Article Text

Abstract
Objective Adipose tissue is known to release inflammatory cytokines and growth factors. In this exploratory study, the authors examined whether the infrapatellar fat pad (IPFP) closely located to cartilage in the knee joint can affect cartilage metabolism. In addition, the authors analysed whether the macrophage types present in IPFP could explain the effect on cartilage.
Methods IPFP explants obtained during total knee replacement of 29 patients with osteoarthritis (OA) were used to make fat-conditioned medium (FCM). Explants of bovine cartilage were cultured with or without FCM. Nitric oxide (NO) and glycosaminoglycan release and gene expression of matrix-degrading enzymes in cartilage were analysed. To stimulate catabolic processes in the cartilage, the authors added interleukin 1β, and the effect of six FCMs was evaluated. The presence of different types of macrophages (CD68+, CD86+ and CD206+) in OA IPFPs was compared with subcutaneous adipose tissue samples and IPFP samples from patients with an anterior cruciate ligament rupture.
Results FCM alone reduced NO and glycosaminoglycan release and matrix metalloproteinase (MMP)1 gene expression by the cartilage. Moreover, when catabolic conditions were enhanced with interleukin 1β, FCM inhibited NO production as well as MMP1 and MMP3 gene expression and increased collagen type II gene expression. Significantly more CD206+ cells were present in OA IPFP samples than in subcutaneous fat or anterior cruciate ligament IPFP samples.
Conclusion In contrast to the authors' expectations, medium conditioned by end-stage OA IPFP inhibited catabolic processes in cartilage. CD206+ cells present in the IPFPs used for making the FCM might have contributed to the inhibition of catabolic processes in the cartilage.
Statistics from Altmetric.com
Introduction
Osteoarthritis (OA) is characterised by cartilage damage, bone alterations and inflammation of the synovium often seen in OA. OA is a multifactorial disease with a greater incidence in women, obese individuals and older subjects.1 2
Isolation and energy storage are two well-known and important functions of adipose tissue. However, adipose tissue is also known to produce inflammatory mediators and adipokines such as interleukin (IL)-1, IL-6, IL-8, monocyte chemotactic protein 1, leptin, adiponectin and others.3 4 The infrapatellar fat pad (IPFP) is an integral part of the joint together with cartilage, ligaments, menisci and synovium. The main role of IPFP is to facilitate the distribution of synovial fluid and distribute mechanical forces through the knee joint. The IPFP is situated in the knee underneath the patella, between the patellar tendon, femoral condyle and tibial plateau5 and thus located closely to the synovial layers and cartilage surfaces where it can influence these structures. Several adipokines and cytokines are known to be locally produced in the knee joint by the IPFP,6,–,9 enabling the IPFP to influence inflammatory processes in the knee. In addition, the IPFP was recently found to be a high producer of IL-6, even higher than subcutaneous adipose tissue.10
Adipose tissue consists of adipocytes lying in the stromal vascular fraction and is often infiltrated by immune cells such as macrophages. In addition, T cells, B cells, natural killer cells and mast cells are also found in adipose tissue.11,–,15 Many secreted proteins are derived from the non-adipocyte fraction of adipose tissue,16 and it is even suggested that most of the cytokines produced by the adipose tissue are macrophage derived.17 18
The macrophage phenotype can change in response to cytokines and growth factors,19 20 resulting in different populations of macrophages with distinct functions. Classically activated macrophages (M1) are important immune cells that are vital to host defence; they can propagate inflammatory responses by producing cytokines IL-1β, tumour necrosis factor (TNF)α, IL-6, IL-12 and IL-23.21 Another subtype of macrophages is the alternatively activated macrophages (M2).22 23 These cells express the mannose receptor CD206, scavenger receptors and IL-1 receptor antagonist. M2 macrophages produce predominantly IL-10 and almost no IL-12 and IL-23, indicative for an anti-inflammatory phenotype.21
Taken together, adipose tissue can be considered as an organ that secretes factors that can contribute to the progression of OA.4 19 24 This holds especially true for the IPFP as a consequence of its anatomical location in the knee joint, close to other joint tissues. The exact effect of IPFP on cartilage in OA has not been elucidated. Here, we hypothesise that the IPFP might exhibit a deleterious effect on cartilage by inducing catabolic changes in chondrocytes.
To investigate whether the IPFP should be considered as an active osteoarthritic joint tissue,5 we investigated the influence of medium conditioned by IPFP on cartilage metabolism. Surprisingly, fat-conditioned medium (FCM) inhibited catabolic processes, which led us to the addition of an extra catabolic stimulus in an exploratory manner. We also analysed whether the type of macrophages present in the IPFP could explain the effect on the cartilage.
Materials and methods
Preparation and analysis of FCM
Twenty-nine IPFPs obtained as anonymous waste material from human subjects with OA who underwent total knee arthroplasty were used to produce FCM. The patients did not give informed consent but did have the right to consent as stated by guidelines of the Federation of Biomedical Scientific Societies (www.federa.org), approved by the local ethical committee in Rotterdam, The Netherlands (MEC-2008-181). The mean age of the donors was 67.9 years (range 54–81 years), and the mean body mass index (BMI) of the donors was 29.6 (range 22.8–48.5). The inner parts of the fat pads where no synovium is present were cut into small pieces of approximately 10 mg and cultured in suspension for 24 h in a concentration of 50 mg tissue/ml in Dulbecco's modified Eagle's medium with Glutamax (GibcoBRL, Grand Island, New York, USA) containing insulin, transferrin, selenic acid and albumin (ITS+, BD Biosciences, Breda, The Netherlands) 100 times diluted, 50 μg/ml gentamicin and 1.5 μg/ml fungizone (both from GibcoBRL). Previously, the release of several signalling molecules from adipose tissue was investigated.16 Based on these results, we chose to incubate the IPFP explants for 24 h. After 24 h, the medium was harvested, centrifuged at 300 g for 8 min and frozen at −80°C in aliquots of 1.5 ml, resulting in 29 different batches of FCM.
Preparation and culture of cartilage explants
The experiment for testing the effect of FCM on cartilage was performed twice: the first time with 17 different FCM batches and the second time with 12 different FCM batches. To be able to test multiple FCM batches in one experiment avoiding cartilage donor differences, we harvested articular cartilage explants from at least eight bovine metacarpophalangeal joints (aged 6–12 months). Full-thickness explants (6 mm diameter, 0.9–1.2 mm thick) were made and pooled followed by random distribution over the wells; three explants were cultured per well and formed one sample. For each of the 29 FCM batches, we had three samples. Explants were precultured for 1 day in Dulbecco's modified Eagle's medium with Glutamax, ITS+ 1:100 followed by FCM 1:1 mixed with fresh medium. Control conditions (one sample of three explants per well, three wells per experiment) were cultured in unconditioned but previously frozen medium 1:1 mixed with fresh medium. To further evaluate the (anti)catabolic effects of FCM, we added 1 ng/ml IL-1β (R&D, Oxon, UK) to cartilage explants during preculture and during the incubation with six different batches of FCM. The six batches for the second set of experiments were based on availability from the 29 earlier tested. Power calculation using the results from the first set of experiments revealed six FCM batches to be sufficient to identify a difference. After 2 days, cartilage explants were snap-frozen in liquid nitrogen and stored at −80°C and medium was collected and stored at −20°C until further use.
RNA isolation and quantitative RT-PCR
Frozen cartilage was processed using a Mikro-Dismembrator S (B. Braun Biotech International GmbH, Melsungen, Germany). RNA was extracted, cDNA was made and gene expression analysis for glyceraldehyde 3-phosphate dehydrogenase, matrix metalloproteinase (MMP)1, MMP3, MMP13, ADAMTS4, ADAMTS5, collagen type II and aggrecan was performed as described previously.25 For primer sequences and RT-PCR conditions, see supplementary methods online.
Biochemical analysis of cartilage culture medium
The amount of glycosaminoglycan (GAG) in the culture medium was quantified using the dimethylmethylene blue assay as described previously.26 The metachromatic reaction of GAG with dimethylmethylene blue was monitored with a spectrophotometer, and the ratio A530:A590 was used to determine the amount of GAG present. Nitric oxide (NO) production was determined by quantifying its derived product, nitrite, in medium using a spectrophotometric method based upon the Griess reaction.27
Characterisation of macrophages in adipose tissue
Subcutaneous adipose tissue was obtained from seven patients being operated for a total knee replacement, from which we also received IPFP, harvested during the total knee replacement proximal of the knee joint. In addition, we obtained IPFPs from five patients undergoing repair of their anterior cruciate ligament (ACL), both with ethical approval (MEC-2008-181). The mean age of subcutaneous adipose tissue donors was 64.8 years (range 54–81 years), and the mean BMI was 31.7 (range 23.5–48.5). The mean age of the ACL IPFP donors was 37.4 years (range 24–58 years), and the mean BMI was 23.5 (range 21.8–27.1). We used antibodies against CD68 (DAKO, Heverlee, Belgium, clone EBM11), CD86 (Abcam, Cambridge, UK, clone EP1158Y) and CD206 (Abcam, clone 15-2) for immunohistochemstry. The staining method can be found in the supplementary methods. Samples were ranked by two blinded observers based on their relative number of positive cells in four sections per sample. Ranking was done for each marker separately, and IPFP, subcutaneous adipose tissue samples and IPFP from patients undergoing ACL repair were given a number between 1 and 41 (29 IPFP samples, 7 subcutaneous samples and 5 IPFPs from ACL repairs) based on the staining intensity. This resulted in two single ranks from each blinded observer (YBJ and CF) for CD68, CD86 and CD206. The average of these two ranks was used for further analysis. To compare OA IPFP and subcutaneous adipose tissue, we used only the samples from patients from which we obtained both adipose tissue types. For the comparison between OA IPFP and ACL IPFP, samples were matched based on BMI.
The stromal vascular fraction of 11 IPFPs was also analysed using flow cytometric analysis for CD45, CD86 and CD206. Details can be found in the supplementary methods.
Statistical analysis
The average gene expression, GAG release and NO production in cartilage cultured in control medium versus FCM and histological scores for OA IPFP and ACL IPFP were compared using a Mann–Whitney U test. For the experiments with(out) IL-1β and FCM, this was done with a Kruskal–Wallis test with a post hoc Dunn's multiple comparison test. The Wilcoxon signed-rank test was used to compare histological scores for OA IPFP and paired subcutaneous adipose tissue (SPSS 18.0).
Results
Effect of FCM on cartilage biology
Cartilage explants were cultured in FCM from 29 different IPFPs to investigate the effect of IPFP on cartilage. Culturing cartilage in FCM resulted in lower NO production (1.9-fold, p=0.004) and GAG release (1.3-fold, p=0.021) than in the untreated controls (figure 1A). FCM downregulated gene expression of MMP1 in comparison to cartilage cultured in the control condition without FCM (6.4-fold, p=0.003). MMP3, MMP13, ADAMTS4 and ADAMTS5 gene expression and collagen type II and aggrecan gene expression were not altered by the FCM (figure 1B,C). To investigate whether the BMI of the IPFP donor influenced the effect of FCM on cartilage, we performed a post hoc subgroup analysis; FCM was made from IPFPs of donors who had a BMI lower than 27 or a BMI equal to or higher than 27. No difference on NO production, GAG release or MMP1 gene expression was observed between the two BMI subgroups. Both groups were still significantly different from the control condition (see supplementary figure 1).

1 Effect of FCM (n=29) on cartilage biology. (A) NO production and GAG release; (B) gene expression of MMP1, MMP13, ADAMTS4 (ATS4) and ADAMTS5 (ATS5); and (C) collagen type II and aggrecan gene expression in healthy cartilage. All expressed relative to the untreated control without FCM (n=6, set at 1). Basal NO content was 63.4±3.5 μM, and basal GAG release was 51.6±5.2 μg/explant. p values indicate a significant difference from the control. FCM, fat-conditioned medium; GAG, glycosaminoglycan; MMP, matrix metalloproteinase; NO, nitric oxide.
FCM counteracts IL-1β-induced effects
In the above-described experiments, the effect of FCM on cartilage was not catabolic in contrast to our hypothesis but rather seemed to inhibit catabolic processes. Although catabolic processes take place as part of normal turnover in healthy cartilage, the expression of most of the matrix-degrading enzymes was very low. Therefore, we conducted experiments where FCM was applied under catabolic conditions (in the presence of IL-1β) to the cartilage to investigate whether IPFP from patients with end-stage OA indeed has anticatabolic effects on cartilage. Cartilage explants were cultured in the presence of 1 ng/ml IL-1β with or without six of the earlier tested FCM batches as proof of principle. When catabolic processes were stimulated, FCM was able to inhibit NO production (p=0.036), MMP1 gene expression (p=0.042) and MMP3 gene expression (p=0.0002) and increase collagen type II gene expression (p=0.012) (figure 2). The same trends were seen for MMP13, ADAMTS4 and ADAMTS5 although not significant. GAG release and aggrecan gene expression were not altered.

FCM lowers IL-1β-induced effects in cartilage (n=6). (A) NO and GAG release, (B) gene expression of extracellular matrix-degrading enzymes and (C) gene expression of matrix proteins. Control without IL-1β or FCM is set at 1 (n=6, dotted line). Basal NO content was 78.2±6.9 μM, and basal GAG release was 48.3±4.6 μg/explant. Box plots indicate relative difference from control. p values indicate a significant difference between the IL-1β-treated condition and IL-1β+FCM. FCM, fat-conditioned medium; GAG, glycosaminoglycan; IL, interleukin; NO, nitric oxide.
Macrophages contributing to the effects seen in cartilage
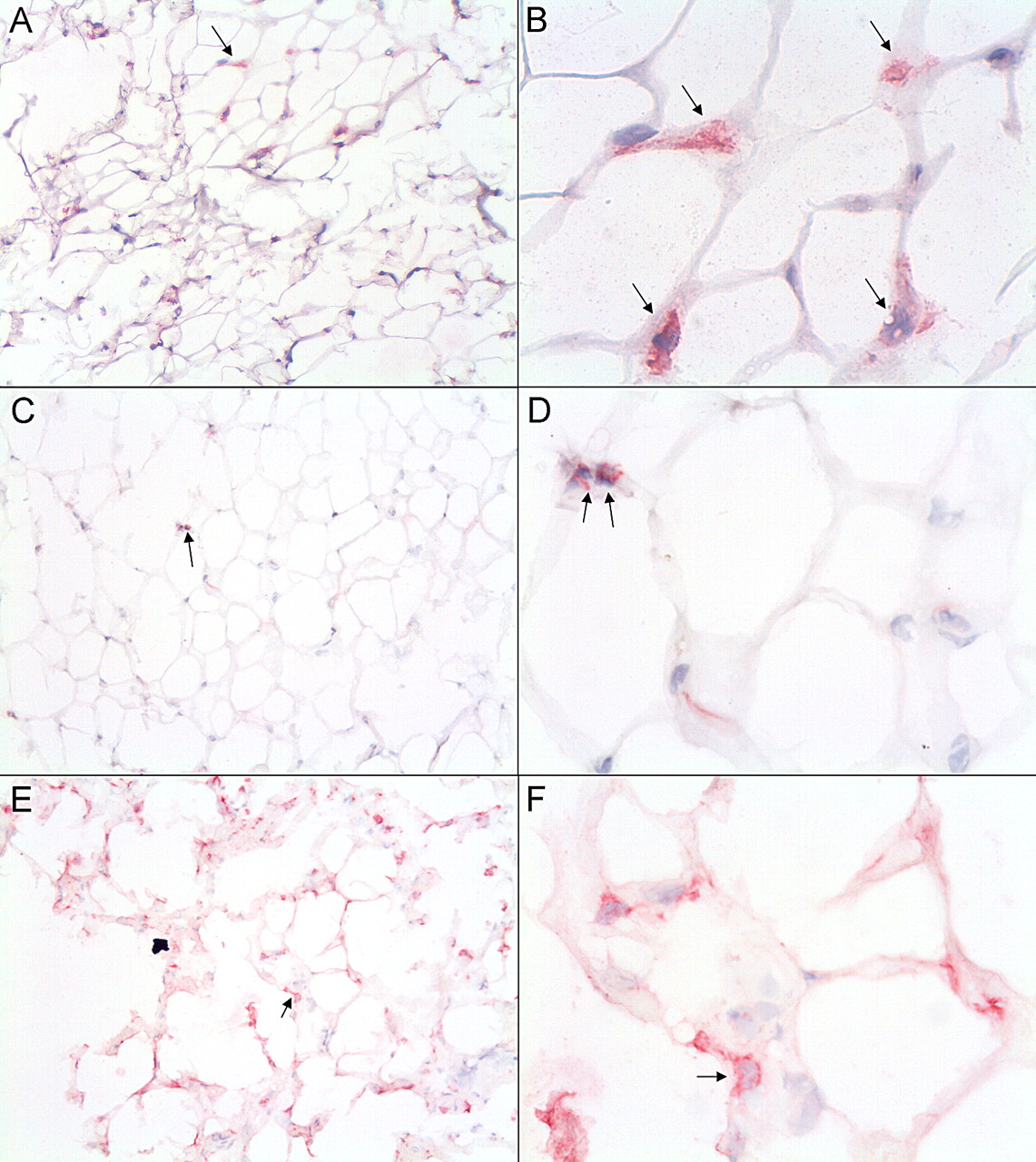
Since the inhibition of catabolic processes in cartilage by FCM was unexpected, we hypothesised that this might be due to the phenotype of macrophages present in the adipose tissue. Macrophages contribute to the secretion of factors by the adipose tissue, and therefore, we determined the presence of macrophages and their phenotype in the IPFPs used to make FCM. Representative immunohistological samples are shown in figure 3. CD68+, CD86+ and CD206+ macrophages were detected in all samples. No correlation was seen between the rank of CD68+, CD86+ or CD206+ cells and the BMI of the IPFP donor.
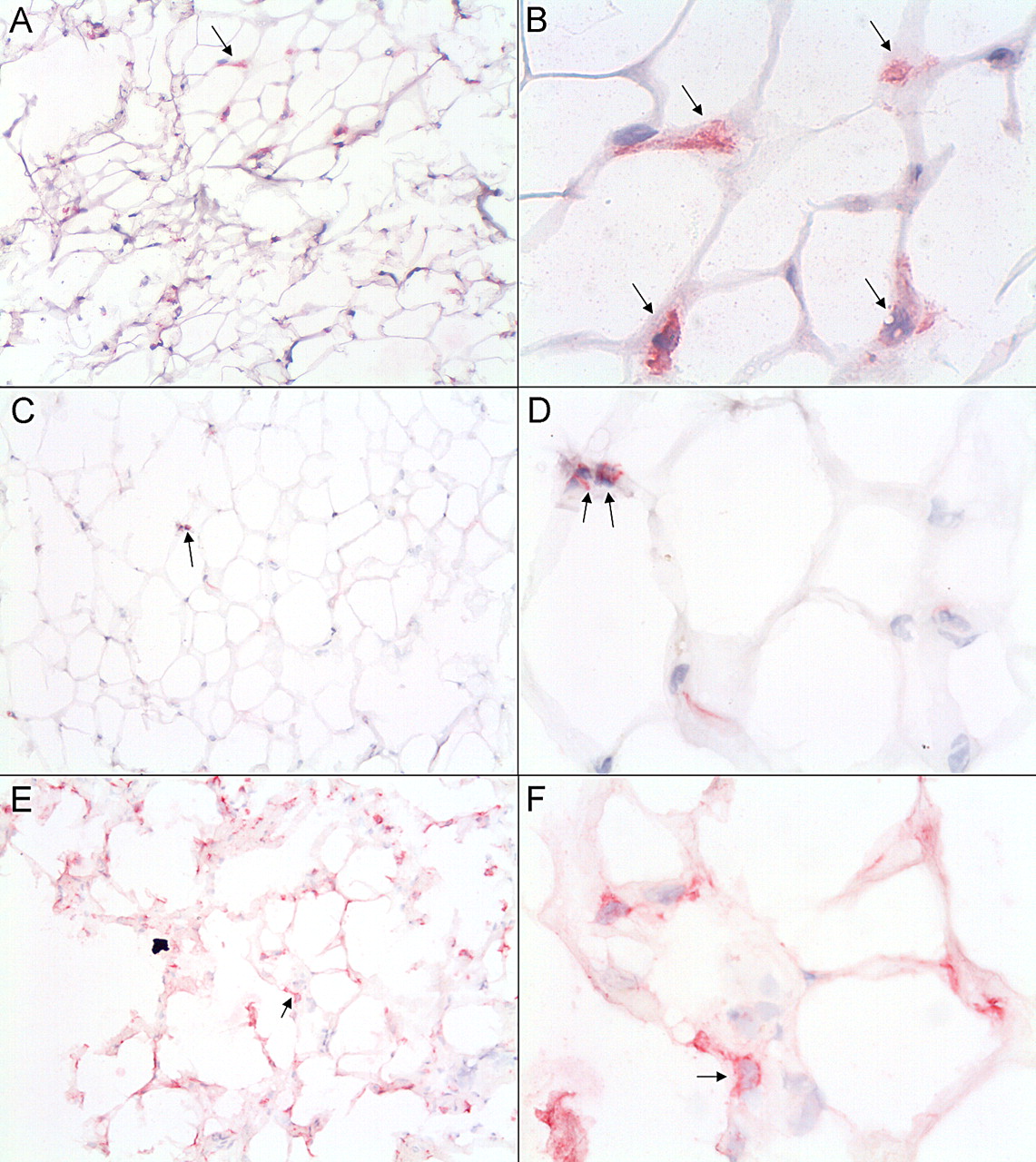
Macrophages in infrapatellar fat of patients with OA. Immunolocalisation of CD68 (A,B), CD86 (C,D) and CD206 (E,F). Positive cells are shown in red and indicated with arrows. Magnification, 100× (A,C and E) and 400× (B,D and F). OA, osteoarthritis.
Comparing IPFP and subcutaneous adipose samples from matched donors (n=7) revealed no difference regarding the presence of CD68+, CD86+ and CD206+ cells (figure 4A). In a separate analysis, IPFPs obtained from patients undergoing a reconstruction of their ACLs (n=5) were compared with BMI-matched OA IPFP samples. The ACL IPFP samples were ranked lower for CD206 (p=0.003 vs OA IPFP) (figure 4B).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
The immunohistochemical rank distribution of CD68, CD86 and CD206 in OA IPFPs and paired subcutaneous fat samples (Sc fat) obtained from patients undergoing total knee replacement (both n=7, mean age 64.8 years (range 54–81 years) and mean BMI 31.7 (range 23.5–48.5)). (B) The immunohistochemical rank distribution of OA IPFPs (n=12, mean age 65.6 years (range 55–78 years) and mean BMI 25.1 (range 22.8–27.0)) with BMI-matched IPFP samples obtained during ACL rupture repair (ACL IPFP, n=5, mean age 37.4 years (range 24–58 years) and mean BMI 23.5 (range 21.8–27.1)). p values indicate a significant difference. ACL, anterior cruciate ligament; BMI, body mass index; IPFP, infrapatellar fat pad; OA, osteoarthritis.
The above-mentioned observations were all made on histological samples. To quantify the presence of CD206 and CD86 cells in OA IPFP, we evaluated 11 IPFP samples with FACS (fluorescence-activated cell sorting) analysis. For this analysis, we used IPFP samples from patients with end-stage OA other than those examined in the earlier experiments, since fresh IPFP samples were necessary. Of the CD45+ cells, 28.9±22.9% were CD86+CD206− and 17.5±13.6% were CD86−CD206+ (p=0.375). Double-positive CD206+CD86+ macrophages were also detected, on average 21.8±20.9% (see supplementary figure 2).
Discussion
OA is a disease of the articular joint, characterised by cartilage damage with an unclear role for inflammation in aetiology and disease progression. However, data suggesting that OA is an inflammatory disease where cytokines and immune cells play a role are accumulating.28 Adipose tissue in general can be considered as an endocrine organ secreting cytokines and growth factors and with significant infiltration of immune cells including macrophages.4 19 24 In this study, we explored the contribution of the IPFP obtained from joints of patients with OA on cartilage metabolism. Earlier publications suggest that IPFP contributes to cytokine and growth factor levels in the synovial fluid8,–,10 and that IPFP from patients with end-stage OA has a different secretory profile than subcutaneous adipose tissue.7 However, the effect of IPFP on cartilage was never investigated before. Here, we report that medium conditioned by IPFP can indeed influence cartilage metabolism.
In contrast to our hypothesis that conditioned media from OA IPFP would induce catabolic changes in cartilage, we observed a decrease in catabolic processes in the cartilage explants as demonstrated by a decrease in NO and GAG release and MMP1 expression when incubated with FCM alone. Although the decrease in GAG release was not supported by changes in gene expression of ADAMTS4 or ADAMTS5, it is well known that aggrecanase activity is regulated by altered activation or post-translational protein production.29 Aggrecan and collagen gene expression were unaltered, indicating no direct effects on production of the main matrix components. Although healthy cartilage has a turnover involving matrix degradation, the expression of some catabolic factors was very low such that addition of FCM could not inhibit it. Therefore, catabolic processes were stimulated by the addition of IL-1β. FCM was able to inhibit the IL-1β-stimulated NO production, MMP1 and MMP3 gene expression and reverse the IL-1β-induced decrease in collagen type II gene expression. Taken together, these findings suggest that osteoarthritic IPFP and its secreted factors inhibit catabolic processes in cartilage. The results might have been clearer if we would have only selected primary end-stage OA samples. However, primary and secondary OA are different regarding the onset of the disease, but it has never been proven that the end stage is different. In this study, medium conditioned by IPFP should be considered as a black box since many known and unknown factors might have contributed to the effects we observed. Future studies using inactivating antibodies or scavenger receptors might identify possible factors or interactions responsible for the inhibition of catabolic processes.
We hypothesised that FCM would have a catabolic effect on cartilage, and therefore, we chose to use healthy bovine cartilage. Bovine cartilage is a frequently used and accepted system to study the effect of growth factors and cytokines on cartilage.26,30,–,32 We needed a large amount of cartilage and preferred to prevent heterogeneity in metabolic status that is known to happen in human osteoarthritic cartilage. Based on the results of our studies, we believe we have indications that factors secreted from end-stage osteoarthritic infrapatellar fat can inhibit catabolic processes in cartilage. However, we cannot exclude the fact that factors secreted by infrapatellar fat into the joint cavity in patients have different effects because of the complexity of the joint, including the presence of synovium and bone, the influence of joint mechanics and the difference between human and bovine cartilage.
We examined whether the phenotype of macrophage (M1 characterised by CD86 or M2 characterised by CD206) might explain the anticatabolic effect of FCM on cartilage. Based on immunohistochemistry, BMI-matched osteoarthritic IPFP samples contained more anti-inflammatory M2/CD206+ macrophages than IPFPs obtained from patients with ACL rupture. The presence of CD206+ macrophages in the OA IPFP supports the finding that FCM from end-stage OA IPFP exhibits anticatabolic effects on cartilage. CD206+ or M2 macrophages express, among many other molecules, arginase,33 an enzyme that competes with NO synthase 2 activity and thereby might inhibit NO production.34 The presence of these arginase-expressing macrophages in the IPFPs might have contributed to the anticatabolic effects of FCM on cartilage. The comparison between OA IPFP and ACL IPFP underlines the more acute inflammation state of the ACL IPFP, since the latter contained less CD206+ cells. Unfortunately, we obtained sufficient amounts of these adipose tissue types only for immunohistochemistry and not for the preparation of FCM, and thus, their effects on cartilage could not be studied. In addition, we were not able to obtain IPFPs from healthy subjects for immunohistochemical comparison.
Subcutaneous adipose tissue of lean people mainly contains M2 macrophages,35 36 and when people gain weight, a shift from M2 to M1 macrophages is seen.37 38 In our study, the BMI of the OA IPFP donors was not associated with the presence of CD68+, CD86+ or CD206+ cells. The effect of the FCM on cartilage was also independent of the BMI of the IPFP donor. In a recent study, OA IPFP produced more IL-6 than subcutaneous adipose tissue from the same donor, although there was no difference in the presence of CD14+ and CD68+ (both used as markers for macrophages) cells between IPFP and subcutaneous adipose tissue.10 This suggests that IPFPs from OA knees may have specific phenotypic characteristics as supported by the presence of different types of macrophages and their secreting factors and that IL-6 production by the IPFP in knee OA is probably not related to a general phenotype in obesity but rather to a specific characteristic of the IPFP. In our previous study,7 a relation between TNFα secretion and the BMI of the IPFP donor was found. This TNFα was produced by CD3+/CD4+ cells, CD3+/CD8+ cells and CD14+ cells. In the current study, we show that the presence of CD68+, CD86+ and CD206+ cells is not associated with BMI.
FACS analysis showed a variation in the presence of pro-inflammatory CD86 macrophages and anti-inflammatory CD206 macrophages between the 11 different donors. This might relate to different subtypes of OA or to different stages in the OA process, earlier observed for synovium,39 which deserves further research. Many double-positive cells were also observed underlining the fact that the division between M1 and M2 macrophages is a gliding scale as proposed earlier.40 41 Further experiments are warranted to elucidate whether our observation is only due to products released by macrophages or that the macrophages interact with adipocytes or other cells, resulting in an altered release profile of the IPFP. Next to macrophages, other immune cells are present in adipose tissue albeit in smaller amounts,7 which might also contribute to the interaction between IPFP and cartilage.
To our knowledge, this is the first study to examine the effect of adipose tissue on cartilage and the potential involvement of the IPFP on cartilage metabolism and activity. This study indicates that the IPFP of patients with end-stage OA has an anticatabolic effect on cartilage and not a pro-catabolic effect originally hypothesised by us. This could be due to a compensatory mechanism in order to prevent further cartilage damage. Additional experiments are required to validate these results and examine them in more detail in human cartilage, to examine the effect of FCM on the entire osteoarthritic process and to investigate whether the effect we found is specific for the IPFP of patients with end-stage OA or whether this is a general (OA) IPFP or phenomenon. This knowledge might eventually contribute to the treatment or prevention of OA and guidelines for whether or not to remove the IPFP during total knee replacement or during the early onset of OA.
Acknowledgments
This study was performed within the framework of the Dutch Top Institute Pharma project # T1-213. Stefan Clockaerts was financially supported by a research mandate of the University of Antwerp.
References
Supplementary materials
Web Only Data
Files in this Data Supplement:
Footnotes
-
Competing interest None.
-
Provenance and peer review Not commissioned; externally peer reviewed.