
Abstract
Interleukin 1β (IL-1β) is critical for the in vivo survival, expansion and effector function of IL-17–producing helper T (TH17) cells during autoimmune responses, including experimental autoimmune encephalomyelitis (EAE). However, the spatiotemporal role and cellular source of IL-1β during EAE pathogenesis are poorly defined. In the present study, we uncovered a T cell–intrinsic inflammasome that drives IL-1β production during TH17-mediated EAE pathogenesis. Activation of T cell antigen receptors induced expression of pro-IL-1β, whereas ATP stimulation triggered T cell production of IL-1β via ASC-NLRP3–dependent caspase-8 activation. IL-1R was detected on TH17 cells but not on type 1 helper T (TH1) cells, and ATP-treated TH17 cells showed enhanced survival compared with ATP-treated TH1 cells, suggesting autocrine action of TH17-derived IL-1β. Together these data reveal a critical role for IL-1β produced by a TH17 cell–intrinsic ASC–NLRP3–caspase-8 inflammasome during inflammation of the central nervous system.
This is a preview of subscription content, access via your institution
Access options
Subscribe to this journal
Receive 12 print issues and online access
$209.00 per year
only $17.42 per issue
Buy this article
- Purchase on Springer Link
- Instant access to full article PDF
Prices may be subject to local taxes which are calculated during checkout







Similar content being viewed by others

References
Frohman, E.M. Multiple sclerosis. Med. Clin. North Am. 87, 867–897 (2003).
Ransohoff, R.M. Animal models of multiple sclerosis: the good, the bad and the bottom line. Nat. Neurosci. 15, 1074–1077 (2012).
Stromnes, I.M., Cerretti, L.M., Liggitt, D., Harris, R.A. & Goverman, J.M. Differential regulation of central nervous system autoimmunity by TH1 and TH17 cells. Nat. Med. 14, 337–342 (2008).
Kroenke, M.A., Carlson, T.J., Andjelkovic, A.V. & Segal, B.M. IL-12- and IL-23-modulated T cells induce distinct types of EAE based on histology, CNS chemokine profile, and response to cytokine inhibition. J. Exp. Med. 205, 1535–1541 (2008).
Lees, J.R., Golumbek, P.T., Sim, J., Dorsey, D. & Russell, J.H. Regional CNS responses to IFN-gamma determine lesion localization patterns during EAE pathogenesis. J. Exp. Med. 205, 2633–2642 (2008).
Jäger, A., Dardalhon, V., Sobel, R.A., Bettelli, E. & Kuchroo, V.K. Th1, Th17, and Th9 effector cells induce experimental autoimmune encephalomyelitis with different pathological phenotypes. J. Immunol. 183, 7169–7177 (2009).
Cua, D.J. et al. Interleukin-23 rather than interleukin-12 is the critical cytokine for autoimmune inflammation of the brain. Nature 421, 744–748 (2003).
Veldhoen, M., Hocking, R.J., Atkins, C.J., Locksley, R.M. & Stockinger, B. TGFβ in the context of an inflammatory cytokine milieu supports de novo differentiation of IL-17-producing T cells. Immunity 24, 179–189 (2006).
Mangan, P.R. et al. Transforming growth factor-β induces development of the TH17 lineage. Nature 441, 231–234 (2006).
Chung, Y. et al. Critical regulation of early Th17 cell differentiation by interleukin-1 signaling. Immunity 30, 576–587 (2009).
Gulen, M.F. et al. The receptor SIGIRR suppresses Th17 cell proliferation via inhibition of the interleukin-1 receptor pathway and mTOR kinase activation. Immunity 32, 54–66 (2010).
Staschke, K.A. et al. IRAK4 kinase activity is required for Th17 differentiation and Th17-mediated disease. J. Immunol. 183, 568–577 (2009).
Matsuki, T., Nakae, S., Sudo, K., Horai, R. & Iwakura, Y. Abnormal T cell activation caused by the imbalance of the IL-1/IL-1R antagonist system is responsible for the development of experimental autoimmune encephalomyelitis. Int. Immunol. 18, 399–407 (2006).
Gulen, M.F. et al. Inactivation of the enzyme GSK3α by the kinase IKKi promotes AKT-mTOR signaling pathway that mediates interleukin-1-induced Th17 cell maintenance. Immunity 37, 800–812 (2012).
Chang, J. et al. MyD88 is essential to sustain mTOR activation necessary to promote T helper 17 cell proliferation by linking IL-1 and IL-23 signaling. Proc. Natl. Acad. Sci. USA 110, 2270–2275 (2013).
Gris, D. et al. NLRP3 plays a critical role in the development of experimental autoimmune encephalomyelitis by mediating Th1 and Th17 responses. J. Immunol. 185, 974–981 (2010).
Inoue, M., Williams, K.L., Gunn, M.D. & Shinohara, M.L. NLRP3 inflammasome induces chemotactic immune cell migration to the CNS in experimental autoimmune encephalomyelitis. Proc. Natl. Acad. Sci. USA 109, 10480–10485 (2012).
Inoue, M. et al. Interferon-β therapy against EAE is effective only when development of the disease depends on the NLRP3 inflammasome. Sci. Signal. 5, ra38 (2012).
Hirota, K. et al. Fate mapping of IL-17-producing T cells in inflammatory responses. Nat. Immunol. 12, 255–263 (2011).
Arbeloa, J., Perez-Samartin, A., Gottlieb, M. & Matute, C. P2X7 receptor blockade prevents ATP excitotoxicity in neurons and reduces brain damage after ischemia. Neurobiol. Dis. 45, 954–961 (2012).
Qu, Y. et al. Pannexin-1 is required for ATP release during apoptosis but not for inflammasome activation. J. Immunol. 186, 6553–6561 (2011).
Gordon, G.R. et al. Norepinephrine triggers release of glial ATP to increase postsynaptic efficacy. Nat. Neurosci. 8, 1078–1086 (2005).
Antonopoulos, C., El Sanadi, C., Kaiser, W.J., Mocarski, E.S. & Dubyak, G.R. Proapoptotic chemotherapeutic drugs induce noncanonical processing and release of IL-1β via caspase-8 in dendritic cells. J. Immunol. 191, 4789–4803 (2013).
Gringhuis, S.I. et al. Dectin-1 is an extracellular pathogen sensor for the induction and processing of IL-1β via a noncanonical caspase-8 inflammasome. Nat. Immunol. 13, 246–254 (2012).
Bossaller, L. et al. FAS (CD95) mediates noncanonical IL-1β and IL-18 maturation via caspase-8 in an RIP3-independent manner. J. Immunol. 189, 5508–5512 (2012).
Oberst, A. et al. Catalytic activity of the caspase-8-FLIP(L) complex inhibits RIPK3-dependent necrosis. Nature 471, 363–367 (2011).
Leverrier, S., Salvesen, G.S. & Walsh, C.M. Enzymatically active single chain caspase-8 maintains T-cell survival during clonal expansion. Cell Death Differ. 18, 90–98 (2011).
Bell, B.D. et al. FADD and caspase-8 control the outcome of autophagic signaling in proliferating T cells. Proc. Natl. Acad. Sci. USA 105, 16677–16682 (2008).
Kang, T.B. et al. Caspase-8 serves both apoptotic and nonapoptotic roles. J. Immunol. 173, 2976–2984 (2004).
Salmena, L. et al. Essential role for caspase 8 in T-cell homeostasis and T-cell-mediated immunity. Genes Dev. 17, 883–895 (2003).
Chun, H.J. et al. Pleiotropic defects in lymphocyte activation caused by caspase-8 mutations lead to human immunodeficiency. Nature 419, 395–399 (2002).
Bauernfeind, F.G. et al. NF-κB activating pattern recognition and cytokine receptors license NLRP3 inflammasome activation by regulating NLRP3 expression. J. Immunol. 183, 787–791 (2009).
Martin, B.N. et al. IKKα negatively regulates ASC-dependent inflammasome activation. Nat. Commun. 5, 4977 (2014).
Bryan, N.B., Dorfleutner, A., Rojanasakul, Y. & Stehlik, C. Activation of inflammasomes requires intracellular redistribution of the apoptotic speck-like protein containing a caspase recruitment domain. J. Immunol. 182, 3173–3182 (2009).
Schenten, D. et al. Signaling through the adaptor molecule MyD88 in CD4+ T cells is required to overcome suppression by regulatory T cells. Immunity 40, 78–90 (2014).
Doitsh, G. et al. Cell death by pyroptosis drives CD4 T-cell depletion in HIV-1 infection. Nature 505, 509–514 (2014).
Shaw, P.J. et al. Critical role for PYCARD/ASC in the development of experimental autoimmune encephalomyelitis. J. Immunol. 184, 4610–4614 (2010).
El-Behi, M. et al. The encephalitogenicity of TH17 cells is dependent on IL-1– and IL-23–induced production of the cytokine GM-CSF. Nat. Immunol. 12, 568–575 (2011).
Wei, D.G. et al. Expansion and long-range differentiation of the NKT cell lineage in mice expressing CD1d exclusively on cortical thymocytes. J. Exp. Med. 202, 239–248 (2005).
Shornick, L.P. et al. Mice deficient in IL-1β manifest impaired contact hypersensitivity to trinitrochlorobenzone. J. Exp. Med. 183, 1427–1436 (1996).
Tomalka, J. et al. A novel role for the NLRC4 inflammasome in mucosal defenses against the fungal pathogen Candida albicans. PLoS Pathog. 7, e1002379 (2011).
Sarkar, A. et al. Caspase-1 regulates Escherichia coli sepsis and splenic B cell apoptosis independent of interleukin-1β and interleukin-18. Am. J. Respir. Crit. Care Med. 174, 1003–1010 (2006).
Kuida, K. et al. Altered cytokine export and apoptosis in mice deficient in interleukin-1β converting enzyme. Science 267, 2000–2003 (1995).
Kaiser, W.J. et al. RIP3 mediates the embryonic lethality of caspase-8-deficient mice. Nature 471, 368–372 (2011).
Qian, Y. et al. The adaptor Act1 is required for interleukin 17–dependent signaling associated with autoimmune and inflammatory disease. Nat. Immunol. 8, 247–256 (2007).
Kang, Z. et al. Astrocyte-restricted ablation of interleukin-17-induced Act1-mediated signaling ameliorates autoimmune encephalomyelitis. Immunity 32, 414–425 (2010).
Suzuki, S. et al. Shigella type III secretion protein MxiI is recognized by Naip2 to induce Nlrc4 inflammasome activation independently of Pkcδ. PLoS Pathog. 10, e1003926 (2014).
Acknowledgements
This investigation was supported by the National Multiple Sclerosis Society (grant RG5130A2/1 to X.L.) and the US National Institutes of Health (grants 5R01NS071996-05, 1RO1AA023722 and MSTP-T32GM007250 to X.L.).
Author information
Authors and Affiliations
Contributions
B.N.M. and C.W. did the experiments and analyzed the data; C.Z., Z.K., M.F.G., J.A.Z., J.Z., C.E.-S. and G.B. contributed to the experiments; J.D. and P.G.P. helped to make the Cd3e−/− and Il1b−/− bone marrow chimera mice; A.A., Y.I., A.S., M.D.W., W.J.K., E.S.M., M.E.R., P.L.F. and A.G.H. provided reagents and participated in discussion; B.M. and G.R.D. contributed reagents, helped design experiments and helped edit the manuscript; B.N.M., C.W. and X.L. wrote the manuscript; R.M.R. and X.L. conceived the study, oversaw the experiments and analyzed the data.
Corresponding authors
Ethics declarations
Competing interests
The authors declare no competing financial interests.
Integrated supplementary information
Supplementary Figure 1 Generation of the Pycard conditional knockout allele.
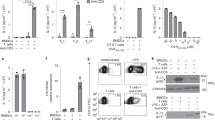
(a) The Pycard gene is localized on chromosome 7 and contains three coding exons. For generation of the conditional knockout allele, the entire open reading frame including all three exons was targeted for conditional deletion. After generation of the targeting vector (using Pcr4.0 backbone), C57BL/6 embryonic stem cells were injected for gene targeting. Southern hybridization was used to identify progeny bearing the recombinant allele. (b) CD4+ T Cells were sorted from Ascf/+Lck-Cre and Ascf/-Lck-Cre mice by flow cytometry, and mRNA expression of three different exons of ASC was measured by Real-time PCR analysis. Unpaired t test was used to analyze the data. Data are representative of three independent experiments (b). n=3 mice per group in each experiment. Error bars represent s.e.m. *P < 0.05.
Supplementary Figure 2 Numbers of T cells and B cells were similar in the lymph nodes, spleens and thymus between Ascf/+Lck-Cre and Ascf/–Lck-Cre mice.
(a) The total cell number of CD4+, CD8+ and B220+CD19+ cells were counted from lymph nodes and spleen of Ascf/+Lck-Cre and Ascf/-Lck-Cre mice. (b) Total cell number of CD4-CD8-, CD4+CD8-, CD4+ and CD8+ were counted from thymus of Ascf/+Lck-Cre and Ascf/-Lck-Cre mice. Data are representative of two independent experiments (a-b). n=5 mice per group in each experiment. Error bars represent s.e.m.
Supplementary Figure 3 The EAE phenotype of adoptive transfer of sorted CD4+ T cells from Ascf/+Lck-Cre and Ascf/–Lck-Cre mice.
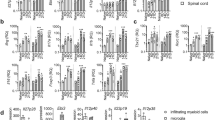
(a) Absolute numbers of CD4+ IL-17A+ cells (left panel) and concentrations of IL-17A in the supernatant of cells (right panel) from the draining lymph nodes and spleens of Ascf/+Lck-Cre and Ascf/-Lck-Cre mice immunized with MOG35-55 and harvested 10 days after immunization, followed by ex vivo re-stimulation in the presence of MOG35-55 and IL-23. (b-d) Cells from lymph nodes of Ascf/+Lck-Cre and Ascf/-Lck-Cre mice 10 days after immunization with MOG35-55 were re-stimulated with MOG 35-55 in vitro in the presence of recombinant IL-23 for 5 days, sorted for CD4+ T cells by flow cytometry, and then transferred into CD45.1 congenic mice. (b) Graph represents the average clinical score after T cell transfer. (c) Absolute numbers of CNS-infiltrating CD45.2+ CD4+ cells were determined at the peak of disease. Brains and spinal cords were harvested together and CD45.2+CD4+ cells were stained with anti-CD45.2 and anti-CD4 antibodies, followed by flow cytometric analysis. (d) Absolute numbers of CNS-infiltrating cells were determined at the peak of disease. Brains and spinal cords were harvested together and mononuclear infiltrating cells were stained with anti-CD45, anti-CD4, anti-CD8, anti-F4/80, anti-Ly6C, anti-CD19 and anti-Ly6G antibodies, followed by flow cytometric analysis. Two-way ANOVA (b) and Unpaired t test (a, c and d) were used to analyzed the data. Data are representative of three independent experiments (a-d). n=5 mice per group in each experiment. Error bars represent s.e.m. *P < 0.05 (a, c and d). Error bars represent s.d. *P < 0.05 (b).
Supplementary Figure 4 Comparison of T cell and macrophage secretion of IL-1β.
Macrophages and TH17 cells were left untreated or treated with LPS for 4 hours (1ug/ul for macrophages) or LPS+ATP (5 mM, 30 mins) or ATP alone (5 mM, 8 hours). Cell pellet and supernatant were collected and subjected to western blotting for IL-1β and Actin. Data are representative of two independent experiments.
Supplementary Figure 6 The EAE phenotype of Cd3e−/−Il1b−/− chimera mice.
(a) Lethal irradiated WT mice were reconstituted with WT+Cd3e-/- bone marrow or Il1b-/-+Cd3e-/- bone marrow. 6 weeks after reconstitution, mice were immunized with MOG35-55 peptide and 200 ng pertussis toxin on days 1 and 4. Graph represents the average clinical score after active immunization. (b) Inflammatory gene expression in the lumbar spinal cord was assessed at the peak of disease. (c) Absolute numbers of CNS-infiltrating cells were determined at the peak of disease. Brains and spinal cords were harvested together and mononuclear infiltrating cells were stained with anti-CD45, anti-CD4, anti-CD8, anti-F4/80, anti-Ly6C and anti-Ly6G antibodies, followed by flow cytometric analysis. Two-way ANOVA (a) and Unpaired t test (b and c) were used to analyzed the data. Data are representative of three independent experiments (a-c). n=5 mice per group in each experiment. Error bars represent s.e.m. *P < 0.05.
Supplementary information
Supplementary Text and Figures
Supplementary Figures 1–6 and Supplementary Table 1 (PDF 1734 kb)
Rights and permissions
About this article

Cite this article
Martin, B., Wang, C., Zhang, Cj. et al. T cell–intrinsic ASC critically promotes TH17-mediated experimental autoimmune encephalomyelitis. Nat Immunol 17, 583–592 (2016). https://doi.org/10.1038/ni.3389
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1038/ni.3389
This article is cited by
-
IL-1α-releasing TH17 cells live long and prosper
Nature Immunology (2023)
-
Inflammasome activation occurs in CD4+ and CD8+ T cells during graft-versus-host disease
Cell Death & Disease (2023)
-
The interplay between T lymphocytes and macrophages in myocardial ischemia/reperfusion injury
Molecular and Cellular Biochemistry (2023)
-
Human TH17 cells engage gasdermin E pores to release IL-1α on NLRP3 inflammasome activation
Nature Immunology (2023)
-
TH17 cells promote CNS inflammation by sensing danger signals via Mincle
Nature Communications (2022)
