

Abstract
Objective. To get a better understanding of inflammatory pathways active in the osteoarthritic (OA) joint, we characterized and compared inflammatory cells in the synovium and the infrapatellar fat pad (IFP) of patients with knee OA.
Methods. Infiltrating immune cells were characterized by flow cytometry in 76 patients with knee OA (mean age 63.3, 52% women, median body mass index 28.9) from whom synovial tissue (n = 40) and IFP (n = 68) samples were obtained. Pain was assessed by the visual analog scale (VAS; 0–100 mm). Spearman rank correlations and linear regression analyses adjusted for sex and age were performed.
Results. Macrophages and T cells, followed by mast cells, were the most predominant immune cells in the synovium and IFP, and were equally abundant in these tissues. Macrophages and T cells secreted mostly proinflammatory cytokines even without additional stimulation, indicating their activated state. Accordingly, most CD4+ T cells had a memory phenotype and contained a significant population of cells expressing activation markers (CD25+, CD69+). Interestingly, the percent of CD69+ T cells was higher in synovial than IFP CD4+ T cells. Preliminary analyses indicated that the number of synovial CD4+ T cells were associated with VAS pain (β 0.51, 95% CI 0.09–1.02, p = 0.02).
Conclusion. Our data suggest that the immune cell composition of the synovium and the IFP is similar, and includes activated cells that could contribute to inflammation through secretion of proinflammatory cytokines. Moreover, preliminary analyses indicate that synovial CD4+ T cells might associate with pain in patients with endstage OA of the knee.
Knee osteoarthritis (OA) is a joint disease characterized by structural damage and pain. During the last decade, it became clear that synovial inflammation is present in a significant number of patients with knee OA1,2,3. In several studies, synovial inflammation has been associated with pain4,5,6 and radiographic progression7,8,9 of the disease, indicating it as an important player in disease pathogenesis.
Signal alterations on (contrast-enhanced) magnetic resonance imaging are used as a surrogate biomarker for inflammation. Signal alterations have been reported both at the synovial and the infrapatellar fat pad (IFP) level, in addition to effusion, and both have been reported to be associated with radiographic damage10,11 and pain4,5,6,12. However, it is still unclear how these signal alterations relate to histological changes and cellular infiltrate in these 2 tissues, and whether inflammatory changes in the synovium and IFP are related. Moreover, the relative contribution of these 2 tissues to clinical variables such as radiographic damage and pain in OA remains to be established.
The inflamed synovium of patients with OA is infiltrated with immune cells, which can secrete various inflammatory mediators (review13) that could underlie the association between inflammation and pain/structural damage. Some studies indicate that cells in the OA synovium could be in an activated state based on surface marker expression (such as CD25 or CD69 for T cells) or the presence of soluble mediators known to be secreted by activated macrophages [i.e., cytokines such as interleukin (IL)-6, tumor necrosis factor-α (TNF-α), etc.] or mast cells (i.e., histamine). However, the limited number of studies as well as the small number of samples used that support the findings suggest that followup studies are necessary to fully characterize the activation state of immune cells in the synovium. Moreover, the relationship between the (phenotype of) immune cells in the synovium and the clinical disease characteristics has not yet been fully investigated.
Another source of immune cells and inflammatory mediators in the joint is the IFP14,15,16. Because the IFP is situated adjacent to the synovium, it is conceivable that crosstalk exists between these tissues and that the IFP could contribute directly or indirectly to disease pathogenesis.
To get a better insight into inflammatory pathways operational in the OA joint, we determined the abundance and phenotype of the main immune cell populations in the synovium and compared these with the IFP. Further, we identified the association of different immune cell populations with pain in a hypotheses-generating setting using a well-defined cohort of patients with knee OA.
MATERIALS AND METHODS
Patients and study design
Patients with OA (n = 76) undergoing joint replacement surgery were recruited into our study, and synovial (n = 40) and IFP samples (n = 68) were obtained and immediately processed. In total, 39 paired samples were obtained.
Forty-two of the patients included in our present study were part of the observational geMstoan study (GEneration of Models, Mechanism & Markers for STratification of OsteoArthritis patieNts) of patients with knee OA that aimed to find new biomarkers for OA progression. The geMstoan patients were included between 2008 and 2013, had symptomatic radiographic primary knee OA following the American College of Rheumatology criteria17, and attended the Orthopedic Department of the Leiden University Medical Center (LUMC) or the Orthopedic Department of the Diaconessenhuis in Leiden. In our study, patients with other rheumatic diseases treated with immunosuppressive drugs or receiving knee injections (corticosteroids, etc.) in the past 3 months were excluded. Written informed consent was available from all geMstoan patients.
From 34 patients, leftover synovial tissues and IFP samples were obtained during total knee arthroplasty, thus no additional clinical data were available besides diagnosis, age, sex, and body mass index (BMI). Both studies were approved by the ethics committee of the LUMC.
Isolation of synovial cells and the IFP-derived stromal vascular fraction (SVF)
Synovial tissue was cut and subsequently digested with collagenase type II (Sigma) for 90 min at 37°C under continuous rolling. The digested tissue was filtered through a cell strainer with a pore size of 70 μm.
Isolation of SVF cells from the IFP was performed as previously described14. Briefly, tissue fragments were incubated with 1.8 mg/ml collagenase type 1A (Sigma-Aldrich) in RPMI 1640 (Gibco) for 1 h at 37°C under continuous rolling. The digested tissue was filtered through a nylon mesh with a pore size of 250 μm, and the remaining tissue was mashed and washed through a 70-μm nylon mesh. The cells suspension was centrifuged and the supernatant containing floating adipocytes was removed. The SVF cells were washed 3 times in warm DMEM 4.5 g/l glucose/F12/0.5% bovine serum albumin/15 mM Hepes/glutamax/pen/strep (Gibco; medium). Erythrocytes were lysed by adding lysis buffer (0.154 mmol/l NH4Cl, 5.7 mmol/l K2HPO4, and 0.1 mmol/EDTA pH 7.3) for 5 min at room temperature.
Both the cells isolated from the synovium and the SVF fraction were further stained with the appropriate antibodies.
Flow cytometric analysis
For surface staining, 100,000 synovial or SVF cells were stained with mixes of the following antibodies: phycoerythrin (PE)-conjugated CD3; fluorescein isothiocyanate (FITC)-conjugated CD45, CD8, and CD45RA; Pacific Blue (PB)-conjugated CD4; allophycocyanin (APC)-conjugated CD117 and CD8; and PE-CY-7–conjugated CD14 and CD25 (all antibodies were from BD Biosciences). All incubations were performed at 4°C for 30 min.
For intracellular cytokine staining, synovial or SVF cells were plated overnight in medium containing 50 IU/ml IL-2 in the presence or absence of 3 μg/ml brefeldin A (to block secretion of proteins). The next day, cells cultured in the absence of brefeldin A were activated with 20 ng/ml phorbol myristate acetate (PMA) and 200 ng/ml ionomycin for 5 h, and 10 μg/ml brefeldin A was added in the last 4 h of culture. Cells cultured in the presence of brefeldin A were stained directly the next day. About 400,000 synovial or SVF cells were stained using the BD intracellular cytokine fixation/permeabilization solution kit (BD Biosciences) according to the manufacturer’s instructions. The following antibodies were used: Alexa Fluor 700-conjugated CD3; PB-conjugated CD4; APC-conjugated CD8; FITC-conjugated CD45RA; PE-CY-7–conjugated CD14 and IL-4; and PE-conjugated interferon-γ (IFN-γ), TNF-α, IL-10, and IL-6 (all from BD Biosciences). Cells were fixed with 1% paraformaldehyde and analyzed with an LSR II flow cytometer using Diva 6 software (BD Biosciences). Dead cells were excluded based on both forward scatter (FSC)/side scatter (SSC) and staining with the dead cell discrimination kit (Miltenyi Biotec), as indicated. Single cells were selected based on FSC/SSC analyses, as described in the gating strategy (Supplementary Figure 1 is available from the authors on request).
Clinical data
In the geMstoan patients, demographics and disease characteristics were collected through standard questionnaires. Measurement of pain in the affected knee was performed preoperatively using a general assessment of self-reported pain by the visual analog scale (VAS; 0–100), a 1-dimensional measure of pain intensity in which a score of 100 mm represents worst possible pain.
Statistics
Paired Wilcoxon signed-rank test was used to compare percentages of immune cells between tissues. Association between percentage of immune cells and VAS pain was determined by linear regression analyses, and was adjusted for age and sex when necessary. Correlations were determined by the Spearman rank test. A correlation < 0.3 was considered weak, 0.3–0.7 moderate, and > 0.7 strong. A p value < 0.05 was considered significant. The Statistical Package for the Social Sciences version 20.0 (SPSS) was used for analyses.
RESULTS
Immune cell populations in the synovium and the IFP
First, we characterized the immune cell population in the synovium in detail. Because of the limited amount of tissue, not all experiments could be performed with each tissue sample. Therefore, variable numbers of samples were included in different experiments.
Flow cytometric analysis of the synovial cells revealed a heterogeneous forward/side scatter (FSC/SSC), indicating the presence of several cell types (Supplementary Figure 1 is available from the authors on request). To determine the abundance of various cell types, we used specific cell-surface markers and CD45 to identify hematopoietic cells. Macrophages (CD14+), T cells (CD3+), and mast cells (CD117+) were readily detectable (Figure 1A–C), while B cells (CD19+ cells) were virtually absent (< 1%; data not shown). Further phenotypic characterization of CD4+ and CD8+ T cells revealed that these populations consisted mainly of memory cells (do not express CD45RA; Figure 1D). Interestingly, a substantial percentage of both CD4+ (T helper cells) and CD8+ (cytotoxic T cells) T cells had an activated phenotype and expressed CD25 and CD69 (Figure 1D). Similar data were obtained when the SVF cells from the IFP were analyzed (Figure 1A–D).

Immune cell characterization of the synovium (closed black symbols) and the stromal vascular fraction of the IFP (open symbols) of all patients with endstage OA (n = 76) by flow cytometry. Gating strategies were performed as described in Supplementary Figure 1 (available from the authors on request). Percentages of (A) macrophages (CD45+CD14+) and mast cells (CD45+CD117+), and (B) T cells (CD3+) are depicted. Percentages of (C) CD4+ and CD8+ T cells, and (D) CD45RA, CD25, and CD69 expressions within the CD4+ and CD8+ T cell populations are indicated. Mean percentage positive cells ± SEM are depicted. Each dot represents 1 patient. syn: synovium; IFP: infrapatellar fat pad; OA: osteoarthritis; SEM: standard error of the mean.
To detect possible tissue-specific effects and to directly compare the prevalence of different cell populations in the synovium and IFP, we have selected and compared paired synovium-IFP samples (Figure 2). We found no differences in the percentages of T cells (Figure 2A), macrophages (Figure 2B), mast cells (Figure 2C), memory T cells, or T cells bearing the late activation marker CD25 (data not shown) between tissues. However, percentages of T cells positive for the early activation marker CD69 were significantly higher in synovium-derived CD4+ T cells compared with IFP-derived T cells (Figure 2D). This difference was likely not due to differences in the collagenase treatment between the synovium and IFP because the treatment of peripheral blood mononuclear cells for 1 h or 1.5 h using collagenase type IA or type II did not induce CD69 expression on CD4+ T cells (data not shown).

Paired syn-IFP samples analysis of different cells population for patients with endstage knee OA. Paired analysis of (A) T cells (percent of CD3+ T cells of CD45+ cells, n = 10), (B) macrophages (CD14+CD45+, n = 15), and (C) mast cells (CD117+CD45+, n = 15) are depicted. Further, (D) percentages of T cells (CD4+ and CD8+ T cells) positive for the early activation marker CD69 are shown (n = 9). Differences between groups were assessed using the paired Wilcoxon signed-rank test. syn: synovium; IFP: infrapatellar fat pad; OA: osteoarthritis.
T cell and macrophage cytokine production
Because macrophages and T cells are the most abundant immune cell types present in the synovium, we further characterized these cells. To assess their possible contribution to inflammatory mediators, we investigated the polarization state of these cells by studying their cytokine secretion. Cytokines typically associated with different polarization states were investigated. Gating strategies are described in Supplementary Figure 1B (available from the authors on request). Synovial macrophages secreted IL-6 and TNF-α, but also some IL-10 ex vivo, in the absence of extra stimulation (Figure 3A). Intracellular cytokine staining showed that both the CD4+ and CD8+ T cell subsets produced mainly IL-6 and IL-4 (Figure 3C) in the absence of extra stimulation. Upon in vitro stimulation using PMA in combination with ionomycin (PMA/ionomycin), these cells were able to produce IFN-γ, TNF-α, IL-6, IL-4, and very little, if any, IL-10 (Figure 3E). Comparable results were obtained for macrophages and T cells from the IFP (Figure 3B, 3D, and 3F).

Macrophage and T cell cytokine production in the synovium (A, C, E) and the stromal vascular fraction of the IFP (B, D, F). Gating strategies were performed as described in Supplementary Figure 1 (available from the authors on request). Spontaneous cytokine production by CD14+ macrophages in (A) the synovium or (B) the IFP is depicted. Likewise, spontaneous (C, D) or PMA/ionomycin-induced cytokine production (E, F) of CD4+ and CD8+ T cells are depicted. Indicated are mean percentage cytokine positive cells ± SEM. Each dot represents 1 patient. IFP: infrapatellar fat pad; PMA: phorbol myristate acetate; SEM: standard error of the mean; IL: interleukin; TNFα: tumor necrosis factor-α; IFN-γ: interferon-γ.
Association of immune cells with clinical characteristics
To get first insight into the possible contribution of immune cells to clinical characteristics of OA such as pain, we next investigated whether the synovial immune cell populations were associated with self-reported knee pain. For this, we made use of the clinically well-characterized population of patients participating in the geMstoan study (n = 42). No significant differences in the immune cell populations or clinical characteristics were observed between the geMstoan and non-geMstoan samples (data not shown). Patient characteristics are shown in Table 1. The median level of VAS pain was 68 and ranged from 32 to 95. The percentage of CD4+ T cells in the synovium was associated with VAS pain, after adjustment for age and sex (Table 2, Figure 4A). No such association was found for other immune cells (Table 2). Moreover, no association was found for IFP-derived T cells with pain (Figure 4B), nor for other IFP-derived cells (Table 2). Because this part of our study was meant as hypotheses-generating, we did not adjust for multiple testing.
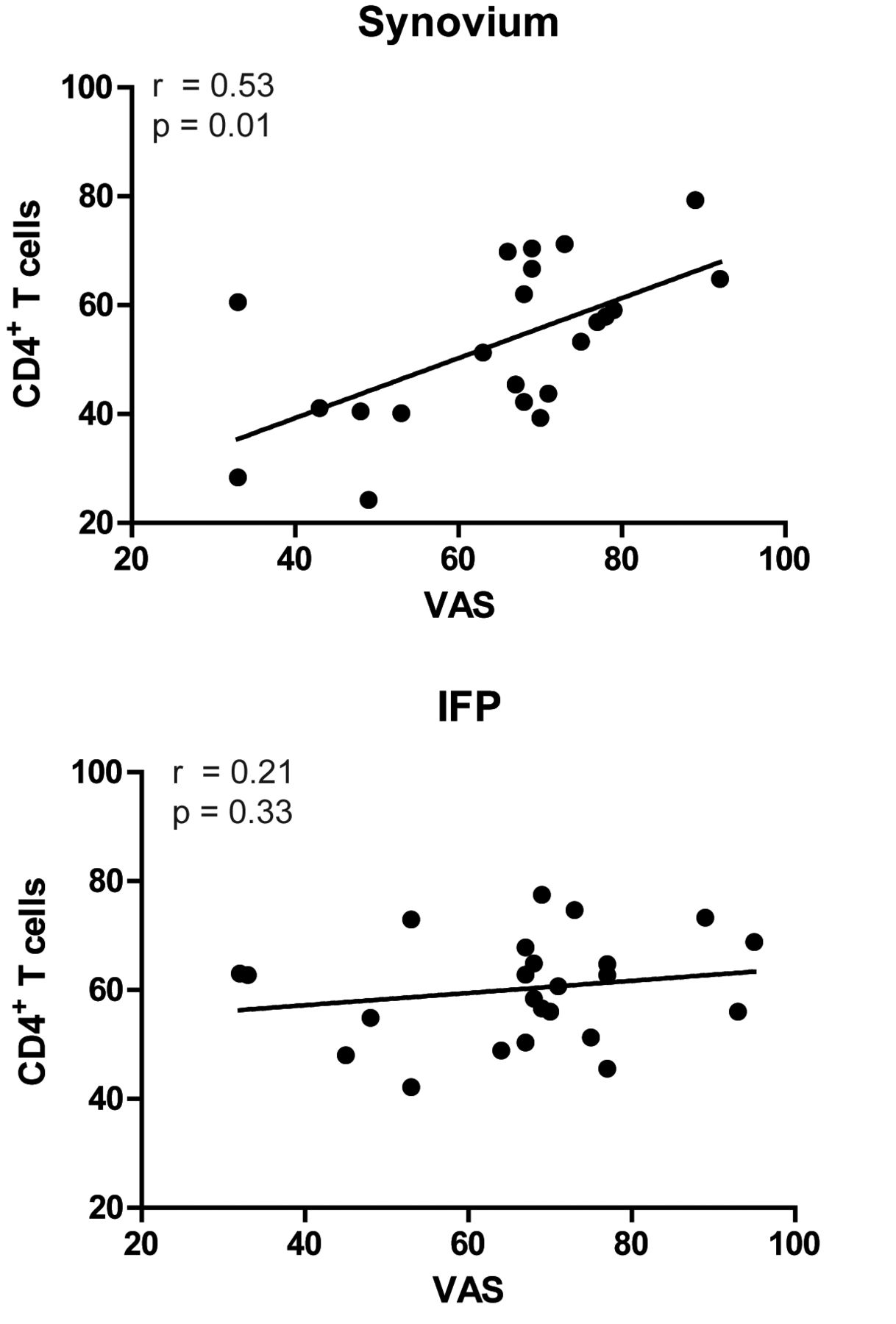
Spearman rank correlations between the synovial and IFP-derived CD4+ T cells and VAS. A p value ≤ 0.05 was considered significant. IFP: infrapatellar fat pad; VAS: visual analog scale.
geMstoan study patient characteristics (n = 42). Values are mean (SD) unless otherwise specified.
Association of various immune cells in synovium and IFP with knee pain, assessed by the visual analog scale in 42 patients with endstage knee OA.
Interestingly, both VAS pain (r = 0.34, p = 0.04) and CD4+ T cells in the synovium correlated significantly with the BMI (r = 0.45, p = 0.03) in our cohort. Therefore, we next investigated whether the association of CD4+ T cells with pain is, in statistical terms, mediated by the BMI. These analyses revealed that the association of CD4+ T cells with VAS pain was independent of the BMI (adjusted β 0.55, 95% CI 0.02–1.08).
In an attempt to get more insight into the mechanisms underlying this association, we investigated the relationship between different T cell characteristics and pain. Although the association between different cytokine-producing T cells and pain could not be studied because of the limited availability of cytokine data in the geMstoan population, we did investigate whether phenotypically distinct CD4+ populations correlated with pain. No correlations could be found between CD4+ T cells expressing CD45RA+, CD25+, or CD69+ with VAS in the synovium or the IFP (data not shown).
DISCUSSION
We investigated the immune cell populations present in the synovium and the IFP of patients with OA. Our data indicate similarities both in the composition and the phenotype of different immune cell populations in these tissues. Moreover, our exploratory studies investigating the association of different cell types with radiographic damage and pain suggest that synovial CD4+ T cells might be related to pain perception in patients with endstage knee OA.
Macrophages and T cells are the most predominant immune cell types present in the synovial tissue and IFP of patients with OA, which is in line with earlier observations13,14. To our knowledge, a direct comparison between the synovial cells and the IFP has never been performed before. We found that the immune cell composition of these tissues was very similar, with no significant differences in the cell type present and in the abundance of different cell populations. However, in a previously reported study14, we described differences in T cells and mast cells between paired IFP-subcutaneous adipose tissue samples from patients with OA14. Together, these data suggest that infiltration of immune cells into tissues might be more affected by the disease process (i.e., knee OA) than by tissue-specific signals. Moreover, these data support the findings16,18 that suggest the existence of crosstalk between synovial tissue and IFP, ultimately leading to a common inflammatory state. The involvement of the IFP in the development of OA requires further elucidation; however, the data reported here are intriguing in view of the existing contradictory literature related to the IFP contribution to the pathophysiology of OA, in which some authors suggested an antiinflammatory/beneficial function of IFP based on the inverse associations of the IFP maximal area with pain or structural damage19,20, while others identified it as a source of proinflammatory mediators with potential harmful effects in the joint (review21).
Our analyses further indicate that both tissues are an important source of inflammatory mediators and therefore could be involved in the pathophysiology of OA. Indeed, both macrophages and T cells could produce cytokines directly ex vivo, which suggests that they are in an activated state in the tissues. Moreover, they secreted predominantly proinflammatory and little or no antiinflammatory cytokines, an action in agreement with the diseased state and with our earlier finding that H&E staining of these tissue samples revealed the presence of at least 1 feature of synovitis (i.e., lining cell layer thickening, stromal activation, presence of cellular infiltrate) in each patient. This was in contrast with the synovial samples from patients with OA undergoing arthroscopy, in which several samples had no detectable signs of inflammation11.
Previous studies focused on the synovium (review13) indicate that CD4+ T cells have an activated phenotype, as indicated by the expression of CD2522,23,24 or CD6923,25,26. Our study indicates that these markers are expressed by T cells in both tissues. Interestingly, percentages of T cells positive for the early activation marker CD69 were significantly lower in IFP-derived compared with synovium-derived CD4+ T cells, indicating that T cells in the synovial tissue might be more frequently activated than in the IFP. In addition, T cells secreted cytokines ex vivo in the absence of extra stimulation. Because it is believed that T cells stop secreting cytokines when their cognate antigens are removed, and because the expression of CD69 is transient and relatively short-lived, it is conceivable that CD69/cytokine-secreting T cells represent recently activated cells that recognize antigens locally in the tissues from which they originate. This would be in line with previous data in which a lower expression of CD3ζ chain was observed on synovial T cells, indicative of chronic activation27. Although it is unclear whether T cells are locally activated and expanded or recruited to the tissue in an activated state, some data in mice indicate that CD4+ T cells could be involved in OA28. This hypothesis remains to be further investigated.
We observed a BMI-dependent infiltration of CD4+ T cells in the synovium (r = 0.45, p = 0.03), revealing an associative link between obesity and synovial inflammation, to our knowledge for the first time. Whether adipose tissue secreted–factors or other obesity-related changes underlie this association needs to be established.
Previous studies have indicated the presence of Th1 cells in synovial tissue. However, IFN-γ, the typical Th1 cytokine, proved difficult to detect by immunohistochemistry29,30 (review13), which might indicate that the Th1 cells present in this tissue are not in an activated state. Indeed, our data support this hypothesis because we could clearly identify IFN-γ–secreting Th1 cells upon ex vivo activation, but not in the absence of activation. In contrast, we could identify T cells secreting IL-6, a proinflammatory cytokine previously associated with OA31 and usually associated with innate immune responses, both in the synovium and IFP. Current studies are investigating the phenotype and functional relevance of this population of T cells.
We have also described an intriguing finding involving the association of pain perception with inflammatory cell infiltration in the OA synovial tissue. Although the number of available samples allows only for a hypotheses-generating study regarding the association of various infiltrating cell types with pain, we believe that the described association of synovial CD4+ T cells with pain is of interest and plausible in view of the literature. Several studies have implicated a potential involvement for T cells in nociception, although the exact mechanisms of their contribution to pain perception are not yet understood. T cells have been shown to infiltrate the site of nerve injury, and the elimination of T cells in nude rats attenuated hyperalgesia and allodynia in a model of neuropathic pain32. In addition, this effect appeared to be facilitated by Th1 cells through the release of IL-2 and IFN-γ32, cytokines that were also secreted by synovial CD4+ T cells in our study. This indicates a possible mechanism by which synovial T cells could be associated with pain in OA. Moreover, in our study, synovial CD4+ T cells also secreted TNF-α and IL-6. Interestingly, these cytokines have also been shown to directly affect sensory fiber function33,34,35,36, further supporting the hypothesis that CD4+ T cells could be associated with pain. Therefore, future studies replicating our preliminary findings are certainly of great interest.
A limitation of our study was the preferential inclusion of patients who underwent arthroplasty as the only possible source of IFP and of sufficient synovial tissue for exhaustive analyses. Therefore, all patients included in our present study had synovial inflammation and experienced pain in the presence of advanced radiographic knee OA, which could bias our data. Although no patients free of pain were included, the variation in the VAS scores (32–95) was sufficient to enable us to study associations between immune cells and pain. Other limitations include the lack of control subjects, either early OA or healthy individuals, because of technical or ethical difficulties, and the low number of samples available for cytokine analysis, precluding the study of the association between cytokines and certain subpopulations of immune cells (such as pro- and antiinflammatory macrophages) and pain measures.
Our study indicates that the inflammatory cell composition and phenotype is similar between the synovium and the IFP of patients with OA, and that part of the infiltrating cells have an activated phenotype and secrete proinflammatory cytokines. Moreover, while our data await replication, our study indicates that synovial CD4+ T cells could contribute to the perception of knee pain in patients with endstage knee OA, offering a cellular basis for the association between synovial inflammation and knee pain in patients with OA.
Acknowledgment
The authors acknowledge the support of the cooperating hospital Diaconessenhuis, Leiden, the Netherlands, and its orthopedic surgeons and nurse practitioners.
Footnotes
Work performed within the framework of Dutch Top Institute Pharma, project “Generation of models, mechanisms and markers for stratification of osteoarthritis patients” (project no. T1-213). Also financially supported by The Dutch Arthritis Association, the European Union 6th Framework Programme (FP6) Autocure, FP7 program Masterswitch, a grant from Centre for Medical Systems Biology within the framework of the Netherlands Genomics Initiative, and The Netherlands Organization of Health Research and Development.
- Accepted for publication January 18, 2016.








{kind=link}
{kind=link}
{kind=link}
{kind=link}