Article Text

Abstract
Background: Proteoglycan aggrecan (PG)-induced arthritis (PGIA) is the only systemic autoimmune murine model which affects the axial skeleton, but no studies have been performed characterising the progression of spine involvement.
Objectives: To follow pathological events in experimental spondylitis, and underline its clinical, radiographic, and histological similarities to human ankylosing spondylitis (AS); and to determine whether the spondyloarthropathy is a shared phenomenon with PGIA, or an “independent” disease.
Methods: Arthritis/spondylitis susceptible BALB/c and resistant DBA/2 mice, and their F1 and F2 hybrids were immunised with cartilage PG, and radiographic and histological studies were performed before onset and weekly during the progression of spondylitis.
Results: About 70% of the PG immunised BALB/c mice develop spondyloarthropathy (proteoglycan-induced spondylitis (PGISp), and the progression of the disease is very similar to human AS. It begins with inflammation in the sacroiliac joints and with enthesitis, and then progresses upwards, affecting multiple intervertebral disks. In F2 hybrids of arthritis/spondylitis susceptible BALB/c and resistant DBA/2 mice the incidence of arthritis was 43.5%, whereas the incidence of spondylitis was >60%. Some arthritic F2 hybrid mice had no spondylitis, whereas others developed spondylitis in the absence of peripheral arthritis.
Conclusions: The PGISp model provides a valuable tool for studying autoimmune reactions in spondylitis, and identifying genetic loci associated with spondyloarthropathy.
- AS, ankylosing spondylitis
- IVD, intervertebral disk
- PG, proteoglycan aggrecan
- PGIA, PG-induced arthritis
- PGISp, PG-induced spondylitis
- SpA, spondyloarthropathy
- animal model
- ankylosing spondylitis
- autoimmunity
- proteoglycan aggrecan
- mouse
Statistics from Altmetric.com
- AS, ankylosing spondylitis
- IVD, intervertebral disk
- PG, proteoglycan aggrecan
- PGIA, PG-induced arthritis
- PGISp, PG-induced spondylitis
- SpA, spondyloarthropathy
Ankylosing spondylitis (AS) is a chronic inflammatory autoimmune disease of the axial skeleton, and considered to be a “prototype” of the spondyloarthropathies (SpAs). AS frequently shows familial aggregation with male preponderance, and the association of AS with the human leucocyte antigen (HLA)-B27 as strong evidence of its autoimmune aetiology, was first described more than 30 years ago.1 The combination of HLA-B27 with other HLA alleles (HLA-B60 and HLA-B35) was found to increase the genetic predisposition up to sixfold,2,3 and genome-wide screening studies suggested that AS has a polygenic character.4,5 Despite intensive research, the pathological mechanism of AS is unknown. Studies on putative autoantigens implicated molecular mimicry, represented by Klebsiella antigens,6–9Yersinia antigens,10–12 self recognised HLA-B27,11,13,14 or epitopes in the cartilage proteoglycan aggrecan (PG)15–19 in AS.
Animal models are invaluable aids in the research of human autoimmune diseases, and there are only a few systemic animal models, mostly in genetically altered rodents, which involve intervertebral disk (IVD) pathology.20–27 Among the rodent models of the SpAs, immunisation of susceptible mice (BALB/c and some C3H substrains) with human cartilage PG,28–30 or with the G1 domain of aggrecan or versican,31,32 induces progressive polyarthritis which is frequently associated with spondylitis resembling human AS. Although spine involvement in PG-induced arthritis (PGIA) has been known since the murine model was first described,28,33 no systemic study has been performed to determine whether it is only a concomitant phenomenon of peripheral arthritis, or whether arthritis and spondylitis represent two different diseases; although both are induced by immunisation with cartilage PG. Here we describe a longitudinal study on the spine during the progression of PGIA in BALB/c mice. We also show a clear divergence of the two models (arthritis and spondylitis) in F2 hybrids of arthritis susceptible (BALB/c) and arthritis resistant (DBA/2) strains of mice.
MATERIALS AND METHODS
Antigens, animals, experimental groups, and immunisation
The use of human cartilage from joint replacement operations for antigen isolation was approved by the Institutional Review Board, and all animal experiments were approved by the Institutional Animal Care and Use Committee. Human cartilage PG was used for immunisation of 24–26 week old female BALB/c mice purchased from the National Cancer Institute (NCI, Kingston Colony, NY). Preparation of cartilage PG was carried out as described earlier.34,35 As a standard method,30,34,35 the first antigen injection (100 μg PG protein) was given in complete Freund’s adjuvant (Difco, Detroit, MI), and the same doses of antigen were injected as second and third boosts in incomplete Freund’s adjuvant on weeks 3 and 6 (n = 210). Control female BALB/c mice matched for age were either not immunised (n = 75) or immunised with ovalbumin, a non-cartilage-related antigen in adjuvant (n = 75). A small group of PG immunised male mice (n = 12) was used as a positive control, and ovalbumin plus adjuvant injected BALB/c male mice (n = 8) as a negative control group, and all were killed at the end of the experiments at week 36 (fig 1).
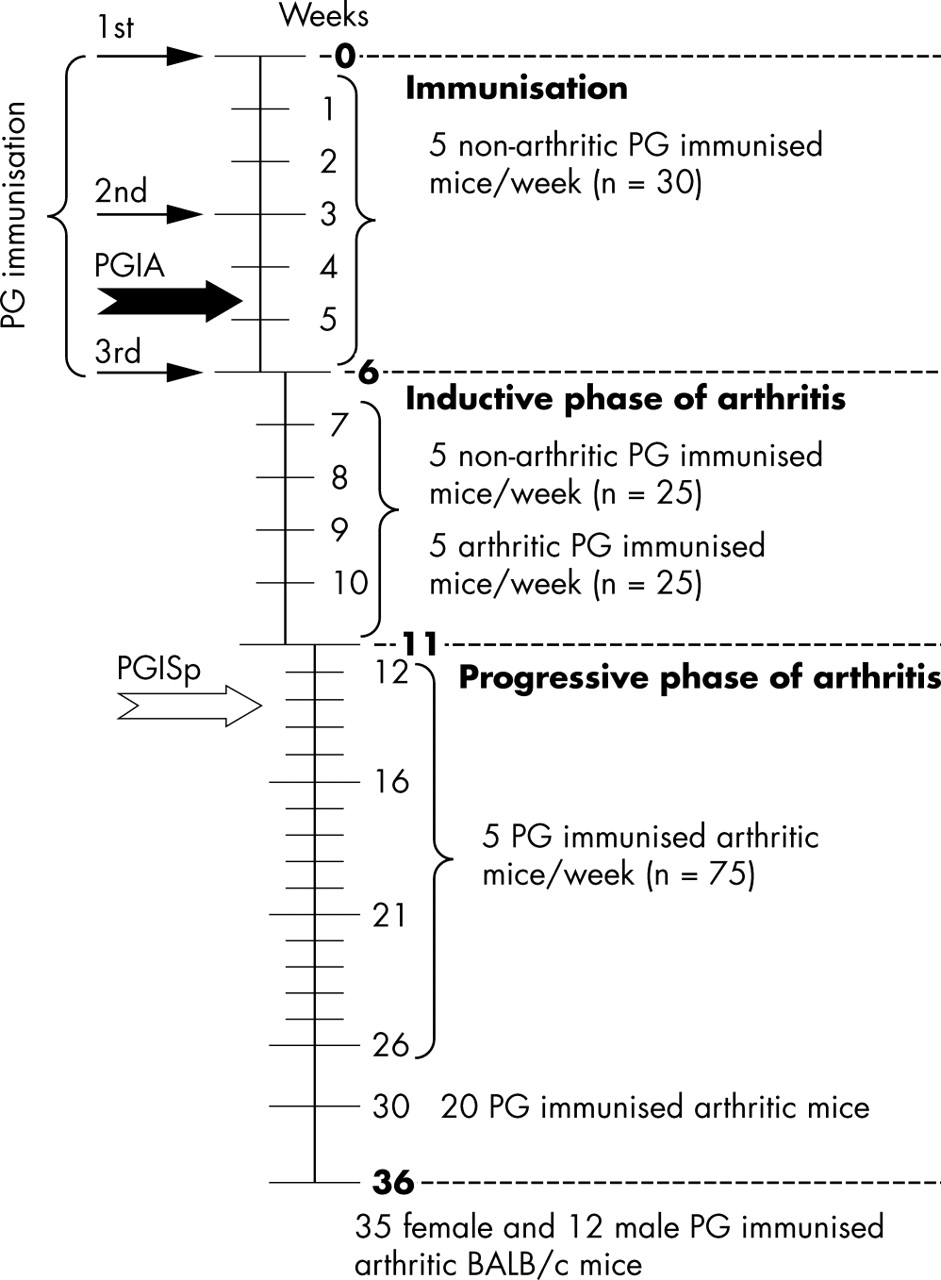
Time course of immunisation, number of animals killed and analysed weekly, and the time of onset of arthritis in peripheral joints (PGIA; thick black arrow) and spondylitis (PGISp; thick white arrow). A total of 210 female BALB/c mice were immunised with human cartilage PG in the first experimental group, and five killed weekly.
In a second set of experiments, arthritis and spondylitis susceptible BALB/c female mice28,30,33,34 were mated with arthritis resistant DBA/2 male mice and the resulting F1 offspring were mated to generate F2 hybrids. These F1 (36 female mice and 12 male mice) and all F2 hybrids (106 female mice and 117 male mice) of BALB/c × DBA/2 intercrosses were also immunised with cartilage PG as described above.29,34,36
Clinical and histological assessments of arthritis and spondylitis
Immunised BALB/c mice were examined twice a week for clinical symptoms of arthritis. A standard scoring system, based upon swelling and redness of each paw (a maximum score of 4 for each paw) was used for the assessment of arthritis.30,34,35 During the first 6 weeks of immunisation (until the third injection), we killed five immunised mice every week. During the next 5 weeks, as arthritic mice became available (fig 1; inductive phase of arthritis), we killed five arthritic and five as yet non-arthritic mice each week (fig 1). From week 12 (that is, 6 weeks after the third injection), when eventually all PG aggrecan immunised BALB/c mice were arthritic, except for three female mice (table 1), we killed five mice weekly until week 26, and then 20 arthritic mice were killed at week 30. The remaining 32 arthritic and three non-arthritic PG immunised female mice, and all male mice, were killed on the last day (week 36) of the experimental period (fig 1).
Incidence of arthritis and spondylitis in cartilage PG immunised mice
In the second set of experiments, PG immunised BALB/c (n = 81), DBA/2 (n = 48), and their F1 (n = 48) and F2 (n = 223) hybrid offspring were scored weekly, and onset and severity of arthritis were recorded as described above for BALB/c mice. Animals were killed on week 30 and disease onset and severity were correlated with the final histological score of the spine.
Serial sections were stained with haematoxylin-eosin, safranin O-fast green, and picrosirius red.37,38 Spines of control, and age matched PG immunised arthritic BALB/c mice were stained with alizarin red and alcian blue for gross pathological examination, as described elsewhere.39
A histology scoring system was established for the severity of spine involvement: score 1, enthesitis, inflammatory cell accumulation around the IVD and/or infiltration of the annulus fibrosus; score 2, <50% absorption/erosion of the IVD; score 3, essentially complete resorption (>50%) of the IVD; score 4, cartilaginous/bony ankylosis. A minimum of 18–22 IVDs (from the distal cervical to the lumbar regions) of each control and immunised mouse was scored. Finally, the spondylitis score of each animal was calculated by dividing the total score (all IVD scores) by the number of IVDs examined histologically. Autoimmune reactions (antibody production and T cell responses) to immunising human and mouse (self) PGs were determined as described previously.28,34,35,40
RESULTS
Development of peripheral arthritis, and PG-specific antibody and T cell responses
As described earlier28,30,34 and above (experimental groups), most of the PG immunised BALB/c mice developed peripheral arthritis between days 7 and 14 after the third antigen injection (table 1). All arthritic mice showed strong humoral and cellular immune responses to PG, as described in detail in our earlier papers.30 In this respect, we found no differences between mice with and without spine involvement, and no correlation between the severity of spondylitis and PG-specific antibody levels. In a similar way to that described above for antibody responses, T cell reactions in PG immunised BALB/c mice with and without spondylitis were highly comparable, and neither PG-specific antibodies nor T cell responses were detected in non-immunised or ovalbumin/adjuvant immunised mice.
Histological assessment of spine involvement in PG immunised BALB/c mice
Histological analysis showed axial involvement as early as week 8 after the third PG injection (data not shown). The disease affected only the PG immunised mice, and abnormalities could not be detected in any of the control animals. Cartilage surface erosions were found first in the sacroiliac joints (fig 2B); IVDs of the proximal tail became affected simultaneously with, or shortly after, the development of inflammation in the sacroiliac joint. During the course of the disease, first the lumbar, and later the proximal, thoracic and distal cervical segments became involved, but not all IVDs were equally affected at a particular time. The fibrocartilaginous articular surface of the sacroiliac joints became eroded (fig 2B), which was followed by a focal (fig 2C) and then more extensive (fig 2D) chondrocyte proliferation. This reactive cell proliferation gave rise to chondrophyte-like tissue; the sacroiliac joint was completely ankylosed within 1–2 months after the first x ray examination (not shown) and histological abnormalities could be seen (fig 2D).

Inflammation of the sacroiliac joints and progression of ankylosis in PG immunised BALB/c mice with spondylitis. The normal sacroiliac joint (A), which is first eroded by a synovial pannus-like hyperproliferative tissue (black arrows) at the articular surfaces indicated with dotted lines in (B), as early as 4–6 weeks after the onset of arthritis in the peripheral joints. This was followed shortly by massive proliferation of chondrocyte-like cells (white arrowheads in (C), which formed a junction of chondrophytes, resulting in complete ankylosis of the sacroiliac joint (D). The tissue sections were stained with haematoxylin and eosin.
In the acute phase of spondylitis, strong mononuclear infiltration appeared,28–30 leading to destabilisation of the IVD. The damaged or resorbed annulus fibrosus allowed the nucleus pulposus to protrude in a ventral direction. The protruding IVD or the growing osteophytes frequently caused compression of the spinal cord, particularly at the lower cervical regions (fig 3D).

Cervical spine segments in (A, B, C) normal mice and in (D, E, F) mice afflicted with spondylitis. The affected disks are shown with the ankylosing osteophytes (black arrows: IVD C4/C5), which protrude and compress the spinal cord (SpC) (D). Massive enthesitis develops with a pannus-like tissue eroding the rest of the IVD at the ventral side of the spine. Safranin O/fast green staining (B, E) demonstrates the PG content (red) in the growth plate (GP), in the cartilaginous end plate (EP), and, less intensively, in the annulus fibrosus (AF) and nucleus pulposus (NP) of the normal (B) or spondylitic (E) spine. In the affected spine (E) the annulus fibrosus and the nucleus pulposus have been completely resorbed and the end plate (black arrowheads) is essentially absent (E). The neighbouring vertebral bodies are connected by chondrophytes (black arrows), which ankylose the spine. Polarisation microscopy of sections stained with picrosirius red (panels C and F) indicates that the regularity of collagen fibres, due to parallel orientation in the normal annulus fibrosus (C; white arrows), is completely lost in the annulus fibrosus of the disk affected by inflammation (F).
To study the changes in the extracellular matrix and its components, sections were stained with safranin O to assess the PG content (figs 3B and 3E), and picrosirius red staining was used to determine the orientation of the collagen fibres by polarisation microscopy (figs 3C and 3F). Safranin O staining disclosed that, in addition to the growth plate and end plate of the vertebral body, the nucleus pulposus and the annulus fibrosus also contained large amounts of PG (fig 3B). PG was lost from end plates, but large chondrophytes, and later osteophytes, grew at connecting adjacent vertebral bodies (fig 3E). Polarisation microscopy showed normal lamellar orientation of the collagen fibres in the annulus fibrosus and end plate in normal IVDs (fig 3C); an orientation which was lost in affected IVDs (fig 3F).
Macroscopic analysis of spondylitic spines
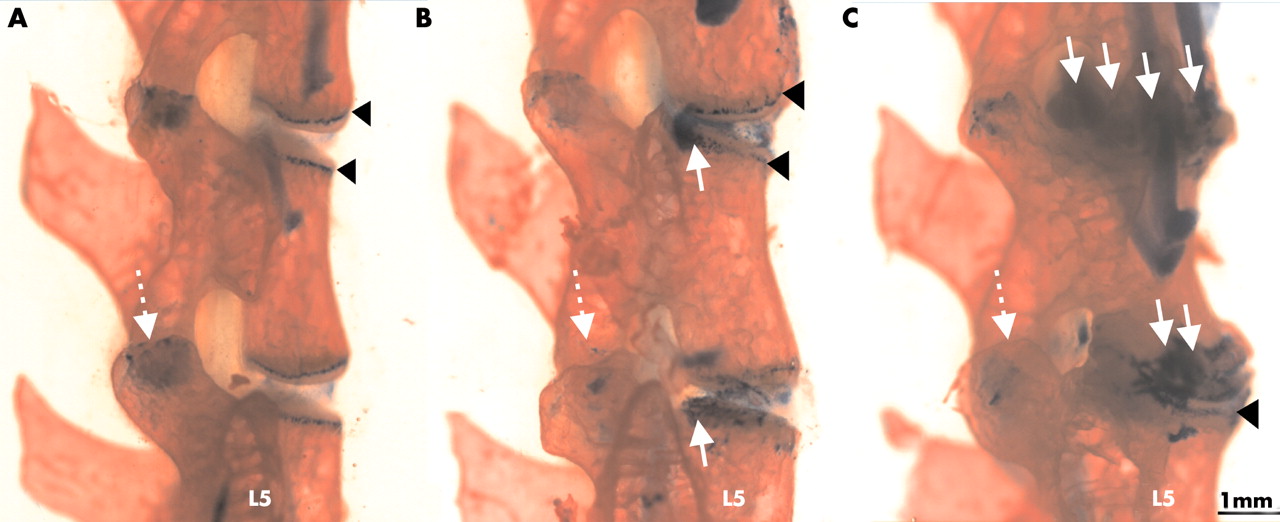
For macroscopic examination, spines were stained with alizarin red and alcian blue (fig 4). This staining shows more details of the cartilaginous and bony components of the deformed spine regions. In contrast with the normal disk, where the growth plates and end plates of the vertebral bodies appeared as sharp solid lines (fig 4A), the contours of these cartilaginous tissues became diffuse owing to chondrophyte formation (fig 4B). Progression of spondylitis was seen in each animal that developed spondylitis: the disk space disappeared and a chondrophyte/osteophyte, fusing the neighbouring vertebral bodies, replaced the original IVD (fig 4C).
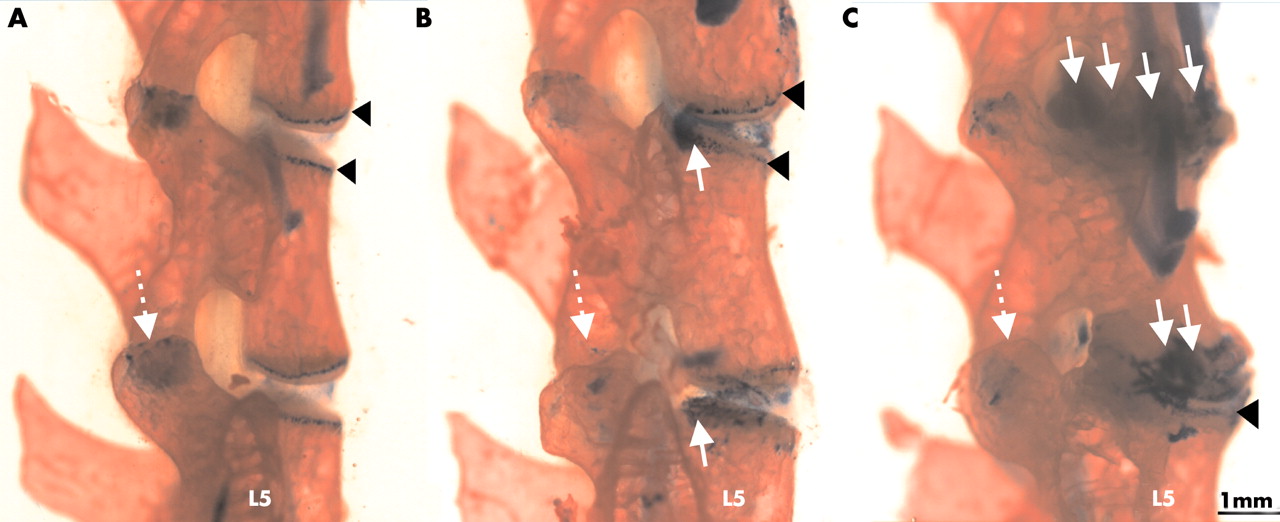
Progression of spondylitis in the lumbar spine (L3-5) in PG immunised BALB/c mice. Alizarin red stains bone red, while alcian blue stains the cartilaginous structures blue. Black arrowheads indicate the cartilaginous growth plates of the vertebral bodies (A; control), which became moderately deformed by 12 weeks (B) and grossly altered by 24 weeks (C) after the onset of peripheral joint inflammation. White arrows show chondrophyte formation at the marginal regions of the vertebral bodies.
Analysis of peripheral arthritis and spondylitis in F2(BALB/c × DBA/2) hybrid mice
As described above, three PG immunised BALB/c female mice did not develop PGIA, but two of the three showed SpA at the end of the experiment (table 1). Therefore, we assume that SpA develops independently of arthritis in PG immunised BALB/c mice. To confirm this hypothesis and separate overlapping clinical, histopathological, and laboratory measures of the two diseases, we immunised 223 F2(BALB/c × DBA/2) hybrids, and their parents. All mice (BALB/c, DBA/2, F1 and F2 hybrids) were killed on week 30 of immunisation, as the development of PG-induced spondylitis (PGISp) achieved a constant (∼70%) level by week 26 of immunisation (table 1). Figure 5A shows the major characteristics and distribution of peripheral arthritis and spondylitis in these experimental groups. About 40% of the PG immunised F2(BALB/c × DBA/2) mice developed PGIA, but unexpectedly, over 60% had spondylitis. Most of the arthritic mice had spondylitis (85/97), and 52/223 (23.3%) PG immunised F2 hybrids had spondylitis without joint inflammation (fig 5A; right panel).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
(A) Analysis and comparison of PG-induced arthritis (PGIA: A) and PG-induced spondylitis (PGISp: Sp) in 81 BALB/c (B/c), 48 DBA/2 (D/2) mice and their F1 and F2 hybrids (BALB/c × DBA/2) immunised with human cartilage PG. All mice were injected four times with PG and killed on week 30 of the experiment—that is, 21 weeks after the last immunisation. (B) Significant differences when male mice and female mice with arthritis or spondylitis, or both, were compared: *p<0.05; **p<0.01.
In this second set of experiments, 81 (63 female and 18 male) BALB/c mice developed PGIA (mean (SD) arthritis score 11.6 (2.4)), which was associated with PGISp (mean (SD) spondylitis score of affected animals 1.45 (0.62)) in 57 (70%) animals.
DISCUSSION
Normal IVD is a complex fibrocartilaginous structure containing poorly defined chondrocyte-like and fibroblast-like cells that can survive in a relatively avascular environment. The extracellular matrix of the IVD contains two major groups of macromolecules: collagens and proteoglycans. Whereas the nucleus pulposus contains almost uniformly aggrecan and type II collagen, the annulus fibrosus is formed mostly by type I collagen fibrils embedded in aggrecan and versican.41 The insertion site of the ligaments into the cortical bone—that is, the enthesis, also contains proteoglycan, mostly versican,42 which might be degraded in IVDs undergoing degenerative processes.43,44
The core protein of human cartilage (and nucleus pulposus) PG comprises more than 2300 amino acids, and contains 20–27 T cell epitopes30 presented to T cells by various major histocompatibility complexes of antigen presenting cells.45 Most of these epitopes, however, are in a “cryptic” position—that is, masked by glucosaminoglycan side chains. A few T cell epitopes of human PG have been characterised as dominant/arthritogenic, and owing to the high sequence homology between human and mouse PGs,46,47 immunisation of susceptible BALB/c and C3H mice with human PG provokes autoimmune responses to mouse (self) PGs, which then induce arthritis and spondylitis.28–30,34
Spondylodiscitis (a robust form of inflammation affecting the entire IVD) occurs in 15% of patients with AS,48 but the incidence is greater (up to 60%) when diagnosed by magnetic resonance imaging.49 The primary site of the inflammation in AS, however, is still debated; it is proposed to be in the subchondrium49 or in the synovium (sacroiliac joint) close to the entheses.50
To date, spontaneous or experimentally induced disk degeneration has been reported only in a few animal models.20–22,24,26,27,51 Autoimmune mechanisms are thought to play a part only in HLA-B27 transgenic rodents23,25,52 and in PGIA.28–30,34 As HLA-B27 transgenic animals, when maintained in germ-free conditions, do not develop spondylitis,25,53 the molecular mimicry of bacterial antigens may be a strong contributing factor to the pathomechanisms of SpA either in these transgenic rodents or in human patients. Nevertheless, none of these animal models is identical with human AS, but may mimic genetic and/or pathological abnormalities present in the human disease. One example is that, in addition to the major histocompatibility complexes, the major genetic loci of the PGISp in F2 hybrids of BALB/c and DBA/2, and BALB/c and C3H intercrosses are localised on chromosome 2 (authors unpublished observation), which corresponds with that identified in patients with AS.5,54 PGIA is characterised by a fine cooperation between antibody producing and antigen presenting B cells and T helper cells,30,55 and affects both the axial and peripheral skeleton. The chronology of pathological events in this model suggests that the involvement of the sacroiliac joint and the spine is similar to AS. PG immunised mice first exhibit strong mononuclear influx and develop pannus-like tissue in the sacroiliac joint. The infiltrated tissue gradually destroys cartilage, which is followed by complete ankylosis of the joint. Simultaneously with, or shortly after, the initial signs of sacroiliitis, spondylitis/spondylodiscitis flares up with mononuclear infiltration around the annulus, followed by destabilisation of the annulus and protrusion of the nucleus. Matrix components from the nucleus could be additional local sources of autoantigens. The abundance of antigens in this environment can initiate self sustaining (auto)immune reactions that ultimately lead to the complete loss of IVD, and reactive chondrophyte/osteophyte formation and ankylosis, fusing together vertebral bodies.
PG is one of the major extracellular matrix proteins located in immunoprivileged sites of the body (such as in the articular cartilage or nucleus pulposus of the IVD), therefore, it might become a target antigen in autoimmune diseases.15–17,19,56–58 Here, we demonstrate that spondylitis develops in arthritis susceptible mice immunised with PG, and the progression of the spine disease resembles that seen in human AS. Therefore, we postulate that PG has an important role in PGISp in this model, and perhaps also has a role in humans with autoimmune SpA.15,16,18,19 As far as we know, this is the first time that a systematic and longitudinal study has been performed on spine disease in mice with systemic immunisation with an autoantigen (cartilage PG). We found that spondylitis developed without arthritis, especially in PG immunised F2 hybrids of PGIA susceptible BALB/c and resistant DBA/2 strains. Vice versa, about 30% of arthritic mice did not show evidence of spondylitis. These observations strongly suggest that PGIA and PGISp are independent diseases, although immunisation with cartilage PG is an absolute requirement for the induction of either PGIA or PGISp.
In conclusion, it is very likely that there are both overlapping/shared and distinct genetic components of the (auto)immune regulation in these two models. Although the autoimmune pathomechanisms of PGIA and PGISp seem to be similar, the incidence, severity, and particularly the genetic control of joint and spine involvement are likely to be distinct. Using F2 hybrids of arthritis and spondylitis resistant DBA/2 and susceptible BALB/c mice, we showed a maximum of 57% (85/149 affected animals) overlap between PGIA and PGISp, whereas 40% of the F2 hybrid mice with spondylitis did not have arthritis. Therefore, this report supports the relevance of performing genome-wide screening studies for identification of susceptibility loci controlling AS in this experimental murine model, which seems to mimic the human disease.
Acknowledgments
This work was supported, in part, by grants (AR40310, AR45652, AR51163) from the National Institute of Health (USA).