
Abstract
Objective. To identify differentially expressed microRNA (miRNA) in muscle biopsies (MBx) from 15 untreated children with juvenile dermatomyositis (JDM) compared with 5 controls.
Methods. Following MBx miRNA profiling, differentially expressed miRNA and their protein targets were validated by quantitative real-time PCR (qRT-PCR) and immunological assay. The association of miRNA-10a and miRNA-10b with clinical data was evaluated, including Disease Activity Score (DAS), von Willebrand factor antigen (vWF:Ag), nailfold capillary end row loops, duration of untreated disease, and tumor necrosis factor (TNF)-α-308A allele.
Results. In JDM, 16/362 miRNA were significantly differentially expressed [false discovery rate (FDR) < 0.05]. Among these, miRNA-10a was the most downregulated miRNA in both FDR and ranking of fold change: miRNA-10a = −2.27-fold, miRNA-10b = −1.80-fold. Decreased miRNA-10a and miRNA-10b expressions were confirmed using qRT-PCR: −4.16 and −2.59 fold, respectively. The qRT-PCR documented that decreased miRNA-10a expression was related to increased vascular cell adhesion molecule 1 in 13 of these JDM cases (correlation −0.67, p = 0.012), unlike miRNA-10b data (not significant). Concurrent JDM plasma contained increased levels of interleukin (IL) 6 (p = 0.0363), IL-8 (p = 0.0005), TNF-α (p = 0.0011), and monocyte chemoattractant proteins 1 (p = 0.0139). Decreased miRNA-10a, but not miRNA-10b, was associated with the TNF-α-308A allele (p = 0.015). In the 15 JDM, a trend of association of miRNA-10a (but not miRNA-10b) with vWF:Ag and DAS was observed.
Conclusion. MiRNA-10a downregulation is an important element in untreated JDM muscle pathophysiology. We speculate that muscle miRNA expression in adult dermatomyositis differs from muscle miRNA expression in untreated childhood JDM.
- JUVENILE DERMATOMYOSITIS
- MicroRNA MICROARRAY
- MicroRNA-10
- INFLAMMATORY CYTOKINES
Although juvenile dermatomyositis (JDM) is a rare disease, it is the most common pediatric inflammatory myopathy with an incidence rate in the United States of about 3.2 children per million per year1,2. This systemic vasculopathy is characterized by a classic heliotrope rash, Gottron papules, symmetrical proximal muscle weakness, elevated serum levels of muscle-derived enzymes, and evidence of a myopathic process on electromyogram3,4. The “gold standard” of diagnosis is the muscle biopsy (MBx) in which perifascicular muscle fiber atrophy is associated with progressive capillary occlusion and an inflammatory infiltrate composed primarily of mononuclear cells5,6. The children manifest muscle and skin involvement, which are evaluated reliably by the Disease Activity Scores (DAS)7,8. Prolonged inflammation results in loss of nailfold capillary end row loops (ERL)9 and the development of dystrophic calcifications in 10% to 35% of children10. Several inflammatory and cell death pathways involving nuclear factor-κB (NF-κB)11,12 and apoptosis13 are documented in juvenile myositis tissue damage, and Type 1 interferons have been implicated as major factors contributing to the inflammatory process14,15. Increased levels of tumor necrosis factor (TNF)-α16 and Type 1 interferon activity17 have been confirmed in JDM sera. Older patients who had JDM in childhood are at risk for considerable damage to their cardiovascular system, as are adults with dermatomyositis (DM)18,19. However, an investigation of the involvement of microRNA (miRNA) in the pathophysiology of JDM muscle involvement has not been previously reported, to our knowledge.
MiRNA are short, noncoding RNA that inhibit miRNA by binding to complementary sequences on target miRNA, resulting in translational repression or degradation and gene silencing20, and have a complex role in striated muscle21. One miRNA can regulate multiple target miRNA21 and altered miRNA expression has been identified in a wide spectrum of human illnesses, including autoimmune diseases22, cancer23, and cardiovascular diseases24. Several miRNA (e.g., miRNA-155, -146a, -146b, -203, -214, -1, -133, and -206) have been implicated in primary muscular disorders in adults, including DM and polymyositis (PM)25,26. The purpose of our study was to assess miRNA expression in muscle from untreated children with active symptoms of definite/probable JDM compared with healthy orthopedic controls, and to determine the association of these regulatory factors with the children’s clinical status.
MATERIALS AND METHODS
Patient population
Under magnetic resonance imaging guidance, a diagnostic MBx was performed of the involved area in untreated children with definite/probable JDM (Bohan and Peter criteria)3,4. Prior to biopsy, age-appropriate JDM-informed consent (IRB# 2008-13457) was obtained and at surgery, small samples of muscle and plasma were collected for research. The patient population consisted of 9 girls and 6 boys for the miRNA profiling and validation, with a mean age of 6.2 ± 3.1 years and the following racial distribution: white 80%, non-white 20%. Five healthy orthopedic controls (2 girls, 3 boys) who were white, mean age 12.3 ± 4.2 years, donated both muscle and plasma (IRB# 2001-11715). In this control group, 2 had scoliosis, 1 had an anterior cruciate ligament reconstruction, 1 underwent an osteochondroma excision, and the last control had an adductor myotomy.
DAS
The DAS of both skin involvement (DAS SK) and muscle weakness (DAS WK) were assessed by 1 physician (LMP) at the time that the plasma and MBx were obtained. Scores ranged from 0–9 for DAS SK and 0–11 for DAS WK; scores greater than 0 were considered abnormal7.
Duration of untreated disease (DUD)
Eight children with JDM had disease duration of < 3 months with a mean of 1.69 ± 0.76 months, while 7 children had JDM disease duration ≥ 3 months with a mean of 11.82 ± 10.08 months; they were classified, respectively, as having untreated disease of short and long durations.
Determination of TNF-α alleles at −308
The primers and probe used for detection of alleles were synthesized at the Northwestern University Biotechnical Facility (Chicago, Illinois, USA). Anticoagulated whole blood was collected in EDTA, and then aliquoted and stored at −70°C until DNA isolation was performed. DNA was isolated with the use of a Qiagen DNA isolation kit (Valencia). TNF-α alleles were identified with quantitative real-time PCR (qRT-PCR), as reported16.
Measurement of the ERL number
All children underwent nailfold capillaroscopy (NFC) at the time of diagnosis, consisting of freeze-frame digital microscopy images of each of the 8 fingers, excluding the thumbs. NFC images were obtained using a Leica Z6APO Stereomicroscope, on which was mounted a Leica DFC295 Digital Microscope Camera. The images were analyzed as described. A mean number of ERL of < 7.00/mm was considered abnormal, as reported9.
von Willebrand factor antigen (vWF:Ag) laboratory testing
Standard determination of the vWF:Ag was obtained at diagnosis when children with JDM visited the Ann and Robert H. Lurie Children’s Hospital of Chicago. The vWF:Ag assay is a quantitative turbidimetric assay (Siemens).
MiRNA expression profiling
Total RNA was extracted using a miRCURY RNA Isolation Kit (Exiqon), and RNA samples from both normal controls and JDM were sent to Exiqon for miRNA expression profiling. RNA quality control was performed using a 2100 Bioanalyzer (Agilent). The 2100 Bioanalyzer Expert software was used to generate an RNA Integrity Number that provides a numerical assessment of the integrity of RNA. Following quality control, RNA samples were labeled using a miRCURY LNA microRNA Power Labeling Kit according to Exiqon’s protocol. Array experiments were conducted as double-channel Hy3/Hy5 experiments in triplicates on Exiqon’s miRCURY LNA microRNA Array (6th Gen). Hybridization of labeled RNA to the array was performed on Tecan HS4800 proautomated hybridization stations and scanned with Agilent G2505B Microarray Scanners. The obtained microarray images were analyzed using Genepix Pro 4.0 software.
Measurements of qRT-PCR
Total RNA was extracted from the muscle samples using a miRCURY RNA Isolation Kit (Exiqon) according to the manufacturer’s instructions. For validation of differentially expressed miRNA, the ABI 7500 thermocycler and TaqMan miRNA-10a, miRNA-10b, IL-6, and vascular cell adhesion molecule 1 (VCAM-1) primer sets from Life Technology were used for qRT-PCR, as previously reported27. Relative miRNA expression levels were calculated according to the 2(−ΔΔC(T)) method28; U6 was used for miRNA-10a and miRNA-10b normalization, and β-actin was used for IL-6 and VCAM-1.
Measurement of plasma levels of IL-6, IL-8, TNF-α, and monocyte chemoattractant proteins 1 (MCP-1)
Plasma samples from the same 15 untreated JDM children and 5 pediatric controls were assayed. A Human MCP-1 Ultra-Sensitive Kit (K151AYC-1) and a Human ProInflammatory-4 II Ultra-Sensitive Kit (K15009C-1) from MesoScale Discovery assessed the levels of plasma MCP-1 and of IL-6, IL-8, and TNF-α, respectively, following the manufacturer’s protocol. The data were analyzed using the MesoScale Discovery Workbench software.
Statistical analysis
The RNA from muscle samples were hybridized and analyzed against a common reference pool of all 20 samples. For these samples, the quantified signals (background corrected) were normalized using the global locally weighted scatterplot smoothing regression algorithm. The background threshold was calculated for each microarray slide as 1.2 times the 25% signal intensity. MiRNA with intensities above threshold in < 20% of the samples were removed from the final dataset used for expression analysis. This filtering procedure discarded 1077 miRNA (out of a total of 1439), leaving 362 miRNA for data analysis. A 2-sample Student t test was used to compare the expression levels between JDM and controls for each miRNA. The false discovery rate (FDR) was calculated using the Bioconductor package, q value. The 2-sample Student t test was used for the following: (1) validation of qRT-PCR for differential expression of miRNA-10a and miRNA-10b, and for IL-6 and VCAM-1 between JDM and control; (2) miRNA and protein–coding gene expression levels in muscle; (3) the difference in the level of inflammatory cytokines, MCP-1, TNF-α, IL-6, and IL-8, in the plasma from children with JDM and pediatric control group; and (4) associations of miRNA-10a and miRNA-10b expression levels with the TNF-α-308A allele (GA/AA). Correlations of the miRNA-10a and miRNA-10b expression levels with inflammatory marker VCAM-1 and other clinical and demographic variables, including DAS SK, DAS WK, DAS Total, ERL, DUD, and vWF:Ag levels, were calculated using the Pearson correlation coefficient (r).
RESULTS
Differentially expressed miRNA in muscle from untreated children with JDM
To perform microRNA expression profiling, we tested 15 JDM and 5 healthy pediatric controls. Information concerning the test populations is presented in Supplemental Table 1 (available online at jrheum.org). MiRNA microarray analysis revealed 362 miRNA expressed in both JDM and the pediatric controls. The expression analysis identified 16 miRNA that were significantly differentially expressed (FDR < 0.05) in JDM compared with healthy pediatric control. Table 1 presents the 12 downregulated miRNA and the 4 upregulated miRNA. Among these 12 miRNA, both miRNA-10a and miRNA-10b displayed the largest fold change (miRNA-10a fold change = −2.27, and miRNA-10b fold change = −1.80) as well as an appropriate q value (Table 1), which was confirmed by qRT-PCR. The qRT-PCR results (Figure 1) documented that miRNA-10a and miRNA-10b levels were decreased by 4.16-fold (p = 0.008) and 2.59-fold (p = 0.001), respectively. These miRNA qRT-PCR expression levels were consistent with the results from the miRNA profiling of the same MBx tissue.

Validation of differentially expressed miRNA-10a and miRNA-10b by qRT-PCR. The qRT-PCR was performed with TaqMan primers on MBx from children with untreated JDM and healthy pediatric controls to validate the differential expression of miRNA-10a and miRNA-10b. Values are mean ± SD. Student t test was used for statistical analysis. p < 0.05 was considered significant. * p = 0.0004. ** p = 0.00002. qRT-PCR: quantitative real-time PCR; MBx: muscle biopsy; JDM: juvenile dermatomyositis.
Exiqon array for microRNA (miRNA) expression: comparison of muscle from 15 children with untreated JDM with muscle from 5 healthy pediatric controls.
MiRNA-10 target genes in JDM MBx
Once miRNA-10a and miRNA-10b were confirmed as downregulated miRNA in diagnostic MBx from patients with untreated JDM, we sought to further define their target genes. A target gene prediction search using Targetscan was performed. This search identified 178 conserved target genes. Comparison of those 178 target genes with our previous gene expression data from MBx in JDM29 disclosed that 32 among those 178 miRNA-10 target genes were significantly altered more than 1.5-fold (p < 0.05; Supplemental Table 2, available online at jrheum.org). This listing of the 32 target genes included mitogen-activated protein kinase kinase kinase 7 (MAP3K7) and ß transducin repeat containing E3 ubiquitin protein ligase (β-TRC), which have been experimentally confirmed as miRNA-10 targets and regulate the induction of proinflammatory cytokines through NF-κB pathway30. The next set of experiments was focused on specific proinflammatory biomarkers regulated by the NF-κB pathway in MBx and plasma from these same children with untreated JDM.
Decreased miRNA-10a is associated with increased IL-6 and VCAM-1 gene expression in the same MBx tissue
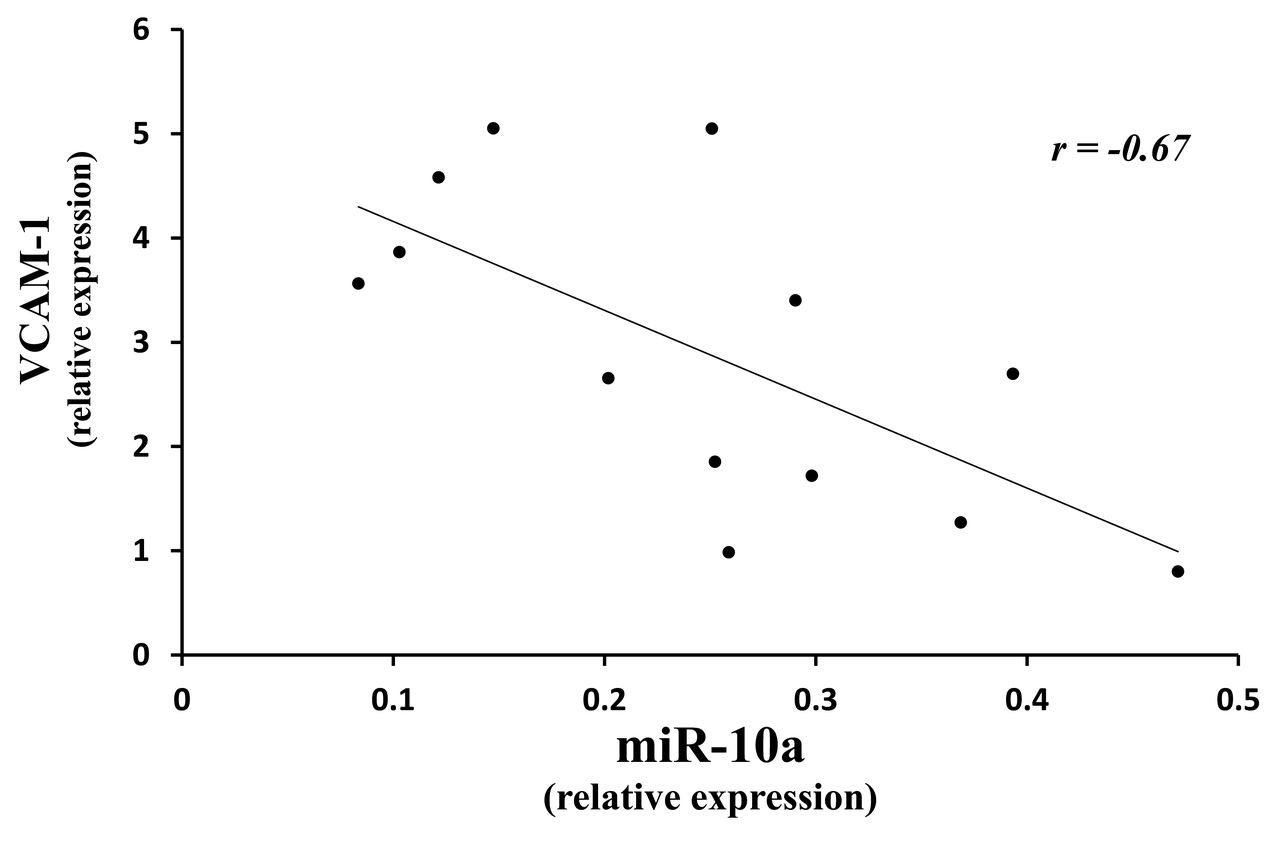
IL-6 and VCAM-1 are induced downstream proinflammatory mediators when the NF-κB pathway is activated. Using qRT-PCR testing for IL-6 and VCAM-1 in the same set of untreated muscle tissue, we found increased expression: IL-6 = 2.8 ± 2.2-fold (p < 0.015), and VCAM-1 = 2.9 ± 1.5-fold (p < 0.014), respectively (Figure 2). Further, the diminished miRNA-10a was inversely related to an increase in VCAM-1 in 13 of the same JDM (no muscle remained from the other 2 children) compared with 5 controls (correlation was −0.67; Figure 3); decreased miRNA-10b levels were not associated with the increased values for VCAM-1 (correlation was −0.38, p = 0.19). These data suggest that miRNA-10a is a more effective facilitator of inflammatory damage than miRNA-10b.

The qRT-PCR quantification for IL-6 and VCAM-1 in muscle of untreated JDM and controls. The qRT-PCR was performed with TaqMan primers on MBx from children with untreated JDM and healthy pediatric controls to measure IL-6 and VCAM-1 gene expression levels. Values are mean ± SD. Student t test was used for statistical analysis. p < 0.05 was considered significant. * p = 0.0151. ** p = 0.0143. qRT-PCR: quantitative real-time PCR; IL-6: interleukin 6; VCAM-1: vascular cell adhesion molecule 1; JDM: juvenile dermatomyositis; MBx: muscle biopsy.

{kind=link}
{kind=link}
{kind=link}
Correlation of decreased miRNA-10a with increased VCAM-1 gene expression by qRT-PCR in muscle from children with untreated JDM. Correlation of the miRNA-10a expression levels with VCAM-1 gene expression was calculated using the Pearson correlation coefficient (r) and r −0.670 (p = 0.012). MiRNA: microRNA; VCAM-1: vascular cell adhesion molecule 1; qRT-PCR: quantitative real-time PCR; JDM: juvenile dermatomyositis.
Increased levels of IL-6, IL-8, TNF-α, and MCP-1 in JDM plasma
Measurement of these proinflammatory mediators showed that they were significantly elevated in JDM plasma compared with healthy orthopedic controls (Table 2).
Elevated levels of IL-6, IL-8, TNF-α, and MCP-1 proteins (pg/ml) in plasma from children with untreated JDM. IL-6, IL-8, TNF-α, and MCP-1 levels in plasma from children with untreated JDM (n = 15) and healthy pediatric controls (n = 5) were assayed using a human ProInflammatory 4-II Ultra-Sensitive Kit and a human MCP-1 Ultra-Sensitive Kit from MesoScale Discovery. Student t test was used for statistical analysis. P < 0.05 was considered significant. Values are mean ± SD.
Association of decreased miRNA-10a, but not miRNA-10b, expression levels with clinical and laboratory features of JDM
Expression levels of miRNA-10a were significantly different between patients with JDM with the G/G allele compared with those with the G/A allele; miRNA-10a relative expression was reduced from 1.00 ± 0.68 in normal pediatric controls to 0.30 ± 0.12 in children with JDM with the G/G allele and further reduced to 0.13 ± 0.03 in children with JDM with the G/A allele (p = 0.015; Table 3). There was a trend of association of miRNA-10a with vWF:Ag data, even though in this untreated patient group, vWF:Ag values were within the normal range for the individual’s blood group antigen for all but 1 child: (r −0.5, p = 0.06) and DAS total (r −0.45, p = 0.09), but not ERL (r −0.15, p = 0.57) or DUD (r 0.23, p = 0.4; Table 3).
Fifteen children with untreated JDM and 5 healthy controls: clinical and laboratory associations. Diminished expression of miRNA-10a was significantly associated with the presence of the A allele at TNF-α-308 promoter region (p = 0.015). Values are mean ± SD unless otherwise specified (n = 15).
DISCUSSION
Relationship of miRNA and DM
Emerging data indicate that specific miRNA may play an important regulatory role in the inflammatory myopathies25,26 and in muscle physiology in general21. Similar miRNA may be present in more than 1 tissue in the same disease. For example, miRNA-223 was decreased in cutaneous Gottron papules of Japanese adults with untreated DM and clinically amyopathic DM, but not the skin of patients with psoriasis31. Another study showed miRNA-223 was also dysregulated in muscle from 5 adult patients with DM25. In our study, 33 differentially expressed miRNA were identified in adult DM muscle compared with controls. Although these 33 miRNA were detected in our present study, none were significantly differentially expressed between untreated JDM and controls. This observation suggests that the miRNA profile in children with JDM is distinctive and differs from that in adults with DM. Another potentially important difference is that investigations of adult inflammatory myopathies may use biopsied material from treated as well as untreated patients25,26 as opposed to our study, in which all the children were untreated.
Published reports of microRNA data from muscle from adults with DM or PM show that dysregulated miRNA appeared to be associated with immune-related pathways such as leukocyte infiltration25. More recent miRNA data from adults (11 DM, 8 PM, 10 inclusion body myositis) identified decreased expression of miRNA-1, miRNA-133a, and miRNA-133b in all 3 types of inflammatory myopathy, as well as decreased expression of miRNA-206 in DM26. In contrast, these 3 miRNA were not dysregulated in the muscle of untreated children with JDM.
Study of miRNA-10a and miRNA-10b
We demonstrated that the expressions of a small set of unique miRNA were altered in inflamed muscle obtained from untreated children with JDM. The most significantly downregulated miRNA were miRNA-10a and miRNA-10b, which are encoded in the homeobox (Hox) genomic clusters and differ by 1 single nucleotide in the seed region32. The miRNA-10 family and Hox genes are coexpressed in the same genomic clusters on chromosomes 2 and 17, and appear to be critical regulators of maturation and development, including hematopoiesis33,34. We found a strong negative correlation between miRNA-10a and HoxA1 gene expression among the patients with JDM (data not shown). MiRNA-10a was reported to be upregulated in both tolerogenic and activated dendritic cells35, and dendritic cells have been identified in muscle of untreated JDM36,37. MiRNA-10a also regulates human cardiomyocyte progenitor cells38. We could not use any in vitro cell line or in vivo mouse models to further understand the potential mechanism of miRNA-10a in JDM because currently there is a lack of a representative in vitro cell line or in vivo mouse model for the vasculopathy present in children with JDM.
In our present study, the level of vWF:Ag in the JDM sera, reflecting endothelial cell damage, trended both toward significance (p = 0.07) and a negative correlation with miRNA-10a (p = 0.06). VWF:Ag has been reported as elevated in a proportion of children with JDM when their disease is active39,40, and in adults with DM, it is considered a marker of disease activity41. Vascular damage is a central feature of JDM pathophysiology5, raising the possibility that miRNA-10a might be a critical component of the JDM vasculopathy as well as contributing to the generation of premature atherosclerotic plaque. We had previously documented that older patients (mean age 38 yrs) with a history of JDM in childhood had evidence of premature atherosclerosis with a significantly increased intima media thickness (p = 0.015)18. Supporting evidence for the involvement of miRNA-10a in vascular damage comes from the observation that decreased levels of miRNA-10a were localized to the atherosusceptible endothelium of freshly isolated aorta from swine30.
MiRNA-10a is a regulator of the proinflammatory cytokine cascade and is augmented by increased levels of TNF-α
The present investigation documented that the cytokines MCP-1, IL-6, IL-8, and VCAM-1 were elevated in the untreated JDM sera at diagnosis. Previously published in vitro studies demonstrated that inhibition of endogenous miRNA-10a in conjunction with exposure to TNF-α resulted in increased phosphorylated IκBα30. MiRNA-10a interacts with MAP3K7 and β-TRC (each of which contains an evolutionary conserved miRNA-10a putative binding site), initiating the cascade of proinflammatory cytokines through NF-κB30. It is well established that in children with JDM, the A > G polymorphism in the TNF-α-308 promoter region is associated with increased production of the TNF-α protein both in vivo and in vitro by peripheral blood mononuclear cells16 and by the muscle fibers themselves42. In our present study, children with TNF-α-308A polymorphism had lower levels of miRNA-10a (p = 0.015).
Because of the limited muscle biopsy material available for study, we could not run gene expression profiles on the same children in our study. However, review of our published gene expression profiles of muscle from a different but very similar set of 19 females with untreated JDM and 4 controls confirmed upregulation of MAP3K7 and β-TRC by 1.67- and 1.85-fold, respectively (p < 0.01)29. In our present study, the downregulation miRNA-10a and miRNA-10b is equivalent in boys and girls with JDM (Table 3).
Clinical associations with miRNA data
Of the 2 isoforms, miRNA-10a, but not miRNA-10b, was associated with JDM clinical features. Specific clinical and laboratory features reflecting JDM disease activity and chronicity — vWF:Ag levels and the DAS Total — displayed a trend of association with the miRNA-10a expression levels, but not with miRNA-10b. Thus, miRNA-10a may contribute to the mechanism of vascular endothelial cell damage in children with JDM. We had previously documented that children with a short duration of untreated disease (≤ 2 months) had a decrease in miRNA-126 (−3.39-fold, p = 0.006)27. In our present study, because of lack of appropriate patient sample availability, the lower limit of a short disease duration was raised to 3 months (instead of 2 mos), and in this cohort, miRNA-126 was not decreased, thus suggesting that miRNA use in this inflammatory myopathy may be fluid and time-sensitive, as it is in other evolving types of aging43.
The small volume of muscle samples for experimental testing precluded extending our study. Further, parents of young otherwise healthy orthopedic patients were reluctant to donate muscle tissue for research, whereas older children who were otherwise healthy orthopedic patients were excited to participate as controls. Although age could be a potential confounding variable for the differential expression of miRNA-10a, miRNA-10b, and cytokines between JDM and controls in our pediatric study sample, we observed no significant correlation (p > 0.1) between age and any of those molecules among the JDM samples.
MiRNA profiles of muscle biopsies from children with untreated JDM document that the downregulation of miRNA-10a was augmented by the increased local TNF-α production as a consequence of the TNF-α-308A polymorphism. This downregulation was accompanied by overproduction of a range of NF-κB–controlled inflammatory mediators including IL-6, IL-8, TNF-α, VCAM-1, and MCP-1. Children with JDM express different spectra of miRNA than their adult counterparts with DM. We speculate that the vasculopathy characteristic of JDM is a direct consequence of the downregulation of miRNA-10a, and that the administration of miRNA-10a mimics might provide a new and effective strategy for the treatment of children with active JDM. We await confirmation, either in a larger patient cohort, or by in vitro testing or both.
ONLINE SUPPLEMENT
Supplementary data for this article are available online at jrheum.org.
Footnotes
Supported by US National Institutes of Health NR012692 and the Cure JM Foundation.
- Accepted for publication August 17, 2015.
REFERENCES
- 1.
- 2.
- 3.
- 4.
- 5.
- 6.
- 7.
- 8.
- 9.
- 10.
- 11.
- 12.
- 13.
- 14.
- 15.
- 16.
- 17.
- 18.
- 19.
- 20.
- 21.
- 22.
- 23.
- 24.
- 25.
- 26.
- 27.
- 28.
- 29.
- 30.
- 31.
- 32.
- 33.
- 34.
- 35.
- 36.
- 37.
- 38.
- 39.
- 40.
- 41.
- 42.
- 43.