lines). Heterogeneity analysis was assessed using the Kruskal–Wallis test (p=0.7141).
Discussion
In this study, we compared the redox state at baseline and after LPS stimulation in monocytes from HDs, patients with FMF and HCs, and correlated it with the rate of secretion of IL-1β and downstream cytokines, such as IL-1Ra.
The first important finding of the study is that the presence of mutated pyrin in monocytes is associated with higher levels of LPS-induced secretion of IL-1β. Moreover, the presence of a ‘dosage effect’ of the mutated pyrin was disclosed by the observation that increased IL-1β secretion in HCs and patients with FMF correlates with the number and penetrance of pyrin mutations. Notably, these data support our previous observations that the frequency of FMF-like symptoms decreases from patients carrying two high-penetrance mutations to patients with a single low-penetrance mutation.33
The present study also shows that, compared with healthy cells, MEFV-mutated monocytes display higher levels of ROS at baseline, placing FMF in the emerging group of autoinflammatory diseases characterised by increased ROS production, and including CAPS,27 ,28 NLRP12-associated autoinflammatory disorders,31 and TRAPS.32 Thus, growing evidence indicates that loss of redox homoeostasis is a hallmark of monocytes in genetic autoinflammatory syndromes.30 Redox alterations are also found in other genetic diseases.34–38 In some cases, misfolded proteins or protein aggregates cause redox stress in the presence of mutations, whereas, in others, the functional link between mutation and cell stress is still unclear.34–38 Given the absence of a clear definition of the subcellular localisation and function of pyrin, the mechanistic link between mutant pyrin and increased ROS is missing in FMF. Nonetheless, the evidence of higher ROS levels in patients carrying two mutated alleles compared with heterozygous patients suggests the presence of a direct correlation between number of mutated pyrin molecules and degree of stress in monocytes. In turn, the amount of ROS correlates with the levels of LPS-induced IL-1β secretion, in both patients and HCs. This finding supports the concept that redox remodelling, with ROS production as an initial event,5 ,23 ,24 ,27 is required for IL-1β production and inflammasome activation. As a consequence, both processes seem to increase in stressed cells.30 Moreover, the strong correspondence between high-penetrance mutations and IL-1β secretion suggests that different mutations may cause different levels of cell stress. The higher redox distress may then be responsible for the higher secretion of IL-1β.
Unlike CAPS monocytes, in which the high level of ROS at baseline induces a rapid rise in antioxidant defences followed by their collapse,28 ,29 in FMF monocytes the antioxidant response is sustained and protects monocytes from oxidative stress. The main functional consequence of preserving the redox balance is that, unlike in CAPS,29 the production of IL-1Ra is fully maintained in FMF. The normal levels of IL-1Ra, which plays a crucial role in limiting inflammation,39 may explain the typical rapid self-limitation of fever attacks in patients with FMF, which is not observed in patients with CAPS.
Although the significance of IL-1β in the pathogenesis of FMF is supported by the spectrum of clinical manifestations1 ,2 and by the therapeutic responsiveness to IL-1 blockers,3 how mutant pyrin actually modulates IL-1β production is still highly debatable. Both loss-of-function and gain-of-function models have been proposed. While the former suggests that wild-type pyrin exerts a suppressive effect on inflammasome activation which is lost in the mutated protein,13 ,15 the latter proposes that mutant pyrin causes caspase-1 activation by interacting with ASC without the involvement of NLRP3.20 These discrepancies may be due, at least in part, to the different experimental systems used. Most data indicating a loss-of-function of mutant pyrin are generated in vitro using transfected cells,13 ,15 whereas the model suggesting that mutated pyrin increases caspase-1 activation is derived from a knock-in murine model, expressing the mouse pyrin protein fused to a human domain.20 Our approach, based on the analyses of monocytes freshly isolated from patients naively carrying genetic variants of the MEFV gene, provides two major advantages, by avoiding: (1) in vitro manipulation of cell lines that display a redox state profoundly different from primary cells and may not represent the relevant counterpart of primary inflammatory cells; and (2) mixing human and mouse proteins, which might lead to unforeseen artefacts. By using this approach, we found that the downregulation of NLRP3 in human MEFV-mutated monocytes inhibits IL-1β secretion, supporting the hypothesis that a direct functional relationship between pyrin and the NLRP3 inflammasome occurs,15 at least after LPS stimulation. Our findings are consistent with a model of gain-of-function mutation, where the penetrance (higher or lower) and the mutant protein dosage affect IL-1β secretion to different extents, and consequently influence the severity of the disease. However, our observations do not preclude the existence of a pyrin inflammasome triggered by different stimuli.17–19 Moreover, the possibility that mutations result in loss of a regulatory function, proportional to penetrance and dosage and thus more evident in patients carrying two mutations at high penetrance, cannot be excluded.
In conclusion, we have shown that MEFV-mutated monocytes are characterised by a mild state of stress and IL-1β oversecretion, which increase proportionally with the number and penetrance of mutations, thus supporting the dosage effect observed in animal models and clinical studies.20 ,33 In human monocytes, the presence of mutated pyrin is not sufficient to promote the LPS-mediated secretion of IL-1β independently of the activation of the NLRP3 inflammasome. However, the actual functional relationship between pyrin and NLRP3 remains unclear and deserves further investigation.
Acknowledgments
We thank Ms Caterina Pellecchia for help with ELISA, and the NCI (Biological Resources Branch) for the anti-IL-1β 3ZD monoclonal antibody. We also thank patients and families for their outstanding collaboration and Drs Roberta Caorsi, Silvia Federici and Matteo Doglio for their invaluable assistance with the selection and management of patients.
References
Footnotes
Handling editor Tore K Kvien
MG and AR contributed equally.
-
Contributors Study conception and design: AO, SC, LD, AM, MG, AR. Acquisition of data: AO, SC, LD, MG, AR. Analysis and interpretation of data: AO, SC, LD, MG, AR. All authors were involved in drafting the article and discussing the data. AR and MG had full access to all the data in the study. MG and AR take responsibility for the integrity of the data and the accuracy of the data analysis.
-
Funding Compagnia San Paolo and Ricerca Corrente from the Italian Ministry of Health. SC is the recipient of the ‘Young Investigators’ grant GR-2010-2309622 from the Italian Ministry of Health.
-
Competing interests None.
-
Patient consent Obtained.
-
Ethics approval Ethics board of G Gaslini Institute, Genova, Italy.
-
Provenance and peer review Not commissioned; externally peer reviewed.