
Abstract
Objective. Rheumatoid arthritis (RA) is the most common chronic, autoimmune, inflammatory disease, with a genetic and hormonal background. The prevalence of women among patients with RA suggests the important role of sex hormones in the pathogenesis of RA. We examined the association between CAG repeat polymorphism in the androgen receptor (AR) gene and susceptibility to RA and its clinical features in white women.
Methods. The study groups consisted of 325 female patients with RA and 238 female controls. CAG repeat polymorphism was determined using polymerase chain reaction and subsequent fragment analysis by capillary electrophoresis.
Results. The number of CAG repeats in patients did not differ from that of controls (22.1 ± 2.9 vs 21.9 ± 2.9, respectively; p = 0.26), but the presence of articular erosions was associated with a lower number of repeats in the shorter allele of patients with RA (20.4 ± 2.2 vs 21.2 ± 2.4; p = 0.031). When alleles with < 22 CAG were classified as short (S) and those with ≥ 22 CAG as long (L), the age at diagnosis of RA was lower in women with S-S genotype in comparison to combined S-L + L-L genotypes (43.0 ± 14.6 yrs vs 47.6 ± 12.5 yrs; p = 0.021). In patients with the L-L genotype, the frequency of erosive disease (OR 0.45, 95% CI 0.25–0.80, p = 0.0085) and extraarticular manifestations (OR 0.50, 95% CI 0.26–0.98, p = 0.047) was lower in comparison to carriers of the S allele. In multivariate analysis, the L-L genotype was an independent factor associated with a lower risk of erosions (OR 0.44, 95% CI 0.22–0.90, p = 0.024).
Conclusion. The results suggest the association of short AR (CAG)n alleles with earlier onset and a more aggressive course of RA.
- ANDROGEN RECEPTOR
- POLYMORPHISM
- RHEUMATOID ARTHRITIS
Rheumatoid arthritis (RA) is a multifactorial disease with a complex pathogenesis. There is a female preponderance in the prevalence of RA and the role of sex hormones such as androgens has been discussed1. Studies have shown increased levels of estrogens and decreased levels of androgens in synovial fluids from patients with RA. It is caused by enhanced conversion of androgens to estrogens through activation of aromatase in patients with RA2. Sex hormones can exert local actions in the tissues. An accelerated peripheral metabolic conversion of androgen precursors (i.e., testosterone) to 17ß-estradiol and conversion to other estrogenic metabolites (i.e., 16α-hydroxyestrone) is observed in patients with RA3. The effects of androgens depend on the plasma concentration available for tissue uptake and the interaction of androgens with the androgen receptor (AR)4. The AR is expressed in many tissues during development and adulthood, influencing an enormous range of physiologic processes including organ and tissue growth and differentiation, cognition, muscle hypertrophy, bone density, and insulin levels5.
The human AR gene, located on the X-chromosome at Xq11.2-q12, encodes protein of the nuclear receptor family of ligand-activated transcription factors. Its regulation of transcription is crucial for androgenic effects in individuals of both sexes6. The AR has 3 major functional domains: the N-terminal transactivation domain, the DNA-binding domain, and the C-terminal ligand-binding domain. The androgen-AR complex activates the expression of other genes (transactivation)7. The N-terminal domain of the protein, encoded by exon 1, is an important determinant of the transactivating capacity. It contains a glutamine repeat encoded by (CAG)nCAA sequence8. The normal length of the CAG repeat is in the range 10–30 repeats (mean 22 repeats) in white populations9. The polyglutamine tract length is inversely correlated to the transcriptional competence of the receptor, longer tracts being associated with lower levels of AR-mediated transcription in both normal and disease states10. Shorter CAG repeats have been associated with increased risk of prostate cancer, adverse plasma lipids, and male-type alopecia11,12. Longer repeats reportedly have been linked to clinical disorders characterized by reduced androgenic function: male infertility, hypogonadism, gynecomastia, and cryptorchidism13,14. The extreme elongation of the CAG repeats to > 40 triplets causes a disorder known as spinal and bulbar muscular atrophy or Kennedy’s disease15, in which AR protein aggregation in the cells occurs, finally leading to the death of motor neurons in the brain and spinal cord.
The altered estrogen production and altered androgen/estrogen balance associated with CAG repeat length may have functional importance in RA16. To date there are only a few studies examining the CAG repeat polymorphism in the AR gene in RA, and the results are inconsistent17,18,19,20.
The aim of our study was to examine the association between CAG repeat length and susceptibility to RA and its clinical features in white women.
MATERIALS AND METHODS
Subjects
The study group consisted of 325 female patients with RA (mean age 57.74 ± 12.52 yrs, age at disease diagnosis 46.52 ± 13.11 yrs), diagnosed according to the criteria of the American College of Rheumatology21. The patients were recruited from the Department of Rheumatology, County Hospital, Szczecin, Poland. All subjects were white, from the Pomeranian (northwestern) region of Poland. In the course of RA, 46.8% of them were treated with methotrexate, 44.6% sulfasalazine, 32.6% leflunomide, 22.5% glucocorticosteroids, 13.8% chloroquine, 4.6% azathioprine, and 2.5% cyclophosphamide. Subjects in the study underwent routine biochemical blood analysis and, when required, assays for anticardiolipin, antinuclear antibodies, and immunological complexes. Radiographs of the chest, hands, and feet were obtained from all patients and radiographs of other joints were performed when required, and interpreted by 2 expert radiologists. Evaluation of the subjects included physical examination, with particular focus on the pattern of joint involvement and the occurrence of extraarticular features (such as vasculitis, anemia, sicca syndrome, amyloidosis, organ involvement) and laboratory features such as rheumatoid factor (RF). Amyloidosis was diagnosed by histomorphology (skin and bowel or duodenum biopsy), vasculitis by histomorphology (skin biopsy) and angiogram. Severity of RA was evaluated using the Disease Activity Score for 28 joints (DAS28)22; the active form of RA was diagnosed when DAS28 was > 2.4, and the nonactive form when DAS28 was ≤ 2.423. We also performed subgroup analysis with regard to the premenopausal compared to postmenopausal age at diagnosis of RA, based on data for the mean age of menopause in the Polish population, 49.8 ± 2.3 years24. The associations between AR CAG polymorphism and RA were analyzed for women with mostly premenopausal onset of RA (RA diagnosis age < 45 yrs) and for women with mostly postmenopausal onset of RA (RA diagnosis age > 55 yrs). The control group consisted of 238 white women without autoimmunological diseases (mean age 68.84 ± 12.38 yrs), selected randomly from the population of the Pomeranian region of Poland. The study was approved by the local ethics committee and written informed consent was obtained from all subjects.
Genomic DNA was extracted from 200 μl samples of whole blood with the GeneMatrix Quick Blood DNA Purification Kit (EURx, Gdansk, Poland), and subsequently standarized to 20 ng/μl concentration using a NanoDrop ND-1000 spectrophotometer. The DNA region containing CAG repeats within the AR gene was amplified with a pair of flanking primers: 5′-TCC AGA ATC TGT TCC AGA GCG TGC-3′, 5′-ACT GCG GCT GTG AAG GTT GCT GT-3′25. Forward primer was fluorescently labeled with 6-carboxyfluorescein (6-FAM). Amplification was carried out in 15 μl volumes, using AmpliTaq Gold 360 Master Mix (Applied Biosystems, Foster City, CA, USA), 0.2 μM of each primer, and 1 μl genomic DNA, in 27-cycle polymerase chain reactions (PCR; primer annealing temperature 57°C). Length of the amplification product was subsequently analyzed by capillary electrophoresis: 1 μl of each sample was mixed with 12 μl Hi-Di Formamide (Applied Biosystems) plus 0.5 μl GeneScan 500 LIZ Size Standard, and processed in the ABI Prism 3130 Genetic Analyzer with an autosampler module. Electrophoresis was run on 36-cm capillary arrays filled with POP-4 polymer. Data were gathered with Data Collection v.3.0 software and analyzed with GeneMapper ID-X 1.1 software.
A number of CAG repeats corresponding to the length of amplified DNA fragments assessed by capillary electrophoresis were verified by sequencing of randomly chosen homozygous samples, using the same primers as described above. PCR product was initially purified using YM-100 centrifugal filters (Microcon, Millipore, Jaffrey, NH, USA) and sent for sequencing to a commercial service (oligo.pl, Warsaw, Poland).
Statistical analysis
The chi-square or Fisher’s exact test was used to compare genotype and allele frequencies between the groups and to analyze associations of clinical characteristics of RA patients with the genotype. Age at RA diagnosis was compared between genotype groups with the Kruskal-Wallis test followed by the Mann-Whitney U test. P < 0.05 was considered statistically significant.
RESULTS
The observed AR (CAG)n allele frequencies for cases and controls are presented in Figure 1. The androgen receptor CAG repeats were in ranges 11–34 and 12–30, respectively (Table 1). The frequency distribution of AR genotypes was in Hardy-Weinberg equilibrium (HWE) in the RA group (p = 0.91), but it was slightly different from HWE in the control group (p = 0.02). The mean number of CAG repeats for all alleles was similar in the patients (22.1 ± 2.9) and controls (21.9 ± 2.9). We also analyzed the shorter and the longer allele CAG repeats separately and found no significant difference between groups. Additionally, for each subject the mean number of repeats was calculated from the formula (shorter allele CAG repeats + longer allele CAG repeats)/2. The median value of the mean CAG repeats was 22 both for women with RA and for controls (Table 1).
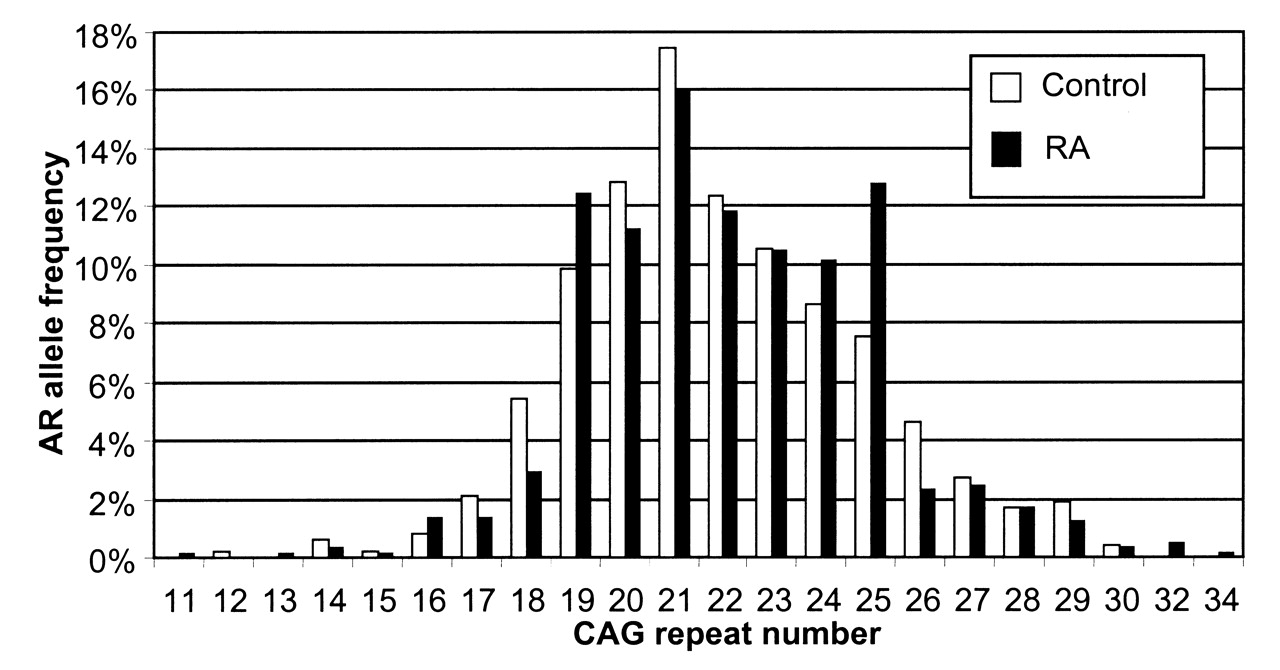
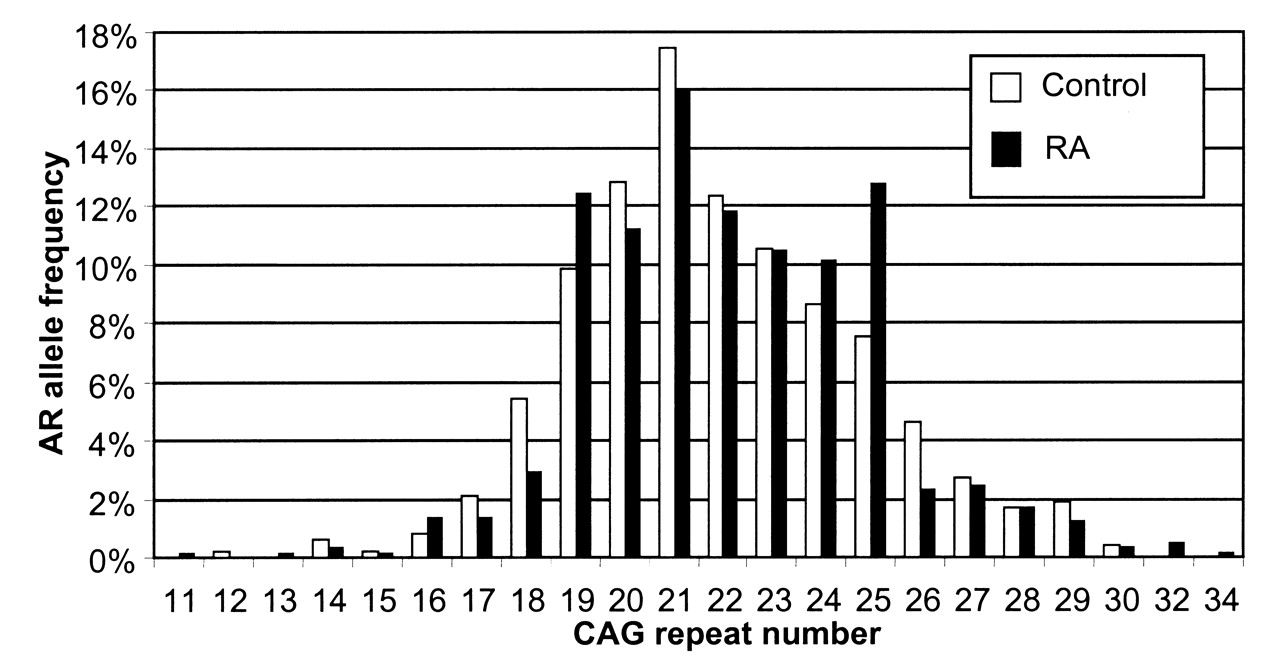{kind=link}
AR (CAG)n allele frequency distribution in patients with rheumatoid arthritis (RA) and controls.
The number of AR CAG repeats in patients with rheumatoid arthritis (RA) and controls.
Table 2 presents the AR genotypes and alleles classified according to median length in patients with RA and in controls. Alleles with < 22 CAG were classified as short (S) and those with ≥ 22 CAG as long (L). The subjects could thus be divided into 3 groups: those with 2 short alleles (S-S), those with 2 long alleles (L-L), and those with 1 short and 1 long allele (S-L). There were no significant differences in frequency of the 3 genotypes (S-S, S-L, L-L) or with S and L alleles between women with RA and controls.
AR genotypes and alleles classified according to the median CAG repeat length in patients with RA and controls.
The additional stratification of patients with RA and controls (Table 3) was performed according to the median of CAG repeat length in the shorter AR allele (n = 20), the longer AR allele (n = 23), and the mean length of the 2 alleles (n = 22). This analysis revealed no statistically significant differences.
Stratification of patients with RA and controls according to the median CAG repeat length of shorter AR allele, longer AR allele, and their mean.
We also analyzed the clinical measures of RA with regard to CAG repeat length (Table 4). The ages at RA diagnosis were lower in women with CAG genotype S-S in comparison with the combined S-L + L-L genotypes (p = 0.021). Patients with erosive RA had lower number of AR CAG repeats in the shorter allele (Table 5). Additionally, in patients with erosive disease a decreased frequency of L-L genotype was observed in comparison to patients without erosions (p = 0.0085, OR 0.45, 95% CI 0.25–0.80; Table 6). Further, a lower frequency of L-L genotype was observed in patients with extraarticular manifestations compared to patients without them (p = 0.047, OR 0.50, 95% CI 0.26–0.98).
Age at RA diagnosis and duration of the disease in patients stratified according to AR genotypes.
The number of AR CAG repeats in patients with rheumatoid arthritis (RA) stratified according to the presence of erosive RA, rheumatoid factor, and extraarticular manifestations.
AR genotypes and alleles in patients with RA stratified according to presence of erosive RA, rheumatoid factor, and extraarticular manifestations, and RA severity.
The mean of shorter and longer allele length > 22 CAG was associated with decreased frequency of erosive disease (Table 7). In the group with mean number of CAG repeats > 22, erosive disease was diagnosed in 75.3% of women, whereas in the group with mean CAG number ≤ 22, erosive RA was present in 85.7% of patients (p = 0.022, OR 0.51, 95% CI 0.29–0.90).
Stratification of rheumatoid arthritis (RA) patients with and without erosive RA, rheumatoid factor, extraarticular manifestations, and severe form of RA according to the median CAG repeat length of shorter AR allele, longer AR allele, and their mean.
To determine whether the AR genotype is an independent risk factor for erosive RA, multivariate analysis was performed, taking into account the duration of disease, age at RA diagnosis, and presence of the L-L genotype. The L-L genotype was found to be an independent factor associated with lower risk of erosions, while longer duration of disease was associated with higher risk of erosions in patients with RA (Table 8).
Multiple logistic regression model for the risk of erosive RA in patients with RA.
Additionally, we analyzed the patients with regard to premenopausal compared to postmenopausal onset of disease. There were no significant differences between subjects with RA diagnosis at age < 45 years and > 55 years, or between these subgroups and control subjects with regard to AR CAG genotypes and allele lengths (data not shown). The lower risk of erosive RA associated with mean number of CAG repeats in shorter and longer alleles > 22 was similar in the groups with age of RA diagnosis < 45 years (OR 0.33, 95% CI 0.11–0.98, p = 0.045) and those with age > 55 years (OR 0.39, 95% CI 0.14–1.08, p = 0.078).
DISCUSSION
We examined the associations between the CAG repeat polymorphism in exon 1 of the AR gene and RA in women. There were no statistically significant differences with regard to the number of CAG repeats between women with RA and controls, whereas we observed that the age at diagnosis of RA was lower in women with the CAG S-S genotype comprising 2 short alleles (< 22 CAG). Moreover, the frequency of erosive disease and extraarticular manifestations was lower in patients with the L-L genotype carrying 2 long alleles (≥ 22 CAG) in comparison to women with S-S + S-L genotypes.
To date, the length of the CAG repeat polymorphism of androgen receptor in RA has not been widely investigated and the results are inconsistent. Japanese investigators found no significant associations between CAG repeat length and RA severity indexes, such as seropositivity and rheumatoid nodule positivity17. However, the shorter CAG repeats of the AR gene were related to younger age at onset in men with RA. Yu, et al18 investigated the relationship between (CAG)n polymorphism of the AR gene and RA in Taiwanese female patients with different disease subtypes. Mean and median values of CAG repeats among cases with refractory RA and remission RA and controls did not differ significantly. However, patients harboring any long CAG alleles with more than 23 repeats had an increased risk of a refractory disease course. CAG repeat length was found to have no significant association with RF18. In another study, Lo, et al19 revealed no significant difference in the mean number of CAG repeats of the AR gene between Taiwanese patients with RA and controls, as well as no association between this polymorphism and extraarticular manifestations or joint erosion. Rheumatoid factor-seropositive female patients had significantly longer CAG repeats compared with seronegative female patients19. There was only one report concerning AR polymorphism in a white population with RA. The authors found no significant association between length of the CAG repeat and risk of RA20.
There were a few reports concerning (CAG)n polymorphism and other bone and joint diseases. The mean number of CAG repeats was higher in control women than in osteoarthritis (OA) cases. Additionally, women with the L-L genotype had lower risk of OA compared to individuals with the S-S genotype26. Data on the effects of AR CAG repeat length on bone mineral density (BMD) are inconsistent. Studies in European populations have produced all the 3 possible patterns: no correlation27, negative correlation28,29, and positive correlation29 between the number of AR CAG repeats and BMD.
CAG length [(CAG)n] is inversely associated with the transcriptional activity of the AR such that having longer (CAG)n reduces the transcriptional activity of the AR and subsequent intracellular androgenic activity7,30. Longer (CAG)n has been linked with lower testosterone levels in both premenopausal and postmenopausal women in some publications31,32, but a few studies reported no association of (CAG)n with testosterone levels33,34. Lower testosterone levels have also been noted in women treated with AR antagonists35,36, supporting some mechanism linking lower AR activity with lower circulating androgen concentrations in women.
Clinical studies suggest that androgens exert antiinflammatory activities in synovial tissue, which contrast with the activities locally exerted by estrogens2. Serum testosterone levels are inversely correlated with RA disease activity3. However, adrenal and gonadal androgens, which exert anti-inflammatory activities, are significantly decreased in inflamed tissues in both male and female patients with active RA. This observation supports a proinflammatory milieu at least in RA joints37. The presence of functional receptors for glucocorticoids, androgens, and estrogens in synoviocytes might link inflammation and the endocrine system38. Interestingly, increased aromatization of androgens has been demonstrated in cultured synovial cells from patients with RA, and the synthesized estrogens are further converted to proliferative estrogens, such as the 16-hydroxylated forms of estrone and 17ß-estradiol39. However, previous studies did not reveal unambiguously an association between AR CAG polymorphism and androgen concentrations in serum and tissues or with the clinical course of RA.
The AR is located on the X-chromosome, which is randomly inactivated in women31. Skewed inactivation is defined as inactivation of the same allele in more than 80% of cells examined40. It seems to be an inherited feature linked to specific loci of the X-chromosome41. In our study, we did not perform X-chromosome inactivation analysis similarly to other investigators examining AR CAG polymorphism in RA. Calvo, et al found that skewed inactivation of the AR is rare, but more importantly the shorter and longer CAG repeat sequence was subjected to skewed inactivation with similar frequency42. Therefore it seems that the lack of data concerning skewed inactivation in our subjects has no significant influence on the results.
Our study was the first, to our knowledge, to examine the association between the length of AR CAG repeat polymorphisms and the risks for and clinical measures of RA in women from a white population. With regard to the number of CAG repeats, there were no statistically significant differences between patients with RA and controls, therefore AR (CAG)n polymorphism does not seem to play an important role in the development of RA. Our results indicated the younger age of disease onset in women with 2 short alleles, which is consistent with the younger age of RA onset in men with the short allele in a Japanese population17. Moreover, women with 2 long alleles had decreased risk of erosive disease and extraarticular manifestations. These results suggest the role of androgens and their receptors in the clinical course of RA disease, and the association of short AR (CAG)n alleles with earlier onset and more aggressive course. Further studies are needed to confirm the potential role of the androgen receptor polymorphism in the pathogenesis of RA.
Footnotes
-
Supported by Pomeranian Medical University.
- Accepted for publication August 23, 2011.
REFERENCES
- 1.
- 2.
- 3.
- 4.
- 5.
- 6.
- 7.
- 8.
- 9.
- 10.
- 11.
- 12.
- 13.
- 14.
- 15.
- 16.
- 17.
- 18.
- 19.
- 20.
- 21.
- 22.
- 23.
- 24.
- 25.
- 26.
- 27.
- 28.
- 29.
- 30.
- 31.
- 32.
- 33.
- 34.
- 35.
- 36.
- 37.
- 38.
- 39.
- 40.
- 41.
- 42.