

Abstract
Objective. Interleukin 23 (IL-23) plays a major role in differentiation and survival of IL-17-secreting CD4+ Th17 cells. Having noted a higher frequency of Th17 cells in ankylosing spondylitis (AS) and rheumatoid arthritis (RA) than in healthy donors (HD), we investigated whether IL-23 secretion is increased in these conditions.
Methods. Monocyte-derived dendritic cells (moDC) were obtained from peripheral blood of 17 HD, 16 patients with RA, and 30 patients with AS, and stimulated with ligands for several pathogen recognition receptors. Messenger RNA (mRNA) expression and cytokine secretion were analyzed by real-time polymerase chain reaction and ELISA, respectively.
Results. The combination of ligands for Toll-like receptors (TLR) 7/8 and TLR3 led to synergistic secretion of both IL-23 and IL-12p70 from all subjects; similar synergy was seen with TLR2 ligands and curdlan. However, for both combinations, moDC from patients with RA produced significantly lower amounts of IL-23 than moDC from patients with AS; in contrast, IL-12p70 secretion did not differ. Similarly, tumor necrosis factor-α, IL-6, and IL-10 were secreted at comparable levels in all subjects, whereas CXCL8 and CCL3 production was actually enhanced in moDC of patients with RA. Equivalent levels of mRNA for both IL-23p19 and IL-12p35 subunits were found in moDC from all donors, suggesting posttranscriptional regulation of IL-23 production in RA.
Conclusion. Our observations show that IL-23 production is decreased in RA and maintained in AS. Because increased numbers of CD4+IL-17+ T cells are seen in both diseases, these observations imply that there are different mechanisms underlying chronic inflammation in these 2 forms of inflammatory arthritis.
Dendritic cells (DC) efficiently detect pathogens and cell damage through several pattern recognition receptors (PRR), including the Toll-like receptors (TLR). Activation of PRR results in the secretion of cytokines that affect the differentiation and function of both innate and acquired immune effector cells1. DC have been implicated in the pathogenesis of autoimmune diseases through presentation of self-antigens, and in perpetuation of chronic inflammation through cytokine production2,3; increased numbers of DC are also observed at inflammatory sites in various rheumatic diseases4,5,6.
Two of the cytokines affecting T cell differentiation are interleukin 23 (IL-23) and IL-12, and activated DC are a major source of these cytokines. IL-23 and IL-12 have unique subunits (p19 and p35, respectively) covalently linked to the common p40 subunit7. In vitro stimulation of naive T cells with IL-12 induces interferon-γ (IFN-γ)-producing Th1 cells, which act against intracellular bacteria. In contrast, IL-23, together with IL-1, IL-6, and IL-21, favors differentiation and expansion of Th17 cells, which can secrete IL-17, along with IL-22, IL-21, IL-6, and tumor necrosis factor-α (TNF-α), and confer resistance to extracellular bacteria and fungi8,9,10,11. The IL-23:IL-17 axis has also been implicated in murine models of autoimmunity; thus mice lacking IL-23p19 are resistant to collagen-induced arthritis, experimental autoimmune encephalomyelitis (EAE), and T-cell mediated colitis12,13,14. Moreover, recent findings have shown that Th17 cells can cease to produce IL-17 (ex-Th17 cells) and instead secrete IFN-γ and granulocyte-macrophage-colony stimulating factor (GM-CSF), suggesting plasticity in T cell subsets15. Again, IL-23 is important in this respect because, together with IL-1/IL-6, it results in differentiation of Th17 cells, which can later proceed to GM-CSF secretion. These Th17 cells are the subset responsible for EAE, in contrast to those differentiated by IL-6 + transforming growth factor-β16,17. Certain cells of the innate immune system, including some capable of producing IL-17, also express IL23 receptor (IL-23R) and can be driven to cytokine production by IL-23; such cells are particularly active in the gut18.
IL-23 has been implicated in human diseases; lipopolysaccharide (LPS)-stimulated DC from patients with multiple sclerosis were shown to produce increased levels of IL-2319. In psoriasis, where IL-23p19 is expressed in inflamed skin, a p40-specific monoclonal antibody has shown remarkable clinical efficacy20,21. Moreover, polymorphisms in the IL-23R gene are associated with susceptibility to AS22 and to psoriasis and Crohn’s disease, both commonly complicated by spondyloarthritis (SpA)23,24; a hypofunctional IL-23R allele decreases susceptibility to AS25. Increased numbers of Th17 cells have been reported in various forms of arthritis, including juvenile idiopathic arthritis, RA, AS, and psoriatic arthritis26,27,28. In RA, while synovial cells are reported to express the IL-23p19 subunit abundantly, levels of IL-23 in synovial fluid are low29,30,31.
We designed our study to determine whether monocyte-derived DC (moDC) from patients with AS and RA secrete higher levels of IL-23 than do moDC from healthy donors (HD) in response to PRR ligands, including TLR agonists and curdlan, a fungal β-glucan recognized through the C-type lectin receptor dectin-1 and the integrin CR3, together with dectin-232,33, and known to be a potent stimulator of IL-23 secretion34.
MATERIALS AND METHODS
Patients
Thirty patients with AS, 16 with RA, and 17 HD were recruited for our study. Written informed consent was obtained from all patients and HD, and the work was approved by the Cambridge Cambridgeshire 2 Research Ethics Committee (Ref: 01/363; date of approval: December 13, 2001, “The role of the immune system and its component in the pathogenesis of inflammatory arthritis”). Clinical data of all donors are reported in Table 1. Patients with AS met the modified New York criteria35 and those with RA fulfilled American College of Rheumatology revised criteria36.
Characteristics of patients and healthy donors (HD).
MoDC differentiation
Peripheral blood (PB) from all subjects was collected in heparinized tubes and mononuclear cells (MC) obtained by centrifugation over a Fycoll-Hypaque density-gradient. Monocytes were obtained from PBMC by positive selection using cluster of differentiation-14 (CD14) microbeads (Miltenyi Biotec) according to manufacturer’s instructions. Purity of CD14+ cells, assessed by flow cytometry, was above 97%. Monocytes were cultured in 24 well plates in RPMI medium (Gibco) containing 10% heat inactivated fetal bovine serum (Biosera), L-glutamine, 50 μg/ml gentamicin (Cydomicin; Aventis), and 10 mM HEPES buffer (Sigma-Aldrich), supplemented with recombinant GM-CSF (50 ng/ml; Biosource) and recombinant IL-4 (1000 U/ml; BD Pharmingen). At Day 6, immature moDC were harvested using cold phosphate buffered saline (PBS) and 2 mM EDTA, and counted with Trypan Blue to exclude dead cells. Four × 105/well viable cells were then placed in U-bottom 96-well plates. The phenotype of immature moDC (CD14−/low CD11c+ CD1c+) was checked by flow cytometry to ensure consistency of differentiation in each experiment.
MoDC stimulation with PRR ligands
Immature moDC were cultured without treatment or stimulated with various TLR ligands, and with a dectin-1 ligand, β-1,3-glucan, curdlan (125 μg/ml; Wako Chemicals), alone or in combination. The TLR ligands used were peptidoglycan from Bacillus subtilis (Sigma-Aldrich) 10 μg/ml, ultra-pure Escherichia coli 0111 LPS 1 μg/ml, poly(I:C) 30 μg/ml, Pam3CSK4, a synthetic triacylated lipopeptide 1 μg/ml, and CL097 5 μg/ml, all provided by Autogen Bioclear. After 24 h, supernatants were collected and stored at −20°C for cytokine measurement; poststimulation moDC were harvested to evaluate expression of the activation and maturation markers CD86, MHC class II, and CD83 using flow cytometry.
Flow cytometry
The antibodies used in the study were as follows: FITC-conjugated anti-CD14 (TüK-4; AbD Serotec), phycoerythrin (PE)-conjugated CD11c (3.9; Caltag Laboratories), allophycocyanin (APC)-conjugated CD1c (AD5-8E7; Miltenyi Biotech), and FITC-conjugated CD83 (clone HB15e) and PE-conjugated CD86 (clone 2331), both purchased from BD Bioscience. HLA-DR (L243), followed by incubation with an APC-conjugated goat anti-mouse secondary antibody, was used for staining MHC class II molecules. Stainings with isotype-matched control conjugated antibodies were performed to control for nonspecific binding for each fluorochrome. Cells were stained by incubation with antibodies at 4°C for 15 min in staining buffer [PBS, 0.1% bovine serum albumin (Sigma-Aldrich), and 0.01% sodium azide] and fixed with 2% paraformaldehyde. Cells were analyzed on a FACS Calibur flow cytometer (BD Bioscience) and data processed with WinMDI software.
Real-time quantitative polymerase chain reaction (RT-PCR)
Total RNA was extracted from cells 6 hours poststimulation, using RNeasy™ columns (Qiagen) as described by the manufacturer. The expressions of IL-23p19, IL-12p35, and IL-12/IL-23p40 were quantified as ratio of fold change differences in target gene expression between treated and untreated cells relative to those in reference hypoxanthine-guanine phosphoribosyltransferase mRNA. All RT-PCR probe primer sets were obtained from Applied Biosystems (Foster City, CA, USA), and their sequence is reported in a previous study37.
Measurement of cytokines by ELISA
IL-23p19/p40 heterodimer was evaluated using an eBioscience kit according to manufacturer’s instructions. For measurement of biologically active IL-12p70, a sandwich ELISA was performed using purified rat anti-human IL-12p70 (0.5 mg/ml) as capture antibody, followed by a biotin-conjugated mouse anti-human IL-12 (p40/p70) antibody (0.5 mg/ml) together with horseradish peroxidase-conjugated streptavidin; all the reagents were provided by BD Pharmingen. Colorimetric data were collected using a Multiscan ELISA reader (Laboratory Systems) after the addition of the chromogen 3-3′,5,5′-tetramethylbenzidine (Neogen); the concentration of cytokines in supernatants was calculated by referring to a standard curve using Ascent software (Thermolab Scientific). The sensitivity of both ELISA assays was 20 pg/ml.
Measurement of cytokines using multicytokine bead arrays
MoDC supernatants were analyzed for IL-1β, IL-6, TNF-α, IL-10, CXCL8, and CCL3 using FlowCytomix™ simplex kits (Bender MedSystems) following the manufacturer’s instructions. Briefly, a mixture of fluorescent polystyrol beads coupled with antibodies specific to each analyte was incubated with the samples. Concurrently, a biotin-conjugated antibody mixture was added that binds to the analytes captured by the first antibodies; then PE-streptavidin was added to allow fluorescent emission signals. Samples were analyzed by flow cytometry based on differential bead sizes and their distinct spectral signature; FlowCytomix Pro 2.3 software enabled calculation of the concentration of each cytokine using a standard curve generated for each analyte.
Statistical analysis
Statistical analyses were carried out using GraphPAD Prism version 4. One-way ANOVA was used to test for statistically significant differences between nonparametric datasets; the limit of significance was set at 0.05. Posthoc testing was done with Dunn’s test. Spearman’s rank correlation coefficient, ϱ, was used to test for the correlations between nonparametric measures.
RESULTS
IL-23 and IL-12p70 production in response to PRR ligands in HD and in patients with AS and RA
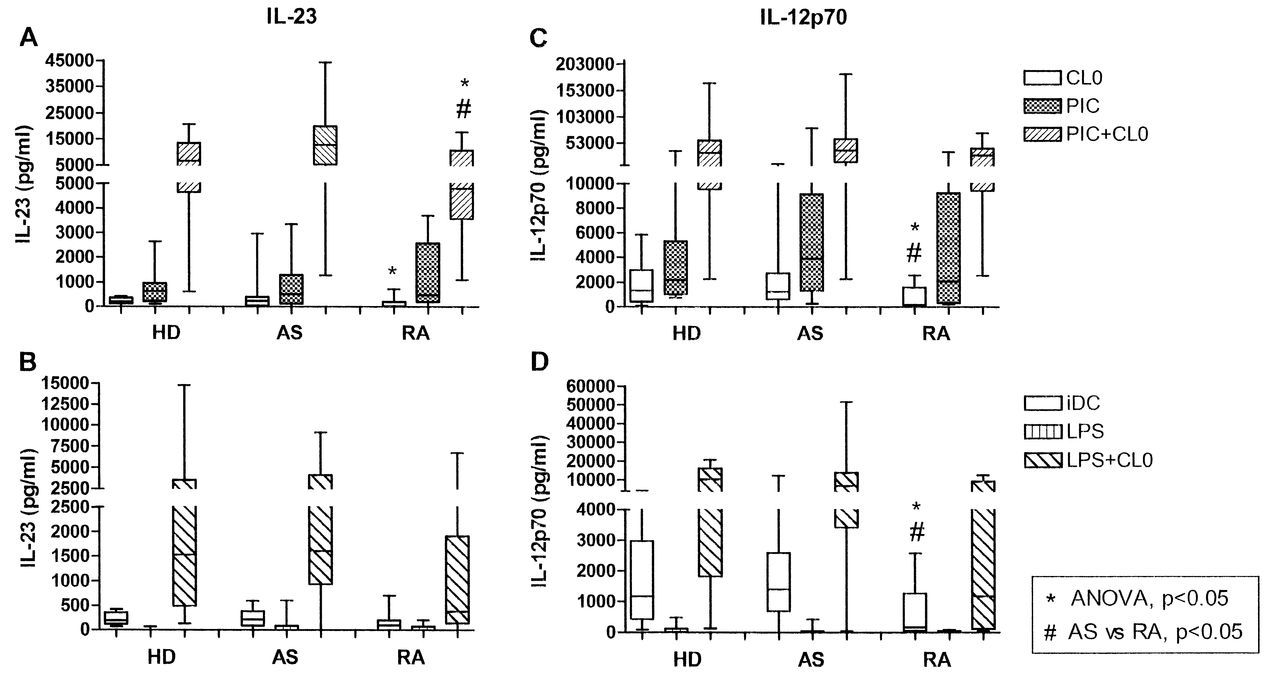
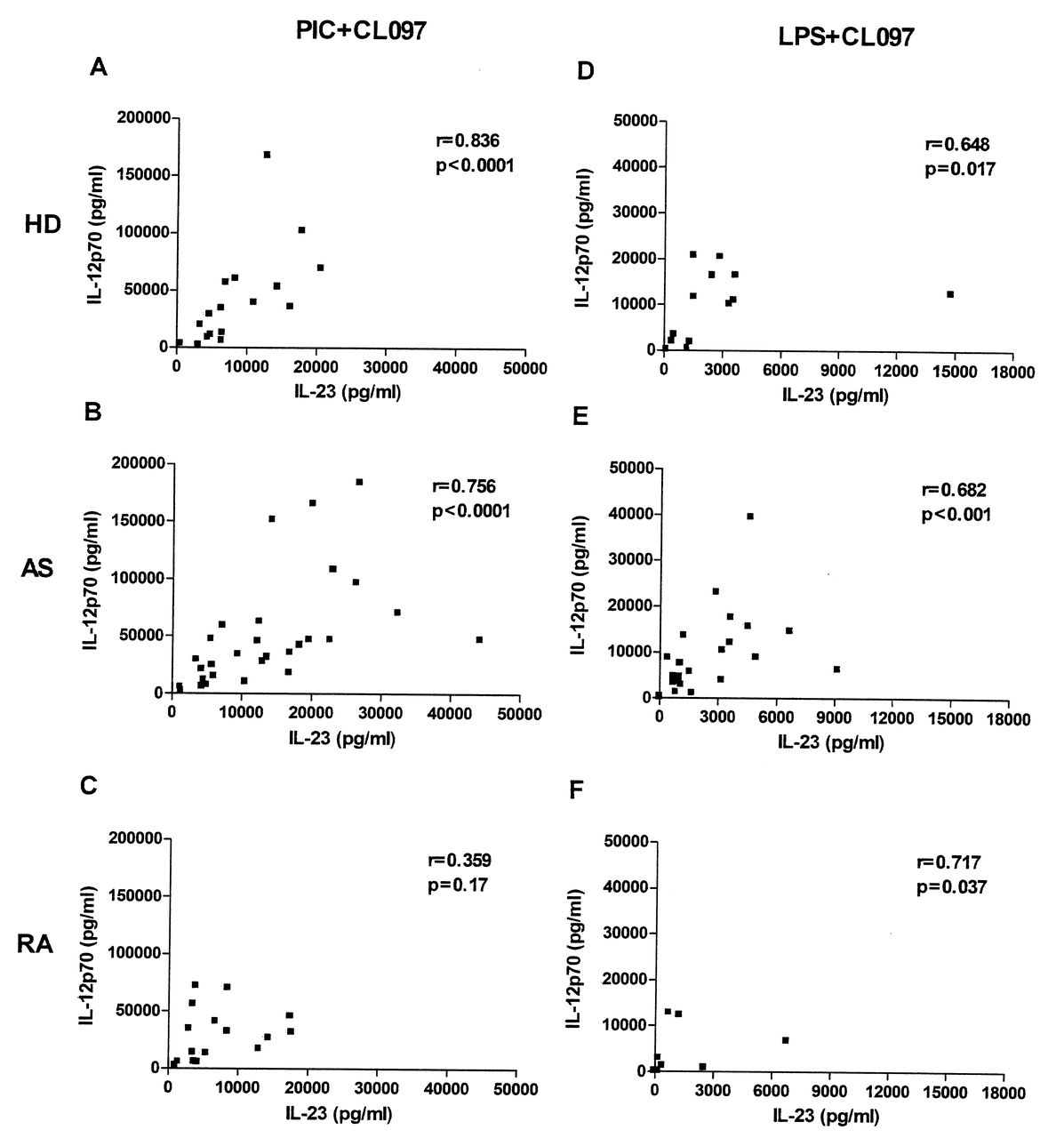
IL-23 and IL-12 secretion was measured following stimulation of moDC with ligands for TLR4, TLR3, and TLR7/8, alone or in combination. The optimal concentrations of ligands and the timepoints at which maximal secretion of the cytokines occurred were determined previously (data not shown). The levels of both cytokines were below the detection limit of ELISA assays in unstimulated (immature) moDC (data not shown), but moDC produced both IL-23 and IL-12p70 following stimulation with either TLR3 or TLR7/8 ligands, although at relatively low levels in all donors (Figure 1). Minimal amounts of IL-23 or IL-12p70 were produced upon stimulation of TLR4 with ultra-pure LPS. However, the combination of a TLR7/8 ligand with those for TLR3 or TLR4 led to marked synergistic production of both IL-23 and IL-12p70, not only in HD, but also in patients with AS and RA (Figure 1). Among the combinations we used, poly(I:C; TLR3 ligand) + CL097 (TLR7/8 ligand) was optimal. Moreover, we observed significant positive correlations between IL-23 and IL-12p70 secretion in moDC from HD and patients with AS, but not in patients with RA, using either poly(I:C) or LPS combined with CL097 (Figure 2).

IL-23 and IL-12p70 secretion by monocyte-derived dendritic cells (moDC) in response to various pattern recognition receptor ligands. IL-23 (panels A, B) and IL-12p70 (panels C, D) produced by moDC from healthy donors (HD) and patients with AS and RA in response to CL097 and poly(I:C) (PIC), singly or in combination (A, C); and to CL097 and LPS, singly or in combination (B, D). Data represent results from 17 HD, 30 patients with AS and 16 with RA (CL097, PIC, PIC+CL097) or 14 HD, 24 patients with AS and 9 with RA (LPS, LPS+CL097). Dashes represent median values. *p < 0.05 by ANOVA, #p < 0.05 AS vs RA posthoc analysis.
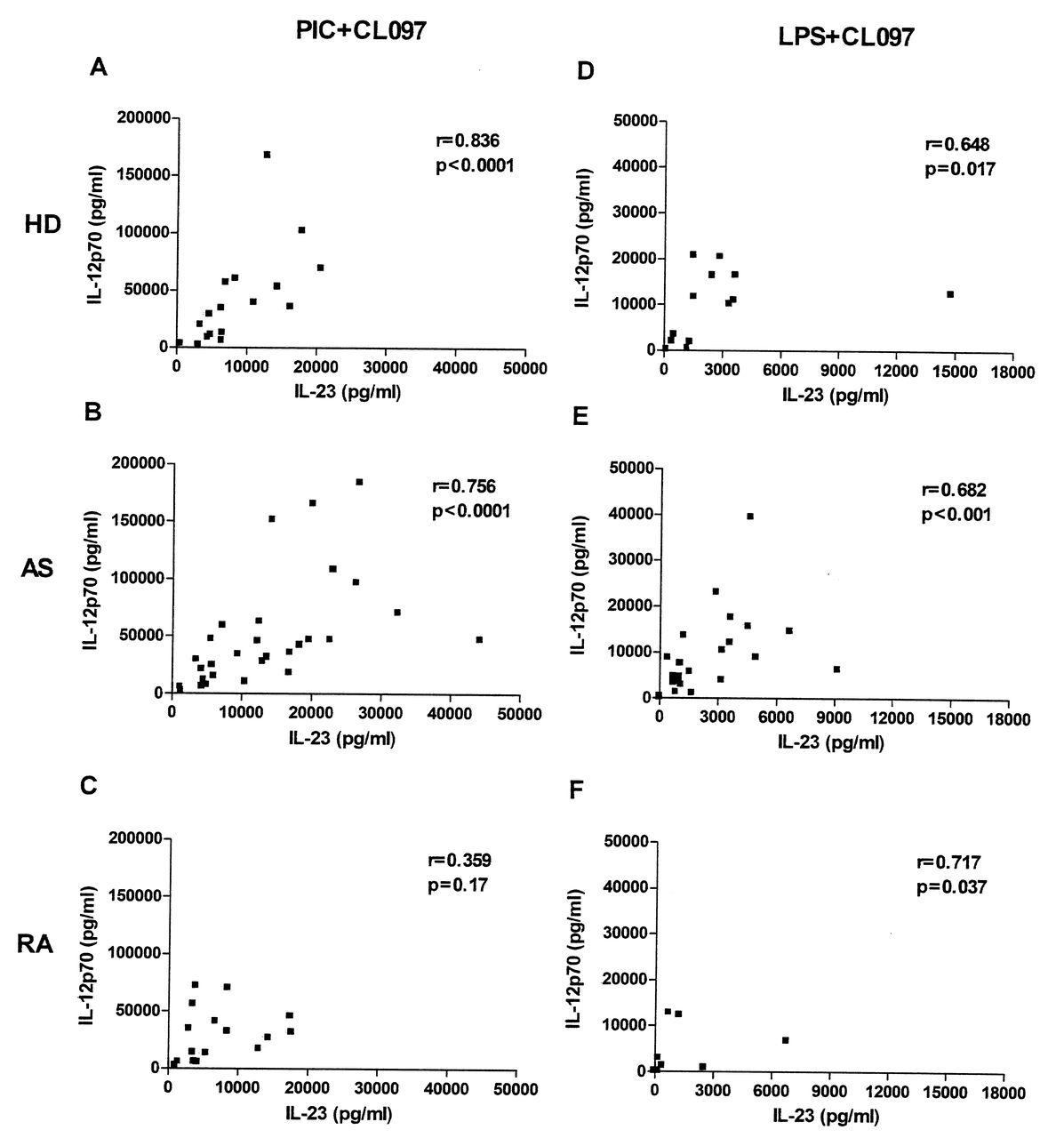
Correlations between IL-23 and IL-12p70 secretion in healthy donors (HD) and patients with AS and RA. Concentrations of IL-23 (X-axis) and IL-12p70 (Y-axis) obtained from monocyte-derived dendritic cells in response to a combination of CL097 and poly(I:C) (PIC+CL097, panels A, B, C), or CL097 and LPS (LPS+CL097, panels D, E, F) from HD (A, D) and patients with AS (B, E) and RA (C, F). The numbers of donors in each group are as in Figure 1. Spearman’s rank correlation coefficient rho (r) and corresponding p values are shown.
When the most potent stimulus, i.e., CL097 + poly(I:C), was tested, analysis of the 3 groups by ANOVA showed a significant difference among the groups (p = 0.028). Posthoc testing showed that moDC from patients with AS secreted significantly more IL-23 than did moDC of patients with RA (p < 0.05). While patients with AS showed a 2-fold increase in the median IL-23 production compared with HD, this difference did not reach statistical significance (Figure 1A). In contrast, there were no significant differences in IL-12p70 secretion among the 3 groups when the optimal combination was used (Figure 1C). The relatively low levels of IL-12 obtained using CL097 only were, however, significantly different between patients with AS and patients with RA (p ≤ 0.05). When stimulation with ultra-pure LPS was combined with the CL097, much lower levels of IL-23 and IL-12p70 were seen (Figure 1B and 1D), and there were no statistically significant differences among groups, although a trend was still evident toward decreased levels of IL-23 secretion by moDC from patients with RA, as compared to moDC from patients with AS.
The difference in IL-23 secretion in RA compared to patients with AS prompted us to challenge moDC with peptidoglycan (a ligand for TLR2/6) and curdlan, a dectin-1 ligand. IL-23 secretion was pronounced upon stimulation with curdlan, while peptidoglycan induced only low levels in moDC from all subjects (Figure 3). However, Pam3CSK4, a specific ligand for TLR2 paired with TLR1 rather than TLR6, enhanced IL-23 production in response to curdlan in all subjects, and ANOVA revealed a significant difference among the groups (p = 0.033). Although this difference again appeared to be due to the lower production by the moDC of patients with RA, this did not reach significance in posthoc testing (Figure 3). Peptidoglycan and curdlan, or curdlan in combination with Pam3CSK4, resulted in very low levels of IL-12p70 secretion in all subjects (data not shown).

IL-23 secretion by monocyte-derived dendritic cells (moDC) in response to peptidoglycan and curdlan. IL-23 produced by moDC from healthy donors (HD) and patients with AS and RA in response to peptidoglycan (PGN) and curdlan singly, or curdlan and Pam3CSK4 (PAM3K) in combination. Data represent results from 17 HD and 30 patients with AS and 16 with RA (PGN and curdlan) or 12 HD and 18 AS and 10 RA (curdlan+Pam3CSK4). Dashes represent median values. *p < 0.05 by ANOVA.
We tested whether the ability to produce IL-23 and IL-12 correlated with clinical measurements in AS or RA. When using an optimal stimulus (poly I:C + CL097), we noted a significant correlation between the Bath Ankylosing Spondylitis Disease Activity Index (BASDAI) and IL-23 production (r = 0.39, p < 0.05), but this was not found for IL-12p70 production. Similarly, when patients with AS were divided into those with low and high BASDAI, those with active disease showed higher levels of IL-23 (p = 0.027). No correlations with age or disease duration were seen in AS. In RA, age, disease duration, C-reactive protein (CRP), erythrocyte sedimentation rate (ESR), swollen joint counts, and presence of erosions were also not correlated with IL-23 or IL-12p70 production, and showed no differences when divided into those with high and low ESR or CRP. CRP and ESR were not measured routinely in the AS cohort because they are known to correlate poorly with disease activity as measured by clinical indices. Interestingly, treatment of AS with TNF-α-blocking drugs had no consistent effect on the ability of moDC to produce IL-23 and IL-12p70, because comparable levels were produced by patients with AS treated with nonsteroidal antiinflammatory drugs (NSAID) alone and those receiving TNF-α blockade (either etanercept or adalimumab). This may reflect our use of moDC derived from PB monocytes by ex vivo culture for 6 days before testing their cytokine secretion, so that effects of drug treatment or the inflammatory environment may no longer be evident.
Expression of IL-23p19, IL-12p35, and IL-12/IL-23p40 mRNA in response to PRR ligands
The findings of dissimilar secretion of IL-23 between patients with RA and those with AS raised the question of whether this reflected altered gene transcription. Following preliminary experiments to establish the time course of expression of mRNA for IL-23 and IL-12 subunits in response to TLR agonists, cells were harvested at 6 h poststimulation. At this time any differences in mRNA expression between different donors were already established and were consistent throughout the rest of the time course (data not shown). Increases in mRNA for each subunit of IL-23 and IL-12 in response to poly(I:C) + CL097 were comparable in all groups, with a small number of outliers in each group, and no evidence of impaired transcription in RA moDC (Figure 4). Similarly, the increase in IL-23p19 RNA in response to curdlan did not differ in limited numbers of patients with RA, AS, and HD tested (n = 2, 6, and 4, respectively; data not shown). These results suggest that IL-23 secretion may be regulated posttranscriptionally, and that the difference between patients with RA and those with AS occurs at this level.

Expression of mRNA for IL-23 and IL-12 subunits in monocyte-derived dendritic cells (moDC) treated with TLR3 and TLR7/8 ligands. Upregulation of mRNA levels for IL-23p19 (panel A), IL-12/IL-23p40 (panel B), and IL-12p35 (panel C) in response to the combination of poly(I:C) and CL097 by moDC from healthy donors (HD; n = 11) and patients with AS (n = 15) and RA (n = 10). Data are reported as fold change. Dashes represent median values.
MoDC activation in response to PRR ligands
To assess whether differences in IL-23 production reflected intrinsic defects in their response to PRR ligands, the effect of TLR agonists on surface expression of CD86, HLA-DR, and CD83 at 24 h poststimulation was measured by flow cytometry. Increased levels of expression for each of these markers were similar in moDC from each group (Figure 5), indicating a similar intrinsic capability of all cells to respond to activation. Similarly, the phenotype of unstimulated moDC, CD14−CD11c+CD1c+, was comparable in HD and patients with AS and RA (data not shown). Moreover, while different TLR ligands varied in their ability to induce expression of particular markers, all subjects showed an identical pattern of response to each of the ligands tested (Figure 6) and also to curdlan (data not shown).
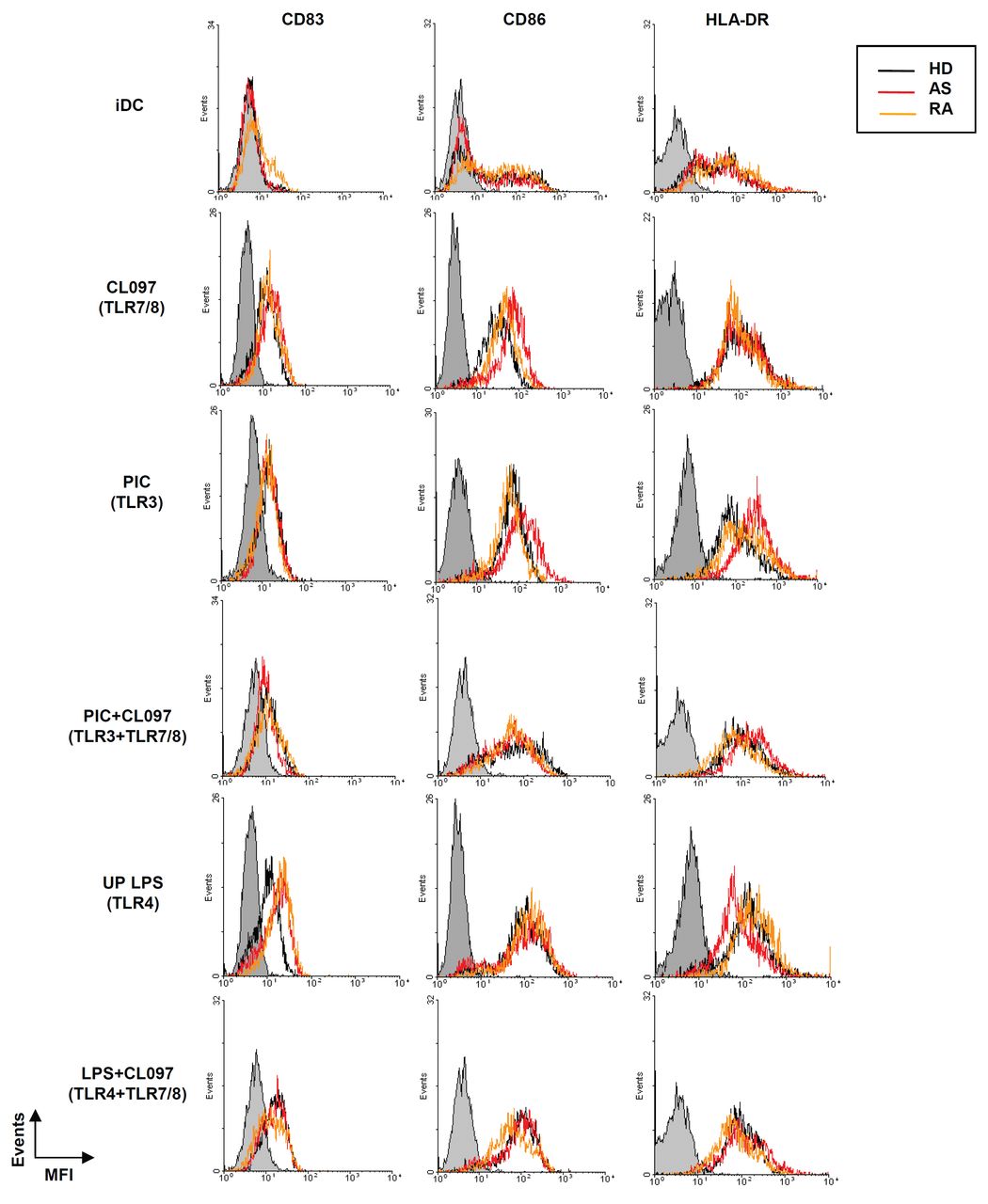
Expression of activation markers on monocyte-derived dendritic cells (moDC) from healthy donors (HD) and patients with AS and RA. Expression of CD83, CD86, and HLA-DR by moDC untreated (iDC) or stimulated with CL097, poly(I:C) (PIC), or LPS alone, or with CL097 in combination with either poly(I:C) (PIC+CL097) or LPS (LPS+CL097). Mean fluorescence intensity (MFI) for each marker is plotted. Data from a single representative donor from each group are shown. Gray shaded lines indicate staining with fluorochrome-conjugated isotype control antibodies.

Analysis of activation marker expression in monocyte-derived dendritic cells (moDC) stimulated with TLR ligands. A. Percentage of moDC positive for both CD83 and CD86 either untreated (iDC) or treated with CL097, poly(I:C; PIC), or LPS, or a combination of CL097 with poly(I:C) (PIC+CL097) or LPS (LPS+CL097) from healthy donors (HD) and patients with AS and RA, using the same conditions as in Figure 5. Panels B-D. Mean fluorescence intensity of CD86 (B), CD83 (C), and HLA-DR (D), represented as fold increase (ratio between treated and unstimulated moDC from each donor). Data represent median ± SEM. Data were obtained from at least 11 HD, 15 patients with AS, and 10 with RA (iDC, PIC, CL097, PIC+CL097) or 7 HD, 12 with AS, and 6 with RA (LPS, LPS+CL097).
Production of additional cytokines and chemokines in response to PRR ligands
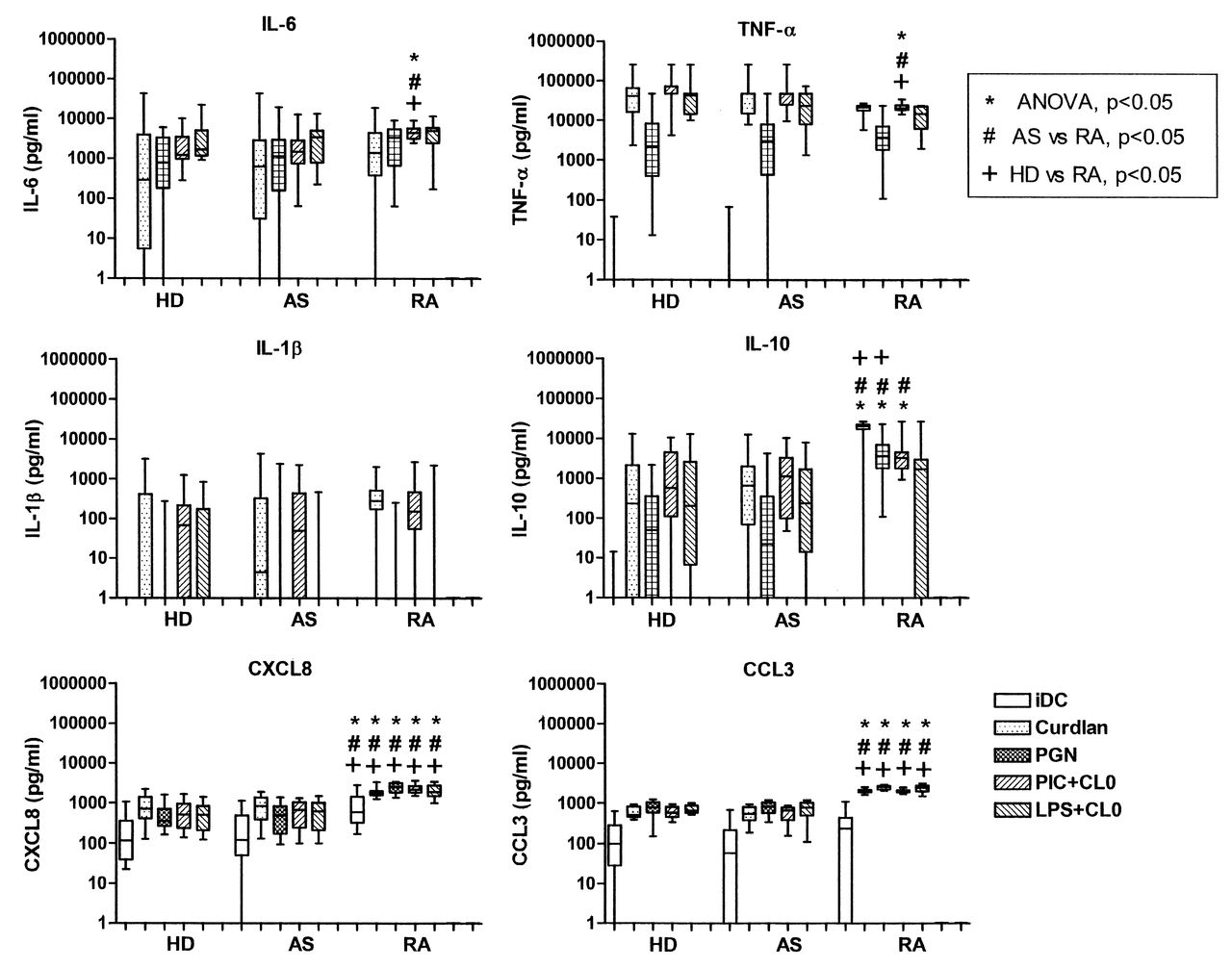
Having observed differences in secretion of IL-23 in moDC from patients with RA, we widened our investigation of the response to PRR ligands by measuring a range of cytokines and chemokines using multiplex bead assays. Levels of IL-6, TNF-α, and IL-10 in response to different PRR ligands, individually or in combination, were comparable in all subjects (Figure 7). Interestingly, moDC from patients with RA produced significantly higher levels of both CXCL8 and CCL3 than moDC from HD or from patients with AS, in marked contrast to their decreased production of IL-23.
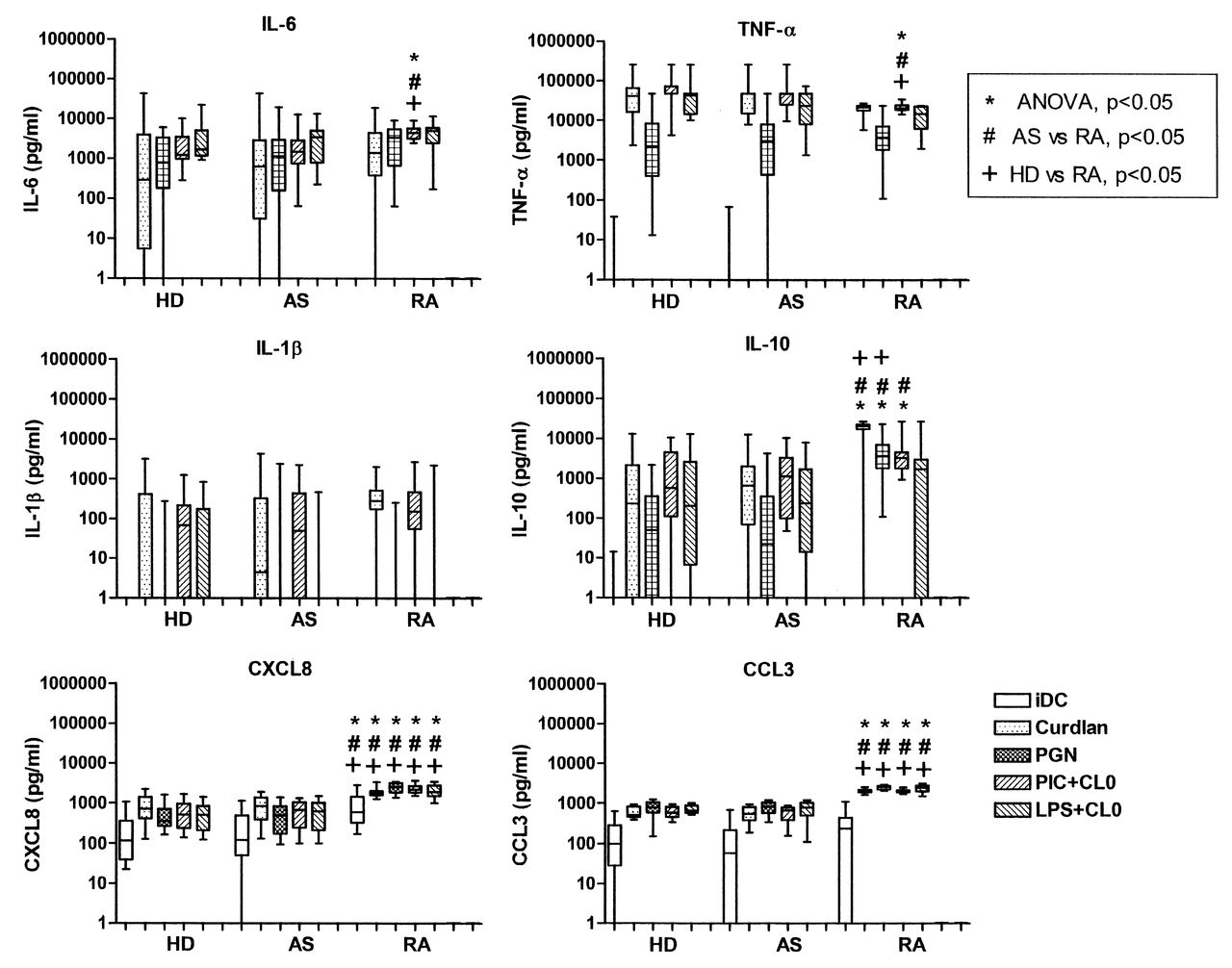
Secretion of cytokines and chemokines by monocyte-derived dendritic cells (moDC) from 16 healthy donors (HD), 28 patients with AS, and 16 with RA. Box and whisker plots for cytokine and chemokine secretion by moDC, untreated (iDC) or stimulated with curdlan, peptidoglycan (PGN), or combinations of CL097 with poly(I:C) (PIC+CL097) or LPS (LPS+CL097). P values are from posthoc analysis.
DISCUSSION
Cytokines secreted by DC have a fundamental role in inducing and sustaining autoimmune and chronic inflammatory diseases; in particular, the balance between IL-23, which sustains the survival of Th17 cells, and IL-12, which induces Th1 cell development, may be a critical factor8,9,20,38. In view of the increased numbers of IL-17-producing T cells demonstrated in both AS and RA, we hypothesized that this might be driven by IL-23, and therefore that IL-23 production might be increased in patients with AS and/or RA as compared to HD. However, when we analyzed IL-23 production by moDC, those from patients with RA produced significantly lower amounts of IL-23 in response to several combinations of TLR ligands than moDC of patients with AS. In contrast, secretion of the related cytokine, IL-12, was comparable in both forms of arthritis. Further, there was no global deficiency in the responsiveness of RA moDC, because they produced higher amounts of CXCL8 and CCL3 than those from HD or patients with AS, and their upregulation of surface expression of CD86, CD83, and HLA-DR was also unchanged. While expression of TLR has been shown to vary in different conditions, these data exclude levels of receptor expression as the explanation for the differences seen in IL-23 secretion, unless IL-23 production was to be much more sensitive to TLR expression levels than other TLR-mediated effects, and there is no evidence that this is the case. The differences in IL-23 secretion were not mirrored by mRNA levels, suggesting that the differences relate to posttranscriptional regulation of IL-23 synthesis. A recent report has shown that IL-23 mRNA stability is controlled posttranslationally in a way that is similar to TNF-α mRNA, involving an AU-rich region in the 3′ untranslated region, and the action of tristetraprolin39.
The combination of TLR7/8 and TLR3 ligands induced synergistic secretion of both IL-23 and IL-12p70 from moDC of all donors, in agreement with studies on IL-12p70 production40,41,42. Synergistic effects on IL-23p19 transcription have also been reported in HD40,42, but in contrast to our clear results, others did not report any synergistic effect on IL-23 secretion42. The reason for this difference is unknown, but could be due to the different imidazoquinoline derivatives used as TLR7/8 ligands, i.e., R-848 by Makela, et al42 and CL097 in our study.
Beta-glucans, including curdlan, engage the PRR, dectin-1, and have been shown to be potent inducers of IL-23 secretion and hence able to drive a robust Th17 response, which is needed to combat fungal infection43. Recently, Plantinga and colleagues showed high levels of expression of dectin-1 in RA synovial tissues, although its contribution to synovial inflammation still remains unclear44. Again, in our study, IL-23 secretion induced by stimulation with curdlan in combination with the TLR2 ligand, Pam3CSK4, resulted in lower levels of IL-23 from RA moDC as compared to those from patients with AS, thereby supporting our findings with the combination of TLR3 and TLR7/8 agonists.
Although responses to single TLR ligands were much lower than responses to combinations, production of IL-12p70 in response to the TLR7/8 ligand CL097 was significantly lower in moDC from patients with RA. This raises the possibility of a defect in TLR7/8-mediated responses in RA, but impairments in signaling by TLR7 and/or TLR8, which are located in endosomes rather than the cell surface, cannot solely account for the differential secretion of IL-23 in RA, because stimulation through TLR2 + dectin-1 also resulted in decreased secretion of IL-23 by moDC from patients with RA.
The precise roles played by IL-23 in AS and RA are still unclear. While IL-23 has been implicated in animal models of both AS and RA, there are no associations between genes involved in response to IL-23 and the generation of Th17 cells in RA. This allows the hypothesis that this cytokine pathway may be of primary importance in AS pathogenesis, as compared to acting as a downstream effector mechanism in RA. Very low levels of IL-23 have been reported in the sera of patients with SpA and RA, and these did not differ from the levels in HD sera29,30,45. It is likely, therefore, that IL-23 may act locally rather than systemically46, although a correlation between serum levels of IL-23 and disease activity measures has been reported in RA, but not SpA45. Low amounts of biologically active IL-23 have been detected in the synovial fluid of both patients with SpA and those with RA29,30,31,45, but the main influence of IL-23 is likely to be on cell:cell interactions within synovial tissue. In this study secretion of neither IL-23 nor IL-12p70 was generally correlated with clinical measures in AS or RA, with the exception of a correlation between IL-23 secretion and BASDAI in AS. While treatment with TNF blockade could affect moDC cytokine secretion, as reported47, this had no discernible effect in our study as far as AS was concerned, because cytokine production in 7 patients with AS treated with TNF blockade (etanercept or adalimumab) did not differ from those treated with NSAID alone (only 1 patient with RA was receiving TNF-blocking agents). Formally excluding all donors receiving anti-TNF drugs from our analysis did not affect the statistically significant differences.
Our findings suggest that moDC derived from patients with RA and AS differ with regard to their ability to produce IL-23. There is a tendency in moDC from patients with RA to make lesser amounts of IL-23, while not altering their production of IL-12 and consequently altering the ratio of these 2 cytokines and their propensity to develop Th1 versus Th17 cells. In contrast, moDC from patients with AS maintain levels of secretion of both IL-23 and IL-12 comparable to those observed in HD, despite having levels of systemic inflammation similar to those with RA. These observations can be compared with the recent report from Zeng, et al48 showing clearly increased IL-23 secretion from macrophages of patients with AS. While our study did not show significantly increased IL-23 production by AS DC, its maintenance in active inflammation (in contrast to RA) may point to a favoring of IL-23 secretion in DC also. Together these observations are consistent with a prominent role for IL-23 in AS pathogenesis, as recently hypothesized.
Acknowledgment
We thank Dominique Raut-Roy for patient recruitment and documentation, and Dr. Robert Busch for helpful discussion.
Footnotes
-
Supported by Arthritis Research UK and by the National Institute for Health Research Cambridge Biomedical Research Centre.
- Accepted for publication June 27, 2012.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}