Article Text

Abstract
Objectives To examine the expression of B-cell-activating factor receptor (BAFF-R) on naive CD27− and memory CD27+ B cells in normal individuals and patients with rheumatoid arthritis (RA) undergoing B-cell depletion therapy with rituximab.
Patients and Methods BAFF-R expression on B-cell subsets was determined in normal controls (NC; n=11), active patients with RA pre-rituximab (pre-RX; n=15), relapsing patients either concordant for B-cell repopulation (C-R, n=13) or discordant, with relapse more than 3 months after repopulation (D-R, n=11) and patients in remission over 3 months postrepopulation (discordant non-relapsing (D-NR), n=5). Serum BAFF was measured by ELISA and analysed using Mann–Whitney.
Results There was no significant difference between NC, pre-RX and D-NR patients in %BAFF-R-positive B cells or mean fluorescence intensity (MFI) in naive and memory B cells. Relapsing patients had significantly lower MFI and %BAFF-R-positive cells in both naive and memory compartments from NC and pre-RX (C-R and D-R; p<0.01). BAFF levels in pre-RX patients were within the normal range and did not correlate with BAFF-R expression in any patient group. D-NR patients had relatively lower proportions of pre and postswitch CD27+ B cells than pre-RX patients (D-NR vs pre-RX; p<0.05 for both) and also lower numbers of postswitch B cells than D-R patients (D-NR vs D-R, p<0.05).
Conclusion BAFF-R expression was significantly reduced on both naive and memory B cells in patients at relapse, regardless of the relationship with B-cell repopulation or serum BAFF levels. Re-establishment of active disease was also associated with an increase in class-switch recombination. Factors responsible for lower levels of BAFF-R may relate to altered thresholds for autoreactive B-cell generation at relapse in patients with RA.
Statistics from Altmetric.com
B-cell-activating factor (BAFF), also known as B lymphocyte stimulator (BlyS), THANK, TALL-1, zTNF-4 or TNFSF13b, is a protein member of the tumour necrosis factor (TNF) family that plays an important role in B-cell maturation, homeostasis and survival.1 BAFF binds to three receptors on B cells: TACI, BCMA and BAFF-R (or BR3), with BAFF-R being the only ligand that exclusively binds BAFF.2
Studies of BAFF-R-deficient mice demonstrated that BAFF was essential for B-cell survival, primarily through the alternative nuclear factor κ light-chain enhancer of activated B cells pathway,3 and for B-cell maturation and immunoglobulin (Ig) class switching through the classic nuclear factor κ B pathway.4 B cells from these animals develop normally up to the IgM+ transitional stage but do not complete their maturation in the spleen.5 The numbers of memory and marginal zone cells and of serum Ig are severely reduced (>90%) as were responses to both T-dependent and T-independent stimuli.2
Human B cells are less dependent on BAFF-R signalling for maturation than murine B cells. In the few reported cases of common variable immunodeficiency related to BAFF-R deficiency in humans, circulating BAFF levels were greatly elevated and peripheral B-cell numbers were reduced (fourfold lower). The condition was of late onset and was not associated with severe infections.6 Although mature but not transitional B cells were reduced in these individuals, class-switched memory B cells were detectable. Correspondingly, vaccination studies showed that T-dependent responses appeared to be much less dependent on BAFF-R signalling than T-independent responses. As in BAFF-R−/− mice, IgA responses in these patients were not compromised.
One of the suggested roles for BAFF, based largely on animal experiments, is that increased levels will lower thresholds for the survival of autoreactive B-cell clones, and decrease competition within germinal centre environments.7 8 In humans, elevated levels of serum BAFF have been associated with both the presence and activity of several autoimmune diseases (Sjogrens syndrome, systemic lupus erythematosus (SLE) and idiopathic thrombocytopaenia).9,–,15 In patients with rheumatoid arthritis (RA), the relationship is less clear, with conflicting reports as to whether levels of circulating BAFF are significantly elevated and if there is a relationship with disease activity or titres of autoantibodies.14 16 17
Following treatment of RA patients with rituximab, serum BAFF levels have been shown to rise, reflecting the loss of the majority of peripheral B cells. The correlation of BAFF levels with peripheral B-cell counts is not, however, perfect and it has been suggested that BAFF levels form a more accurate indicator of total B-cell mass.18 Chronically raised BAFF levels have been suggested to exert a negative feedback on BAFF-R expression.19
One of the key initial findings of the clinical response to rituximab was that B-cell return was not always associated with relapse, and that disease could remain in remission for months or even years following B-cell repopulation.20 In patients with RA, rises in autoantibodies, particularly rheumatoid factor (RhF), seem to herald relapse following regardless of whether clinical relapse has been concordant or discordant with B-cell return.21 Serum BAFF levels rose rapidly after rituximab, and then fell with rising peripheral B-cell numbers, although the relationship was not always clear. There was also some evidence for an increased uptake of BAFF if relapse occurred concurrently.17 21
The re-establishment of clinical disease following B-cell depletion therapy relies on engaging pro-inflammatory pathways, which are quiescent in the absence of circulating B cells, and therefore provides a unique opportunity to study the re-emergence of the disease process. As a result of its importance in B-cell survival, particularly for new emigrant and naive B cells, we hypothesised that the balance between BAFF-R expression and soluble BAFF may influence the relationship between returning B-cell populations and the resumption of autoimmunity. With this in mind, we measured serum BAFF levels and BAFF-R expression on B-cell populations to determine whether there was any relationship between serum BAFF levels and BAFF-R on naive and memory B cells at different key time points after rituximab therapy.
Materials and methods
Patients
Blood samples were obtained from 11 normal controls (NC) and 44 consecutive patients with a diagnosis of RA who fulfilled the American College of Rheumatology criteria for RA22 The patients were all attending the Department of Rheumatology at University College London Hospital and were treated with rituximab, on the basis of clinical need. Patients received 2-weekly infusions of 1 g rituximab preceded by 100 mg intravenous methylprednisolone. The study was approved by the hospital ethics committee and all patients gave informed consent before entering the study. The normal range for cluster of differentiation 19+ (CD19+) B cells used by the local pathology laboratory was 0.03–0.40×109/l. Levels less than 0.005×109/l were defined as undetectable.
Detection of B cells
Depletion of B cells in the peripheral blood was deemed to have occurred when CD19+ B cells were undetectable. In all patients, total peripheral blood B-cell depletion was achieved for at least 2 months. B-cell return (B-cell repopulation) was defined as when B cells were again detectable in the peripheral blood (ie, when the CD19+ cell count was ≥0.005×109/l).
Clinical relapse
Clinical relapse after rituximab was based on any return of symptoms of RA plus a rise in C-reactive protein following an original fall of at least 50% in C-reactive during the previous course of B-cell depletion therapy. Patients at clinical relapse were classified as concordant (relapse ≤3 months after repopulation in peripheral blood) or discordant (relapse >3 months after repopulation in peripheral blood).
Peripheral blood mononuclear cells isolation and staining
Peripheral blood mononuclear cells (PBMC) were freshly isolated from 10 ml heparinised whole blood following density gradient centrifugation with Ficoll-Paque Plus (GE Healthcare, Uppsala, Sweden) and stained on the same day of collection. Cells were washed and cellular viability was estimated with Trypan blue (Sigma, St Louis, USA). PBMC (1×106/sample) were incubated with appropriate conjugated antibodies for 15 min at room temperature in the dark and then washed. Erythrocytes were lysed with BD PharmLyse (BD, Biosciences, San Diego, USA) buffer according to the manufacturer's instructions and cells were then fixed with paraformaldehyde 2% for 5 min at room temperature. PBMC were stored in the dark at 4°C until analysed by flow cytometry. A total of 300 000 cells/sample gated in total lymphocytes were acquired with FACSCaliBur (BD). Data were analysed with FlowJo (TreeStar, Stanford University, California, USA). Absolute cell counts were calculated from the differential leucocyte count determined at each time point for all patients.
Phenotypic analysis
Immunophenotyping of B and T cells in PBMC samples was performed using matched combinations of antihuman murine monoclonal antibodies conjugated to fluorescein isothiocyanate (FITC), phycoerythrin (PE), peridinin chlorophyll protein cyanin (PerCP-Cy5.5), or allophycocyanin (APC). For B-cell analysis, combinations of anti-CD19 conjugated to PerCP-Cy5.5 or APC (clone HIB19), anti-IgD FITC (clone IA6-2) and anti-CD27 PE (clone O323) were used to define naive (CD19+IgD+CD27−), preswitch memory (CD19+IgD+CD27+), postswitch memory (CD19+IgD−CD27+) and IgD−CD27− B cells.
The expression of BAFF-R was analysed using combinations of anti-CD19 PerCP-Cy5.5, anti-CD27 PE and anti-BAFF-R FITC (11C1; mouse IgG1, clone 8A7) and the mean fluorescence intensity (MFI) of BAFF-R was determined for naive (CD19+CD27−) and memory B cells (CD19+CD27+). All antibodies used were purchased from BD Biosciences and eBioscience (San Diego, USA)
Measurement of BAFF
BAFF levels were quantified in serum samples from healthy controls and from patients at the various time points, using the Human Quantikine BAFF/BLyS immunoassay ELISA kit. The mean±SD for normal sera (n=36) were given as 1.17±0.28 ng/ml (range 0.67–2.45 ng/ml).
Statistical analysis
Statistical differences were determined using the non-parametric Mann–Whitney test using GraphPad Prism (GraphPad, San Diego, California, USA). Differences were considered statistically significant for p<0.05. Linear regression was used to correlate BAFF levels and BAFF-R expression.
Results
Patient cohorts
The mean age was 42 years (range 33–81) and the mean disease duration was 15 years (range 2–20). All patients had erosive disease. Two patients were seronegative for both RhF and anticitrullinated protein antibodies before treatment. The cohorts of consecutive patients with RA included 15 patients with active disease who were studied pre-rituximab (pre-RX) and 29 patients who had already received one or more courses of rituximab. In consecutive patients presenting at clinic, blood for FACS analysis was taken from all relapsing patients. CD19 counts were subsequently examined and patients assigned to concordant relapsing (C-R) or discordant relapsing (D-R) groups. If the patients had already repopulated according to the laboratory notes, but had remained well for more than 3 months after the date of repopulation, they were assigned to the discordant non-relapsing (D-NR) group. Five groups were therefore described for analyses: (1) NC (n=11); (2) patients with active RA pre-RX (n=15); (3) C-R (n=13); (4) D-R (n=11); (5) D-NR (n=5), taking into account repopulation and relapse after rituximab as described (table 1).
Serum BAFF levels and BAFF-R expression on CD19+ B cells
Serum BAFF levels in NC and in RA patients
Median BAFF levels in the 16 patients with RA tested before rituximab were within the normal range (table 1). As previously described, BAFF levels rose following rituximab.22 Levels were found to remain raised at relapse in both patient groups (C-R and D-R; p<0.01, compared with both NC and patients pre-RX) and to a lesser extent in patients who had repopulated but not relapsed (D-NR; p<0.05) when compared with NC only (table 1), but no statistically significant differences in BAFF levels were observed between C-R, D-R and D-NR patients.
Percentage of naive and memory B cells expressing BAFF-R
The percentages of CD19+ B cells expressing BAFF-R in CD27+ and CD27− subpopulations in patients and NC are represented in table 1. Significant differences were seen between the percentage expression of BAFF-R on naive and memory B cells in relapsing patients (C-R and D-R groups) compared with NC, and for the memory B-cell population, levels in relapsing patients were also significantly reduced compared with pre-RX values. In the D-R group, decreased expression on naive B cells was also significant compared with pre-RX values. In patients whose B cells had returned but had not yet relapsed (D-NR), no such decreases were found between the percentages of BAFF-R expressing naive or memory populations compared with NC or with the pre-RX group. Furthermore, when the percentages of BAFF-R expressing naive and memory B cells in patients who were relapsing (both C-R and D-R) were compared with D-NR patients, a significantly lower percentage expression was found in both naive and memory populations (naive D-NR vs C-R and D-R; p<0.05 and p<0.01, respectively; memory D-NR vs C-R or D-R, p<0.01 for both). The low expression of BAFF-R was not caused by contamination of the CD19+ gate with plasmablasts (which express low levels of BAFF-R) as they were gated out of the analysis.
MFI of BAFF-R expression on naïve and memory B cells
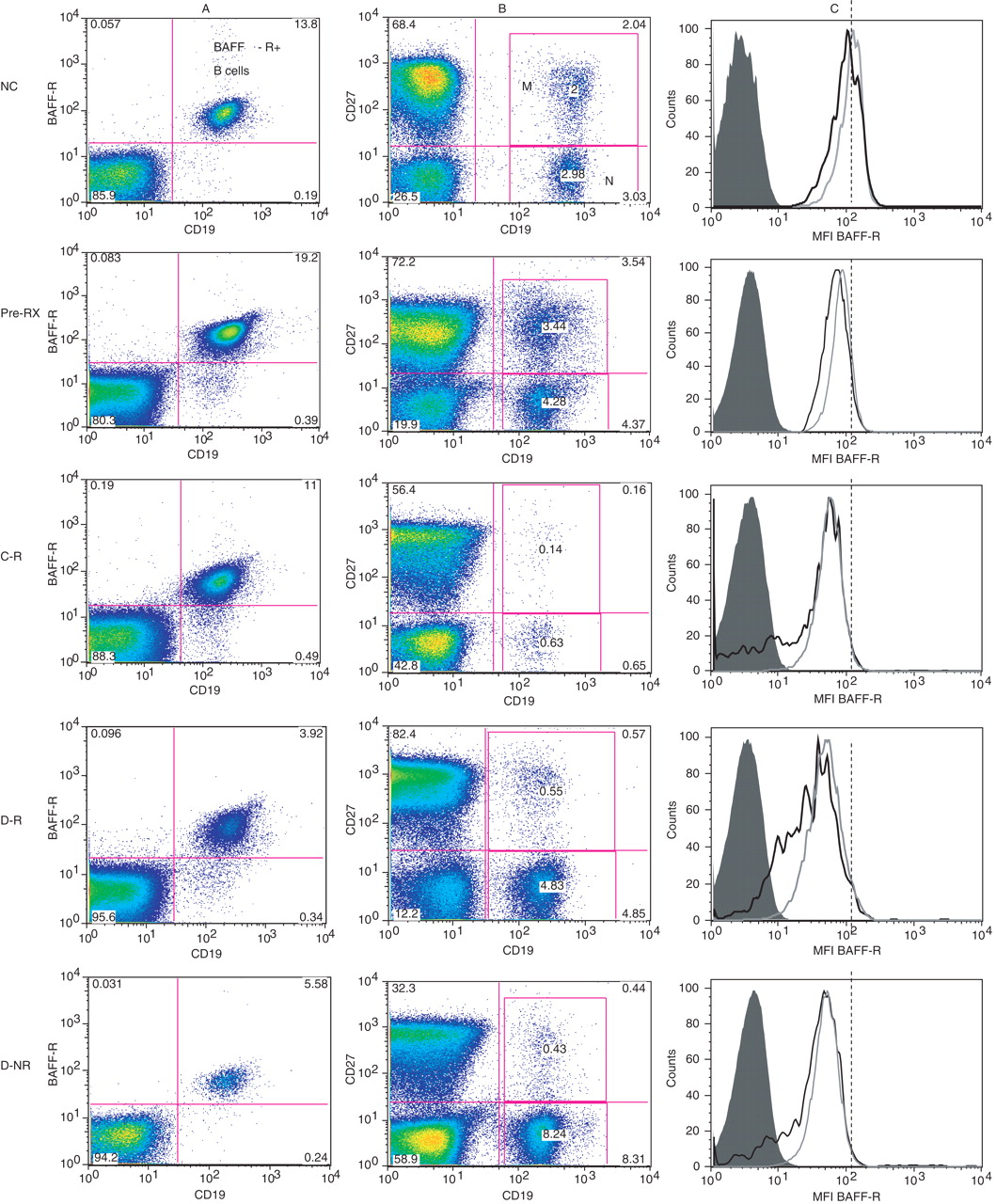
Figure 1 shows representative dot plots and histograms of BAFF-R expression on CD19+ B cells (panel A), PBMC differentiated into naive and memory B cells on the basis of CD27 expression (panel B) and histograms of the MFI of BAFF-R on naive (dashed lines) and memory (dotted lines) B cells (panel C). BAFF-R expression appeared to be decreased in patients who were discordant for relapse after B-cell return, both when remaining well or relapsing, in the examples shown. These individuals also had a tendency for a higher proportion of naive to memory B cells (panels A and B). A left shift in MFI of BAFF-R expression was also observed in all naive and memory populations following rituximab compared with those of NC (panel C). In order to explore these initial observations, the MFI of BAFF-R expression on naive and memory CD19+ B cells pre-RX were compared with those of B cells from NC (figure 2). Naive B cells from both groups of patients at relapse (C-R and D-R) expressed significantly lower levels of BAFF-R when compared with NC (MFI; C-R 58.8 and D-R 53.7 vs NC, 85.5). Memory B cells from both cohorts of relapsing patients also showed significantly lower expression of BAFF-R than NC (MFI; C-R 56.5 and D-R 46.7 vs NC, 78.0). Furthermore, C-R patients showed a tendency to lower MFI levels than pre-RX patients in both naive (58.8 vs 70.4, p=0.06) and memory B-cell populations (56.5 vs 61.2; p=0.07). We also observed a tendency towards lower BAFF-R MFI in D-R patients when compared with pre-RX levels in memory B cells, although again, this did not reach statistical significance (46.7 vs 61.2; p=0.07). Of interest, no significant differences were found in the MFI of BAFF-R expression on either naive or memory B-cell populations in the D-NR patient group when compared with NC or RA patients before treatment.

Peripheral blood mononuclear cells were gated on the CD19+ B-cell population and the mean fluorescence intensity (MFI) of B-cell-activating factor receptor (BAFF-R) expression determined on CD27-naive (A) and CD27+ memory (B) B-cell subpopulations. Normal controls (NC) and patient groups were compared using the Mann–Whitney U test and statistical significance indicated (*p<0.01; **p<0.001). C-R, concordant clinically relapsing; D-NR, discordant clinically non-relapsing; D-R, discordant clinically relapsing; pre-RX, prerituximab.

Characterisation of B-cell-activating factor receptor (BAFF-R) expression and B-cell phenotypes in normal controls (NC) and patients with rheumatoid arthritis before B-cell depletion therapy (pre-RX), relapsing patients concordant (C-R) or discordant (D-R) with B-cell return and patients who had repopulated in the periphery but who remained well (D-NR). The expression of BAFF-R on CD19+ B cells in the lymphocyte gate in peripheral blood mononuclear cells are shown in panel A. In panel B, combinations of CD27 and CD19 were used to differentiate memory (M; CD19+ CD27+) and naive (N; CD19+ CD27−) B cells. Numbers in each quadrant represent percentages of each subpopulation in individual experiments. The histograms in panel C show representative experiments in which the mean fluorescence intensity (MFI) of BAFF-R expression for CD27+ (dotted lines) and CD27− (dotted lines) are shown. Dotted lines depict the MFI of BAFF-R expression on the naive B-cell (CD27−) population from the normal individual to enable comparison with other histograms in each example.
Relationship between BAFF levels and BAFF-R MFI in naive and memory B cells
Our initial observation was that BAFF-R expression was significantly lower on B cells from patients undergoing relapse than on B cells from patients whose B cells had returned but in whom disease was not yet re-established (figure 2). We therefore examined whether there was any relationship between circulating BAFF levels and expression (MFI) of BAFF-R (figure 3). Although no statistically significant correlations were found, we could distinguish different patterns between pre-RX, C-R, D-R and D-NR patients. When patients were relapsing (C-R and D-R), BAFF-R MFI was consistently lower, irrespective of serum BAFF levels. Nevertheless, in patients who had repopulated but remained well (D-NR), there was a tendency for a negative correlation between circulating BAFF and BAFF-R MFI more evident in memory B cells (r2=0.88; p=0.06) (figure 3). However, samples from only four patients were available for analysis in this group.

The values for the mean fluorescence intensity (MFI) of B cell-activating factor receptor (BAFF-R) expression in individual patients were plotted against BAFF levels that were measured in serum stored at the same time point (upper limit of normal range 2.45 ng/ml). Regression analysis was performed and significance values determined for each curve. C-R, concordant clinically relapsing; D-NR, discordant clinically non-relapsing; D-R, discordant clinically relapsing; NC, normal controls; pre-RX, prerituximab.
B-cell subpopulations in peripheral blood associated with different patterns of response to rituximab
It has previously been suggested that patients with higher numbers of circulating memory/postswitch B cells, either before treatment induction or at relapse were more likely to have a shorter clinical response to rituximab.23 24 We showed here that memory B cells from relapsing patients in particular tended to express lower levels of BAFF-R (MFI), and that this was so over a wide range of circulating BAFF levels. We therefore examined whether there was any relationship between subpopulations of B cells in patients with different patterns of clinical response/relapse (figure 4). Circulating B-cell (CD19+) subpopulations were defined according to IgD and CD27 expression and classified as naive (IgD+CD27−), preswitch memory (IgD+CD27+), postswitch memory (IgD−CD27+) and double-negative (IgD−CD27−) B cells. The median and range of total numbers of CD19+ B cells in each patient group were for pre-RX 175 (58–472)×106/l, C-R 42 (15–192)×106/l, D-R 61 (13–166)×106/l, D-NR71 (21–328)×106/l. The median percentage and interquartile range of each subpopulation are shown for NC and patient groups in figure 4. The distribution of naive to memory B cells in humans is approximately 60:40. The B-cell subpopulation distribution of B cells from patients with RA pretreatment was broadly similar to NC, except for a slightly increased proportion of double-negative B cells (7.3% vs 3.9%; p=0.04). Following rituximab, repopulation of the periphery begins with naive B cells, which mature at a greater or lesser rate into pre and postswitch CD27+ B cells. Full restoration of memory cell populations may not be achieved for years after treatment. As expected, the relative ratio of naive to memory B cells was increased at repopulation, mirroring ontogeny, but the relative percentage of naive cells was only significantly higher in the non-relapsing patients compared with NC (p<0.05). The preswitch memory B-cell population was significantly decreased in all patients postrituximab (C-R 4.2%, p=0.0004; D-R 4.9%, p=0.002 and D-NR 4.7%, p=0.03) when compared with NC (12.2%). In patients who relapsed at B-cell return, postswitch memory B cells had already reached levels approximating those of NC (C-R 12.4% vs NC 19.6%, p=0.28), but remained reduced in the D-NR group when compared with NC (6.6% vs 19.6% p=0.03), pre-RX (6.6% vs 18.1%, p=0.02) and D-R (6.6% vs 17.9%, p=0.05).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Peripheral blood mononuclear cells were gated on CD19 to characterise B-cell subpopulations further on the basis of IgD and CD27 expression into naive, pre and postswitch memory and double-negative B cells. Significance levels, using the Mann–Whitney U test, for comparisons between the different groups are depicted as *p<0.05 versus normal controls (NC), **p<0.05 versus prerituximab (pre-RX), ***p<0.05 discordant clinically non-relapsing (D-NR) versus discordant clinically relapsing (D-R). C-R, concordant clinically relapsing.
We also used regression analysis to determine whether serum BAFF levels were related to the percentage of particular B-cell subsets. This produced mostly negative results, although higher BAFF levels were statistically associated with numbers of preswitch memory B cells in patients who were relapsing more than 3 months after B-cell repopulation (D-R; p=0.02; Mann–Whitney; data not shown).
Relationship of BAFF-R expression and B-cell phenotypes with months between repopulation and relapse
It is also possible that differences in levels of BAFF-R and B-cell phenotype distribution in patients discordant for repopulation and relapse (D-R and D-NR cohorts) were related at least partly to differences in the time of sample collection following repopulation. We therefore compared months between repopulation and relapse in the two cohorts and found them not to be significantly different (months between repopulation and sample collection in the D-NR cohort range 4–46, median=13; D-R cohort range 5–24, median=7; Mann–Whitney rank sum p=0.594).
Discussion
Using the 11C1 monoclonal antibody, which recognises occupied and unoccupied BAFF-R on human cells,25 we found that BAFF-R levels on CD27+ and CD27− B-cell subpopulations in RA patients before rituximab were similar to those of B cells from NC, although there was a tendency towards lower values especially in memory B-cell populations (MFI; p=0.098; table 1 and figure 2). We also report that BAFF-R expression was significantly reduced on B cells from patients with RA when they were undergoing relapse following rituximab. Low BAFF-R levels were not due to an increase in circulating plasmablasts (data not shown), which have been associated with clinical relapse.26 Significantly, reduced BAFF-R expression was found in both naive and memory subpopulations and on B cells from patients who relapsed when B cells returned, as well as in those patients who relapsed more than 3 months after B-cell return. In addition, as all patients in the pre-RX group had active RA and were studied before the induction of B-cell depletion therapy for resistant disease the results also imply that reduced BAFF-R expression was not necessarily a consequence of inflammation and active disease, but was more likely related to the re-establishment of disease.
The bioactive trimeric form of soluble BAFF is produced by a variety of cell types, namely myeloid and dendritic cells, neutrophils and some stromal cells.27 In humans, BAFF signalling through BAFF-R does not appear to be directly involved in B-cell maturation or in most T-cell-independent responses.6 BAFF-R is expressed on human B cells from the transitional stage onwards, and is then lost from memory B cells as they differentiate into Ig-secreting plasma cells.28 Stimulation through Toll-like receptor (CpG IL2,15) pathways is more efficient than T-dependent pathways (CD40L,IL2,10) at downregulating BAFF-R expression on memory B-cell populations.7 28 It was also found that stimulation of naive cells through either pathway had no effect on BAFF-R expression, although positive responses of naive human cells to either stimulus are poor.28
Investigations of BAFF-R expression on B cells from patients with rheumatic diseases are limited and to some extent report contrasting findings, and none describe patients with RA or the effect of rituximab. Carter et al25 showed that normal levels of BAFF-R were present on B cells in patients with SLE but were of reduced availability due to occupancy by soluble BAFF. Low levels of BAFF-R expression and a negative correlation between BAFF-R expression and higher disease activity in patients with SLE have been described.19 In patients with Sjogrens syndrome, BAFF-R expression was decreased on both naive and memory B cells, with the lowest levels found in patients with extraglandular involvement.19 Analysis of BAFF-R messenger RNA from B cells from these patients suggested that the lower expression was due to a post-translational modification and not to any reduction in BAFF-R mRNA. This could be explained if decreased BAFF-R expression reflected the recent binding of bioactive BAFF to BAFF-R followed by internalisation or shedding.25 In turn, this could be due to the chronic exposure of B cells to constantly high circulating levels of BAFF, which are often found in these patients. Carter et al25 showed that the receptor/cytokine complex was not internalised. Loss of the receptor following ligation by BAFF was, however, confirmed by Sellam et al,19 who showed that overnight, but not short-term, culture with physiologically encountered concentrations of BAFF reduced BAFF-R levels on B cells from normal donors.
Our experience here and in earlier studies17 was that BAFF levels were not significantly raised in patients with active RA. However, following rituximab, BAFF levels rise rapidly and remain raised for variable periods of time. In our serial studies,17 we found that in individual patients, BAFF levels fell more sharply in patients in whom relapse was associated with B-cell return, than in those patients who relapsed months after B-cell return. This may have reflected increased BAFF binding to BAFF-R or TACI following B-cell receptor (BCR) engagement. We could not confirm these results here as individual patients were not studied serially over the course of their cycle of treatment. In the present study, serum BAFF remained raised to similar levels in sera from patient cohorts discordant or concordant with respect to B-cell return and relapse. Within each group of patients, however, there was no correlation between BAFF levels and BAFF-R expression (figure 3) suggesting that reduced expression was not solely due to chronically raised BAFF levels, although this possibility could not be excluded with certainty.
When levels of expression of BAFF-R on naive and memory populations in pre-RX patients and controls were compared, CD27+ memory B-cell values tended to be lower, although this did not reach statistical significance (p=0.07; data not shown). The main finding of the study was that BAFF-R levels were significantly reduced on both naive and memory B cells from relapsing, compared with non-relapsing, patients and pre-RX patients. This suggested that the reduction in BAFF-R levels may have been initiated in naive cells, which then carried the ‘low’ BAFF-R phenotype into the mature memory B-cell stage. In-vitro experiments by others have shown that BAFF could modulate BAFF-R expression in a dose-dependent manner.25 As we found no correlation between raised BAFF levels and reduced BAFF-R expression and low BAFF-R expression on naive populations as well as memory B cells associated with the re-establishment of disease, we suggest that another mechanism may be responsible.
Patients with RA have been shown to possess a number of genetic polymorphisms linked to tolerance checkpoints in the bone marrow or the periphery.29 Evidence of altered thresholds for BCR signalling30 has been described in patients with RA and suggested to contribute to the generation of autoreactive cells. There is also accumulating evidence that pre-B/immature B cells from patients with RA are pre-activated before they exit from the bone marrow.31 It was recently shown that B cells at the pre-B/immature naive stage in the bone marrow of patients with RA had evidence of increased TLR9 protein expression and mRNA compared with osteoarthritis patients and also expressed the activation marker CD86.32 Disease-induced changes, such as the formation of germinal centre-like structures, cytokines such as TNFα, or increased levels of bacterial DNA/RNA in the bone marrow environment in patients with RA may disrupt the environment in which pre-B cells are developing.33 Reduced ability to express receptors for the pro-survival cytokine BAFF on transitional and naive B cells after exiting the bone marrow may result as part of the unusual cytokine environment in which the B cells developed.
Once in the circulation, BCR specificity becomes a crucial determinant of B-cell survival and function. The expression of BCR specific for citrullinated proteins and increased numbers of RhF-specific B cells have been described by Samuels et al31 within early (CD10+IgM+) populations exiting the bone marrow and also in mature naive B cells (CD10−IgM+) in RA patients compared with normal individuals. The conclusion is therefore that alterations of tolerance checkpoints both within the bone marrow and in the periphery may be occurring in patients with RA. Within the polyreactive pool exiting the bone marrow, BCR specificities with promiscuous potential, such as those possessed by the relatively high proportion of RhF B cells that can obtain help from T cells with specificity for a wide range of processed peptides,34 may have a survival advantage that could override the disadvantages of low BAFF-R expression/signalling in a genetically predisposed individual. We have shown that BAFF levels do not appear to be limiting in relapsing patients, but lower levels of BAFF-R expression on newly generated B cells may reduce survival potential and favour ‘expanded’ cohorts of B cells able more easily to obtain productive, but not necessarily antigen-specific, T-cell help.
As well as being an effective treatment for many patients with RA, rituximab gives us an opportunity to study elements important for relapse through our ability to study the re-establishment of disease. Following clearance of rituximab from the bone marrow, newly generated B cells expand into an environment that has retained a variable number of resistant B cells, a mature T-cell compartment and long-lived plasma cells. First, we know that B-cell return to the periphery is mandatory for relapse.20 Second, the gap between B-cell return and relapse may reflect the relationship between the re-expansion of surviving memory B-cell clones under the influence of newly generated B cells. In both scenarios, maturation into Ig-secreting cells is often coincident with relapse, as shown by rises in autoantibodies,17 35 and also by the reported association with circulating plasmablasts.23 Our finding of a lower proportion of postswitched memory B cells in non-relapsing, repopulated patients compared with relapsing patients supports the view that B-cell class-switch recombination is associated with relapse. The depletion of CD27hi memory B cells has previously been associated with clinical response to rituximab23 36 and low circulating numbers of IgD−CD27+ MB cells associated with low clinical activity indices and a longer clinical response, as reported by our group and others.20 37 The parallel finding that BAFF-R levels in the non-relapsing repopulated patients were similar to those of pre-RX patients support our hypothesis that the signals required for BAFF-R downregulation are intimately linked to the re-establishment of disease. We also found that a population of double-negative (IgD−CD27−) B cells was significantly raised in patients with RA compared with controls, but their proportions did not change following rituximab. These cells may be similar to a previously described memory B-cell population in patients with SLE by Wei et al,38 although others have found that only approximately 50% of this double-negative B-cell population bore markers of mutation and class switch.39
Although no statistically significant reduction in BAFF-R expression was present in RA patients before treatment, the ranges of percentage expression and MFI were wide (table 1, figure 2). It is possible that a proportion of B cells with low BAFF-R levels is being generated throughout the clinical course of RA, but that the numbers of detectable BAFF-Rlo B cells may vary.
B-cell survival and differentiation is the result of a complex interplay between survival factors, in particular the BAFF/APRIL system in concert with the specificity and strength of signal through the BCR.40 From this study, we have shown that low levels of BAFF-R expression are related to the re-expansion of possibly pathogenic species of B cells associated with relapse. The re-establishment of active disease in patients with RA after rituximab may therefore involve preferential survival properties of autoreactive B cells, due to the unusual properties of their BCR and the ability also to overcome possible consequences of lower levels of BAFF-R signalling.27 31
References
Footnotes
-
Funding The authors wish to thank the Worshipful Company of Grocers of the City of London for their generous support. RAM was supported by a grant from Fundação para a Ciência ea Tecnologia SFRH/BD/30247/2006 and IDLTO by the Alonso Martin Escudero Foundation.
-
Competing interests None.
-
Ethics approval This study was conducted with the approval of the University College Hospitals London Ethics Committee.
-
Patient consent Obtained.
-
Provenance and peer review Not commissioned; externally peer reviewed.
Linked Articles
- Miscellaneous