

Abstract
Objective. To compare levels of the chemokine CCL20 and its receptor CCR6 in donor, osteoarthritic (OA), and rheumatoid arthritis (RA) synovium; and to determine the molecular mechanism of cellular activation induced by chemokine/receptor ligation in human fibroblast-like synoviocytes (FLS).
Methods. Synovia and isolated FLS from donor, OA, and RA joints were analyzed for CCL20 and CCR6 expression by RT-PCR and immunohistochemistry. The effect of CCL20 on cytokines and mediators of cartilage degradation was examined by PCR for mRNA expression levels and ELISA, and Western blotting for protein. CCL20-dependent transcriptional and posttranscriptional activation of target genes was monitored using reporter constructs and luciferase assays in transfected donor FLS.
Results. CCL20 and CCR6 proteins were abundantly expressed in RA synovial lining cells compared to donor or OA synovia as judged by immunohistochemistry. RT-PCR of synovial extracts confirmed the predominance of CCL20/CCR6 mRNA expression in RA synovium. CCL20 mRNA expression was low in donor FLS, but increased dramatically after stimulation with recombinant human (rh) interleukin 1ß (IL-1ß). rhCCL20 increased mRNA and protein expression of COX-2, IL-1ß, tumor necrosis factor-α, IL-6, and the matrix-destructive metalloprotease MMP-3 in donor FLS cultures. High constitutive levels of IL-6 were released from RA synovia; CCL20-induced expression of IL-6 occurred through an NSAID/COXIB-sensitive process. CCL20-induced expression of COX-2 was mediated by a PLCP1/PKCα/MEK1/2/ERK1/2-dependent pathway involving both transcriptional and posttranscriptional mechanisms.
Conclusion. CCL20/CCR6 may play an important role in the pathogenesis of RA by assembling the molecular and cellular components orchestrating synovitis.
Rheumatoid arthritis (RA), the most common of inflammatory arthritides, is a chronic systemic autoimmune inflammatory disorder with characteristic joint involvement, the hallmarks of which are erosive synovitis with pannus formation, bone lesioning, cartilage degradation, and joint failure1,2. The disease is associated with a dramatic increase in the number of fibroblast-like synovial cells (FLS) in the lining layer and infiltrating inflammatory cells, including T and B lymphocytes, macrophages, and mast cells in the sublining space3. The latter cell phenotypes produce cytokines, chemokines, and other inflammatory mediators that drive the chronic inflammatory process, while the FLS contribute to inflammation and joint pathology by releasing matrix-destructive metalloproteases, chemoattractants, cytokines, growth promoters, neoangiogenic factors, and small bioactive lipids; it is unknown, however, how the ambient cell phenotypes coordinate and interact to orchestrate synovitis3,4.
Chemokines (e.g., CCL2, CCL3, CCL5, CCL13, CCL18, and CCL20) represent a family of cytokines that are primarily known to induce adhesion and directional migration of specific cell types to extravascular sites of inflammation5. The implication of CCL20, for example, in the pathogenesis of RA has been reported, particularly with regard to specific recruitment of circulating CD8+ CTL, Th17 cells, B cells, and dendritic cells (DC) to joint tissues, regulating local immune responses and possibly accounting for the chronicity of the disease6,7. Indeed, CCL20 was shown to be expressed in FLS upon activation by proinflammatory cytokines [interleukin 1ß (IL-1ß), IL-6, tumor necrosis factor-α (TNF-α)] and can also be detected in RA synovial fluid6,7,8,9. The chemokine, also known as macrophage inflammatory protein a (MIP-3α), liver activation regulated chemokine (LARC), or EXODUS, binds to a single septahelical G-protein-coupled receptor, CCR, the latter being expressed abundantly on activated cells of myeloid, epithelial, and mesenchymal lineages (reviewed in Schutyser, et al10). Given the contribution of FLS and the putative role of CCL20 in RA joint pathology11, we hypothesized that CCL20 may serve as a critical factor assembling the molecular and cellular components responsible for synovitis. The objective of our study was to confirm the concomitant expression of CCL20 and CCR6 in RA-affected synovium/FLS, in comparison to noninflammatory donor and osteoarthritic (OA) tissues, and to examine the molecular signaling mechanisms involved in CCL20/CCR6-triggered proinflammatory cytokine/mediator release.
MATERIALS AND METHODS
Isolation and culture of fibroblast-like synoviocytes
Synovial lining cells (fibroblast-like synoviocytes) were isolated from synovial membranes obtained at necropsy from 8 donors (mean age 30 ± 7 yrs) with no history of arthritic disease and no inflammation. Additional experiments were conducted (where specifically indicated) with FLS specimens obtained from 19 OA and 7 RA patients (mean age 67 ± 19 yrs) undergoing arthroplasty who were diagnosed based on the criteria developed by the American College of Rheumatology Diagnostic Subcommittee for OA/RA12,13. McGill University Ethics Review Board and local ethics committees at the Jewish General Hospital and Royal Victoria Hospital (Montreal, Canada) approved the use of specimens obtained surgically or by necropsy. Next of kin provided informed consent for samples obtained by necropsy and OA and RA patients were asked to approve/sign consent forms prior to surgery. The 7 RA patients, with duration of disease of 11.9 ± 2.8 years, were treated with varying algorithms that included anti-TNF-α antibody (infliximab 3 to 5 mg/kg) for 9.3 ± 2.7 years, corticosteroids (prednisone 2 to 10 mg/day) for 9.6 ± 1.9 years, and methotrexate (10 to 20 mg/wk) for 3.5 ± 1.6 years.
Human FLS were released by sequential enzymatic digestion with 1 mg/ml pronase (Boehringer Mannheim, Laval, QC, Canada) for 1 h, followed by 6 h with 2 mg/ml collagenase (type IA; Sigma-Aldrich) at 37°C in Dulbecco modified Eagle’s medium (DMEM) supplemented with 10% heat inactivated fetal bovine serum (FBS), 100 units/ml penicillin, and 100 μg/ml streptomycin14,15. Released synovial fibroblasts were incubated 1 h at 37°C in tissue culture flasks (Primaria 3824; Falcon, Lincoln Park, NJ, USA), allowing the adherence of nonfibroblastic cells possibly present in the synovial preparation, particularly from OA and RA synovia. In addition, flow cytometric analysis (Epic II, Coulter, Miami, FL, USA) using the anti-CD14 (fluorescein isothiocyanate) antibody, was conducted to confirm that no monocytes/macrophages were present in the synovial fibroblast preparation14. The cells were seeded in tissue culture flasks, and cultured until confluence in DMEM supplemented with 10% FBS and antibiotics at 37°C in a humidified atmosphere of 5% CO2/95% air. The cells were incubated in fresh medium containing 0.5%–1% FBS for 24 h before the experiments and only primary, first, or second passage FLS were used.
Immunohistochemistry
Synovial tissues were fixed with 4% paraformaldehyde and embedded in paraffin to cut 5 μm sections. The tissue sections were placed on slides, deparaffinized in toluene, and dehydrated in graded series of ethanol, then washed in PBS and in 0.2% peroxidase. The presence of CCL20 was determined using the avidin-peroxidase method with goat antibody to human CCL20 (R&D Systems, Minneapolis, MN, USA). This antibody does not cross-react with CCL20-2, CCL20-3 or any other known chemokine. The antibody was used at 5 μg/ml and incubated with the sections for 1 h at room temperature. Biotinylated goat anti-rabbit was used as a secondary antibody. The presence of CCR6 was also determined as described above using 2 μg/ml mouse antibody to human CCR6 (R&D Systems). This antibody does not cross-react with CCR1, CCR2, CCR3, CCR4, CCR5, CCR7, CXCR1, CXCR2, CXCR3, CXCR4, and STRL33 transfectants. Biotinylated goat anti-mouse was used as secondary antibody. Negative controls were performed by replacing the primary antibody with species and isotype-matched immunoglobulin.
Semiquantitative RT-polymerase chain reaction
FLS (500,000 cells per well in 6-well plates) were stimulated with cytokines for 4 h. Total RNA was isolated by a single-step guanidinium thiocyanate-phenol-chloroform method. Cells were lysed directly in the flasks using RNA Stat-60 (Tel-Test B Inc., Friendswood, TX, USA). cDNA was prepared from RNA using random hexamers and SuperscriptII (Life Technologies, Gaithersburg, MD, USA). PCR was performed with the following primers, sense/antisense; CCL20: 5′-ACT GAG GAG ACG CAC AAT ATA T-3′ and 5′-TGT ACC AAG AGT TTG CTC CTG G-3′; COX-2: 5′-TTG TCC CAG ACA AGC AGG C-3′ and 5′-CAT TCC TAC CAC CAG CAA CC-3′; IL-1ß: 5′-GAG CTC GCC AGT GAA ATG ATG GC-3′ and 5′-CAA GCT TTT TTG CTG TGA GTC CCG-3′; IL-6: 5′-CAC AGA CAG CCA CTC ACC TCT TC-3′ and 5′-GCT GCG CAG AAT GAG ATG AGT TGT-3′; GAPDH: 5′-TGG TAT CGT GGA AGG ACT CAT GAC-3′ and 5′-ATG CCA GTG AGC TTC CCG TTC AGC-3′.
The PCR conditions for CCL20 were 4 min at 94°C, threshold cycling of 30 s at 94°C, 30 s at 60°C, and 30 s at 72°C. The same conditions were applied for IL-6, COX-2, and IL-1ß mRNA measurements. The conditions for GAPDH were 4 min at 94°C, cycle-matching of 30 s at 94°C, 30 s at 60°C, and 30 s at 72°C. Threshold cycling refers to performing increasing numbers of elongation/polymerization cycles to establish a linear range for product measurement. PCR products were separated on 1.5% agarose gels and banding was quantified with a densitometry digital imaging system (Alpha G-Imager 2000; Canberra Packard, Mississauga, ON, Canada) for semiquantitative measurements; changes in product expression were always considered as a ratio, product/GAPDH mRNA.
IL-6, MMP-3, and prostaglandin E2 assays
Half-area ELISA plates (Costar, Cambridge, MA, USA) were coated with 25 ml of 2 mg/ml monoclonal mouse anti-human IL-6 antibody (R&D Systems) for 4 h and blocked with 3% bovine serum albumin. Twenty-five milliliters of sample or human IL-6 standard were added to the plate and incubated 2 h, followed by 1 mg/ml rabbit anti-human IL-6 (R&D Systems). Biotin-labeled goat anti-rabbit immunoglobulin (Sigma, St. Louis, MO, USA) was then added (1:2000 dilution). Bound IL-6 was detected with 0.16 ng/ml poly-horseradish peroxidase-labeled streptavidin (Accurate Chemical and Scientific, Westbury, NY, USA) and TMB plus hydrogen peroxide substrate (Kirkegaard & Perry, Gaithersburg, MD, USA). Sensitivity of the assay was 1–1200 pg. MMP-3 and prostaglandin E2 (PGE2) were determined by ELISA according to the manufacturer’s instructions (R&D Systems; Cayman Chemical, Ann Arbor, MI, USA, respectively).
Cytokines
Recombinant human IL-1ß was purchased from Peprotech Inc. (Rocky Hill, NY, USA). The ED50 of recombinant human IL-1ß determined by stimulation of 3H-thymidine incorporation by murine C3H/HeJ thymocytes was 0.1 ng/ml. Endotoxin level in the IL-1ß preparation was < 0.1 ng/mg cytokine. CCL20 (R&D Systems) ED50 (0.3 nM) endotoxin content was < 0.1 ng/ml. Recombinant human IL-17 and recombinant human TNF-α were purchased from R&D Systems. In this study IL-1ß was used at 2 ng/ml and CCL20 at 0.3–12 nM, thus the endotoxin contamination in cultures for IL-1 and CCL20 was less then 0.1 pg/ml. Using lipopolysaccharide from Salmonella minnessota, these levels of endotoxin do not increase IL-6 levels in FLS.
Preparation of cell extracts and Western blotting
Fifty to 100 μg of cellular protein extracted in RIPA buffer (50 mM Tris-HCl, pH 7.4, 150 mM NaCl, 2 mM EDTA, 1 mM PMSF, 10 μg/ml each of aprotinin, leupeptin and pepstatin, 1% NP-40, 1 mM sodium orthovanadate, and 1 mM NaF) or hot SDS-PAGE loading buffer, from control and treated cells, were subjected to SDS-PAGE through 10% gels (16 × 20 cm, final concentration of acrylamide) under reducing conditions, and transferred onto nitrocellulose membranes (GE Healthcare/Amersham Pharmacia Biotech, Piscataway, NJ, USA). After blocking with 5% BLOTTO for 2 h at room temperature and washing, the membranes were incubated overnight at 4°C with polyclonal anti-COX-1 or anti-COX-2 (1:1000 dilution; Cayman Chemical) in TTBS containing 0.25% BLOTTO. The second anti-rabbit antibody-HRP conjugate (1:10,000 dilution; Cell Signaling Technologies, Danvers, MA, USA) was subsequently incubated with membranes for 1 h at room temperature, washed extensively for 30–40 min with TTBS, and a final rinse with TTBS at room temperature. After incubation with an ECL chemiluminescence reagent (Amersham Pharmacia Biotech), membranes were prepared for autoradiography, exposed to Kodak X-Omat film (Kodak, Rochester, NY, USA), and subjected to digital imaging (Alpha G-Imager 2000; Canberra Packard) for semiquantitative measurements15. In addition to the anti-COX-1/2 anti-sera (Cayman Chemical), total and anti-phospho p44/42 (T202/Y204), anti-phospho MEK1 (S217/221), and anti-PKCa/ß II (T638/641) were purchased from Cell Signaling Ltd. (Waverly, MA, USA).
Plasmids and transfection experiments
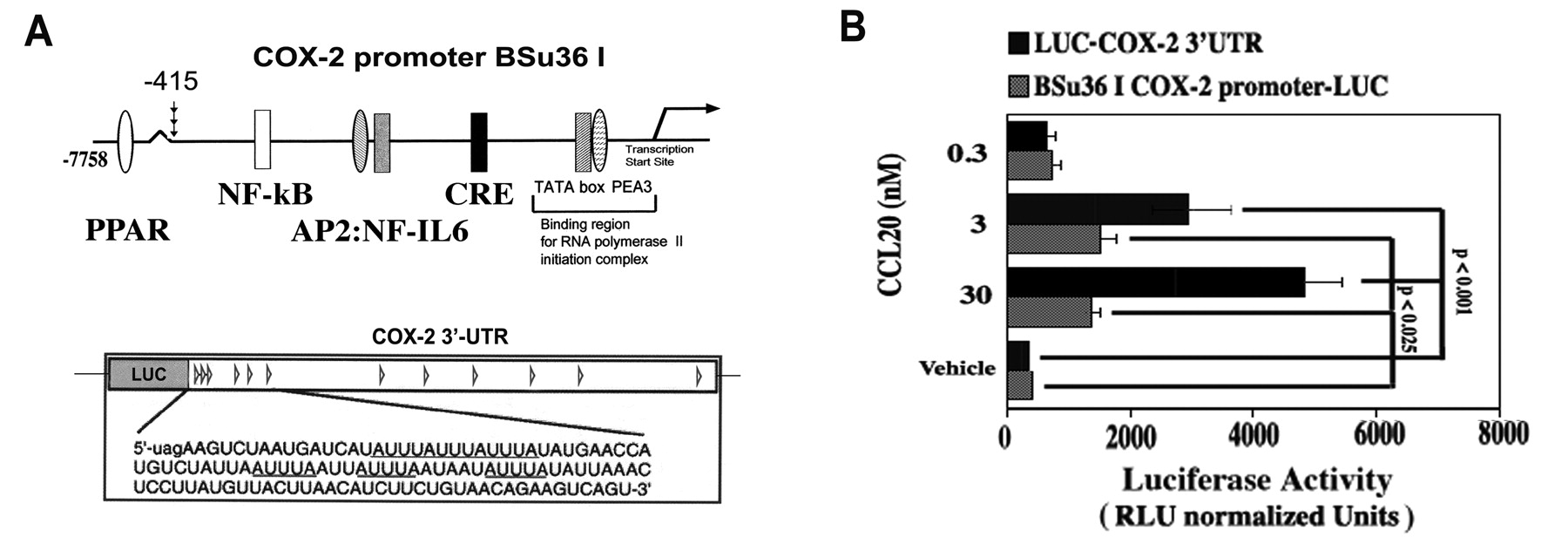
Transient transfection experiments were conducted in 12-well cluster plates as described15. Transfections were conducted using FuGENE6™ (Roche Applied Science, Indianapolis, IN, USA) for 6 h according to the manufacturer’s protocols with cells at 30%–40% confluence. Cells were reexposed to a culture medium with 1% FBS for 2 h prior to addition of the biological effectors. Transfection efficiencies were controlled in all experiments by cotransfection with 0.2–0.5 μg pCMV-ß-gal, a ß-galactosidase reporter vector under the control of CMV promoter (Stratagene, La Jolla, CA, USA) or a pHSV-TK-driven Renilla luciferase construct (Promega Corp., Madison, WI, USA). A BSu36 I COX-2 promoter (−415 to + 34)-LUC chimera was kindly provided by Dr. S. Prescott, University of Utah. In addition, chimeric luciferase reporter plasmids fused with the human COX-2 mRNA 3′-UTR (1451 bp; 22 Shaw-Kamen sequences) and a construct completely devoid of the COX-2 3′-UTR but containing the SV40 poly A signal were also employed to study posttranscriptional regulation16.
Statistical analysis
All results are expressed as mean ± SEM and statistical treatment of the data was by nonparametric (Mann-Whitney) analysis. Significance was acknowledged when the probability that the null hypothesis was satisfied at < 5% (p < 0.05).
RESULTS
CCL20 expression in normal and arthritic synovium and in cultured FLS
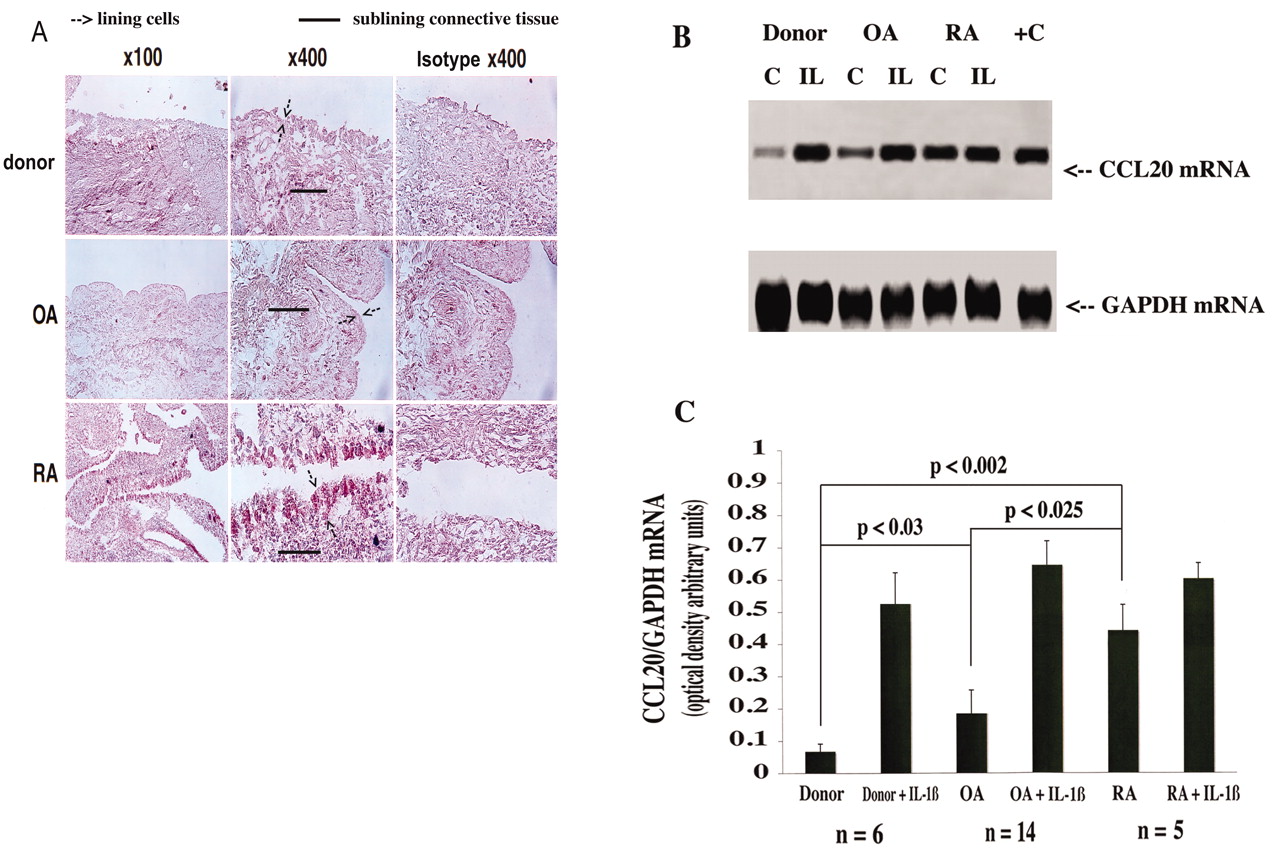
Preliminary immunohistochemical analysis demonstrated increased CCL20 expression in RA synovium (Figure 1A), primarily in the fibroblast population of the lining cell layer. Normal and OA human synovium processed under identical conditions expressed little or no CCL20 (Figure 1A). In order to substantiate and confirm these findings, we isolated freshly dispersed fibroblast populations from donor, OA, and RA-affected membranes that were depleted of blood-borne cells (see above) and measured CCL20 mRNA levels by semiquantitative cycle-controlled threshold RT-PCR. In donor FLS, CCL20 mRNA was either undetectable or present at low levels. Higher constitutive CCL20 levels were expressed particularly in RA (RA vs donor, 6.91 ± 1.56-fold higher, p < 0.002; RA vs OA, 2.71 ± 0.42-fold higher, p < 0.025) and, to a lesser extent, in OA FLS populations (OA vs donor, 3.07 ± 0.9-fold higher; Figure 1B, 1C). In order to determine and compare the response of donor, OA, and RA FLS to clinically relevant proinflammatory cytokines, we established cells in low-passage culture and treated them with the proinflammatory cytokines IL-1ß and TNF-α (data not shown); we observed increased CCL20 mRNA levels in all 3 types of FLS, although rhIL-1ß-dependent induction was relatively modest in RA-derived FLS (Figure 1B, 1C).

CCL20 protein and mRNA expression in synovium and FLS. Donor, OA, and RA synovium samples were analyzed by immunohistochemistry for CCL20 protein expression (A). Results are representative of 4 different donors for 4 separate experiments each for OA, RA, and donor samples. B. FLS derived from donor, OA, and RA synovial cultures were stimulated with or without IL-1ß (2 ng/ml), then RNA was analyzed by PCR using specific primers for CCL20 and GAPDH (representative study of 4 analyses). C. PCR products resolved on gels were subjected to densitometric and statistical analyses.
CCR6 expression in normal and arthritic synovium and in cultured FLS
Immunohistochemical analysis of CCR6 expression demonstrated specific staining in normal, OA, and RA synovium in the lining fibroblast layer and in blood vessels and was increased in arthritic tissues as compared to donor specimens (Figure 2A). We endeavored to confirm these findings in isolated FLS populations from donor, OA, and RA-affected membranes by measuring CCR6 mRNA levels by semiquantitative cycle-controlled threshold RT-PCR. CCR6 mRNA was detected in unstimulated donor, OA, and RA FLS cultures although the constitutive levels of CCR6 in RA samples were greater than in donor cells (RA vs donor, 2.47 ± 0.34-fold higher, p < 0.02; Figure 2B, 2C). rhIL-1ß but not TNF-α (data not shown) increased CCR6 mRNA expression in normal and arthritic FLS cultures (Figure 2B, 2C).

CCR6 protein and mRNA expression in synovium and FLS. Donor, OA, and RA synovium samples were analyzed by immunohistochemistry for CCR6 protein expression (A). Results are representative of 4 different donors for 4 separate experiments each for OA, RA, and normal synovium. B. FLS derived from donor, OA, and RA synovia were stimulated with or without IL-1ß (2 ng/ml), then RNA was analyzed by PCR using specific primers for CCR6 and GAPDH (representative study of 4 analyses). C. PCR products resolved on gels were subjected to densitometric and statistical analyses.
Regulation of FLS function by CCL20
Although CCL20 is considered a potent chemokine and chemoattractant, its abundance in RA-affected synovial lining cells suggested that it might serve as a critical factor assembling the cellular and also the molecular components orchestrating synovitis. In this connection, we examined specific CCL20-dependent signaling pathways and changes in the genetic program of target cells. In particular, we examined those substances (e.g., readouts like IL-6, IL-1, TNF-α, and MMP) with a strong association to the pathophysiology of RA1,2,3. In order to clearly define the responses, we chose donor FLS cultures, where presumably there was no background signal activation; we stimulated with CCL20 (0.3–30 nM, physiological range) and effects on gene expression were screened by threshold RT-PCR. The results showed that CCL20 markedly induced the expression of IL-6 and COX-2 (Figure 3A, 3B) at low relative concentrations (8.62 ± 0.33-fold higher; 0.3 nM CCL20 vs control) and to a lesser but statistically significant extent, IL-1ß, TNF-α, and MMP-3 (data not shown). Further, the pattern of release was not proportional to the CCL20 concentration in donor FLS culture supernatants (Figure 3A mRNA; Figure 3B protein), suggesting either complex ligand/receptor binding kinetics or receptor downregulation. As expected, RA-affected FLS cultures (Figure 3B) released considerable levels of IL-6 in the absence of CCL20, and CCL20-dependent activation resulted in a 1.59 ± 0.12-fold increase at 0.3 nM. Since CCL20 stimulated COX-2 expression with concomitant elevated levels of prostaglandin release (see below), and because prostanoids are recognized as potent feed-forward/feedback autocoids17, we tested whether effects of CCL20 might be mediated by PGE2, the most abundant of prostanoids in FLS18. Indeed, CCL20 stimulation of IL-6 expression was largely blocked in the presence of a COX inhibitor, but recovered with the addition of 1 μM PGE2 (Figure 3C).
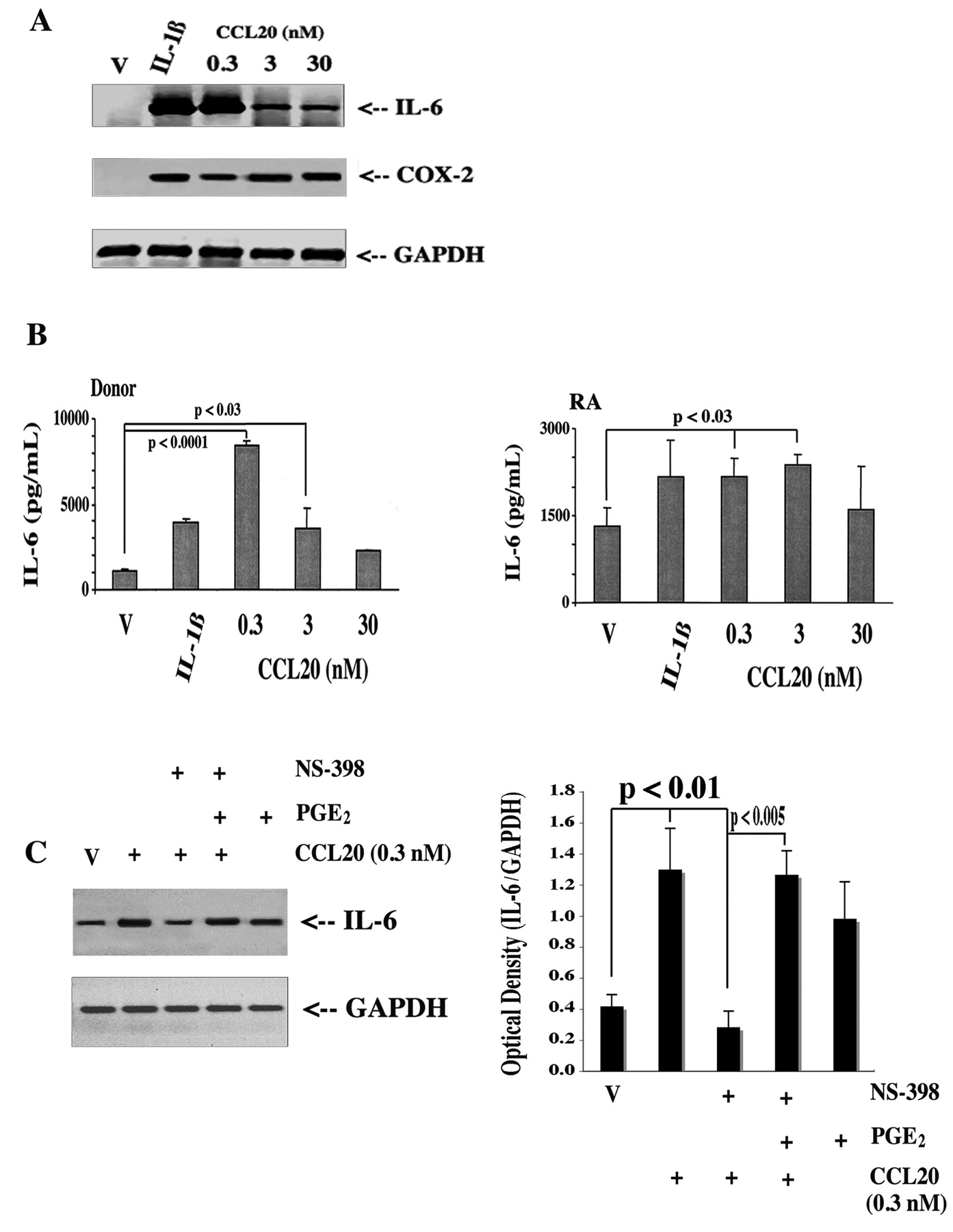
IL-6 protein and mRNA expression induced by CCL20: the role of endogenous prostaglandins. FLS cultures were stimulated with CCL20 (0.3–30 nM) or IL-1ß (2 ng/ml) for 4 h, then RNA was analyzed by RT-PCR for IL-6, COX-2, and GAPDH using specific primers (A). Results are representative of 4 different donors and 4 separate experiments. B. Conditioned media were collected after 72 h and analyzed for IL-6 by ELISA from both donor and RA-derived FLS. Data represent mean ± SEM of triplicate analyses from 4 donor and RA synovia. C. FLS cultures were treated for 4 h with vehicle (V) or CCL20 (0.3 nM) in the presence or absence of 100 nM of the COX inhibitor NS-398 with or without 1 μM PGE2. RNA was isolated and subjected to RT-PCR for IL-6 and GAPDH using specific primers. Representative gel of 3 separate experiments.
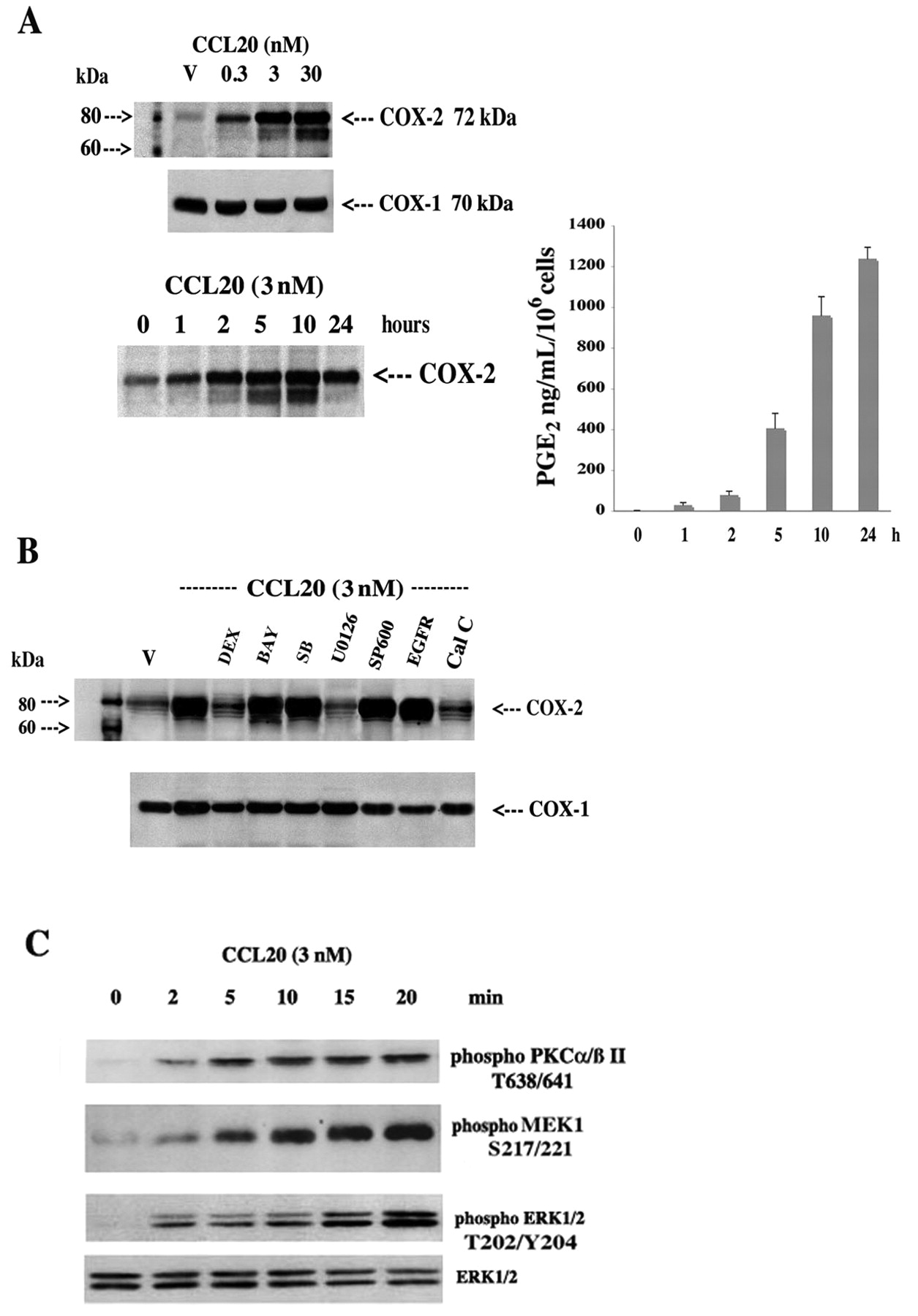
To enhance our understanding of the critical CCL20/PGE2 axis, we investigated the cell signaling and nuclear mechanisms governing CCL20 activation of COX-2 in donor FLS (avoid background COX-2 expression in RA FLS). CCL20 induced a dose- and time-dependent increase in COX-2 expression with a concomitant increase in PGE2 release; COX-2 protein expression was sensitive to the MEK1/2 inhibitor U0126 and the PKC inhibitor calphostin C, but not inhibitors of other key stress, growth, and inflammatory pathways (Figure 4A, 4B). This was confirmed using Western blot analysis of PKCa/ßII/MEK1/ERK1/2 phosphorylation and activation (Figure 4C). Further, CCL20 increased COX-2 promoter activity as judged by luciferase activity generated from a transfected COX-2 promoter-LUC chimeric construct (3.48 ± 0.61-fold increase over control at 3 nM; Figure 5A, 5B). Interestingly, posttranscriptional regulation was also revealed, as CCL20 markedly upregulated luciferase activity from a COX-2 3’UTR-LUC chimera (Figure 5A, 5B): a 7.04 ± 0.83-fold increase over control at 3 nM.
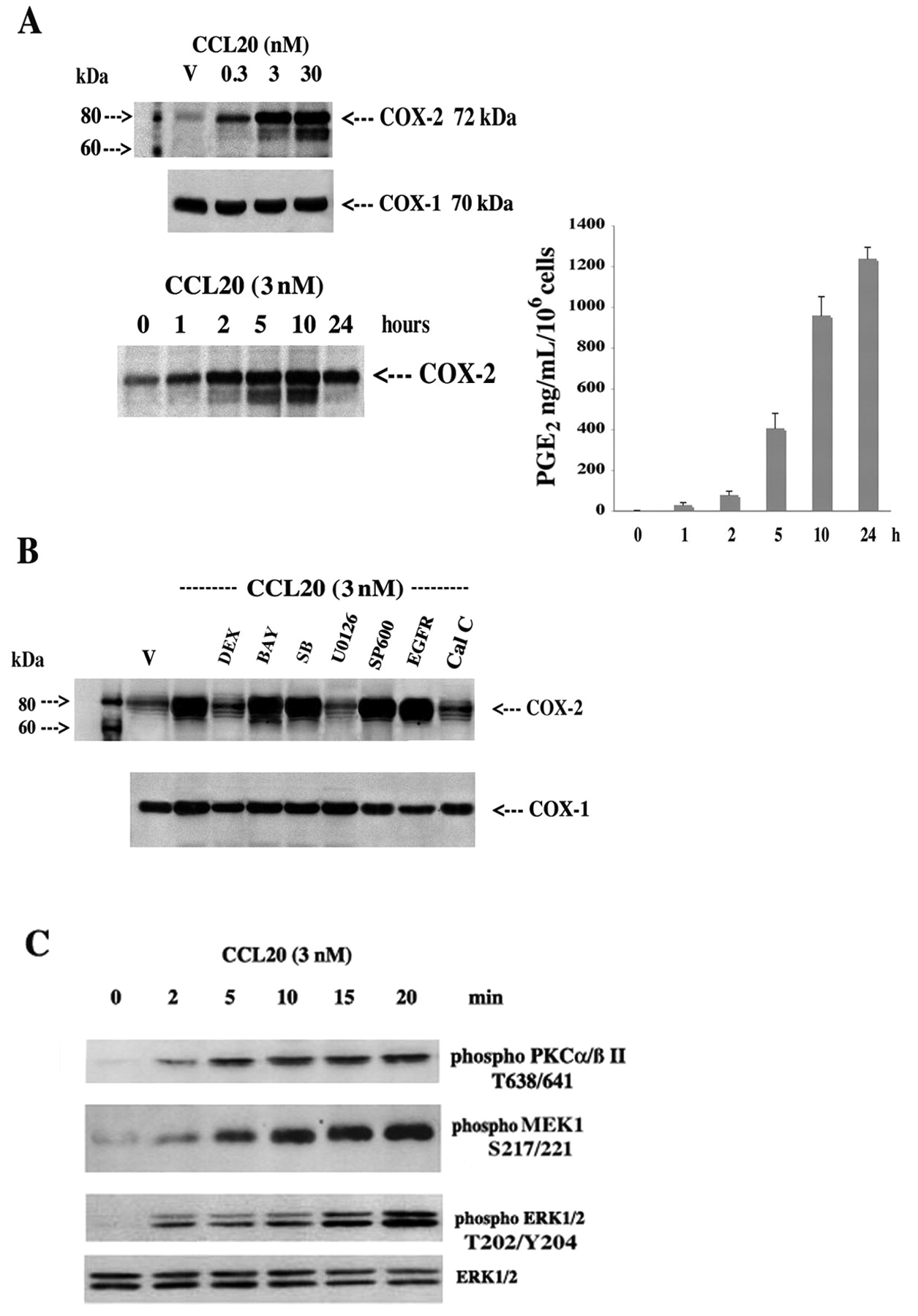
CCL20 regulation of COX-2 and PGE2 involves restricted PKC/MEK1/ERK1/2 activation. A. FLS were treated with vehicle (V) or increasing concentrations of CCL20 (0.3–30 nM) for 4–6 h or with 3 nM of CCL20 for 0–24 h, then cells were extracted for protein and 50 μg were subjected to SDS-PAGE and Western blot analysis using specific antibodies against COX-2 and COX-1. COX-1 serves as a control for protein loading. Culture supernatants were also collected and PGE2 was measured by ELISA; results represent mean ± SEM of 3 separate experiments performed in triplicate. B. Cells were treated with vehicle (V) or 3 nM of CCL20 for 4–6 h in the presence or absence of dexamethasone (DEX; 100 nM), nuclear factor-κB inhibitor Bay-11-7082 (BAY; 2 nM), p38 MAP kinase inhibitor SB202190 (SB; 0.5 μM), MEK1/2 kinase inhibitor U0126 (2 μM), c-Jun terminal kinase inhibitor SP600125 (SP600; 1 μM), epidermal growth factor receptor (EGFR) inhibitor tyrphostin AG1478 (Tyr; 10 μM), and the PKC inhibitor calphostin C (Cal C; 250 nM). C. FLS were deprived of serum for 4 h, then treated with CCL20 (3 nM) for 0 to 20 min and processed for Western blot analysis using total and anti-phospho p44/42 (T202/Y204), anti-phospho MEK1 (S217/221), and anti-PKCa/ß II (T638/641).

CCL20 regulation of COX-2 expression involves downstream transcriptional and posttranscriptional control. A. The human COX-2 promoter construct and the COX-2-3’UTR luciferase chimera. B. FLS were plated at 40% confluence in DMEM supplemented with 10% heat inactivated FBS and antibiotics; 1 μg of LUC-COX-2-3’UTR (luciferase) chimeric fusion construct, BSu36 I COX-2 promoter-LUC, or pGL3-LUC empty vector and 50 ng of a pHSV-TK-driven Renilla luciferase plasmid were cotransfected for 6 h using FuGENE 6. Cells were incubated O/N in complete medium; after change to medium containing 1% FBS (2 h), transfected cells were incubated 16 h with vehicle (V) or CCL20 (0.3–30 nM). Cells were lysed and luciferase activity and protein content were determined. Values are relative light units (RLU) normalized to Renilla luciferase (constitutive activity and control for relative transfection efficiency) and cellular protein content with background subtracted (i.e., pGL3-LUC empty vector).
DISCUSSION
That chronic synovitis, with pannus formation, contributes to the cartilage loss and bone lesions observed in RA joints is well accepted19,20. What is far less clear is how the proliferative lining of FLS orchestrates the recruitment and ultimate behavior of infiltrating inflammatory cell populations, which are almost certainly responsible for the chronicity of the disease21,22. CCL20/CCR6 is an inducible ligand/receptor tandem that is associated with the pathophysiology of RA because of the potent chemokine function in the recruitment and migration of blood-borne inflammatory cells, particularly antigen-presenting cells, which foster both innate and adaptive (antibody) immune responses10,11. This study was conducted to evaluate the role of CCL20/CCR6 in FLS cellular activation, and our results indicate that a classic inflammatory profile was obtained with cytokine, chemokine, MMP, and bioactive lipid release.
CCL20 and CCR6 were highly expressed in unstimulated RA synovium and RA-affected FLS, in accord with studies demonstrating increased levels of CCL20 not only in explants of RA synovium cultured in vivo but also in elevated levels in synovial fluids of RA patients23,24. In contrast, other studies failed to report constitutive expression of CCL20 in RA FLS in culture25, although these differences may be explained by different methodological approaches, as we used both semiquantitative PCR (mRNA) and immunohistochemistry (protein), while other investigators relied on ELISA technology to measure CCL20 released in culture supernatants. One other issue may be that we used primary/first passage RA FLS in an attempt to maintain in situ/in vivo conditioning.
CCR6 expression was detectable in donor, OA, and RA synovium, suggesting that the CCR6 gene may be constitutively expressed (perhaps in most cell types) and primarily the increased secretion of CCL20 may drive that CCL20/CCR6 contribution to RA pathology. Chemokines such as IL-8, CCL2, CCL5, CCL3, and cytokines such as TNF-α and IL-lß have been shown to have a pivotal role in chronic synovitis, a hallmark of RA pathophysiology. They are involved in cellular migration, cellular proliferation, activation of adhesion molecules, and cytokine production6,26,27,28. We show for the first time that CCL20 induced the gene expression of IL-6, IL-lß, and COX-2 and also upregulated the release of IL-6 and MMP-3 in FLS. Moreover, we provide strong evidence that CCL20/CCR6 signal through a PLCL1/PKC/MEK1/2/ERK1/2 pathway with downstream transcriptional and post-transcriptional control mechanisms. It seems likely that CCR6 is coupled to Gαi and/or Gαq-type G-proteins and with only one ligand, in contrast to other chemokine receptors; it would be straightforward to further delineate the signaling cassette. Moreover, we identified COX-2 as an important target gene and our data suggest that PGE2, and perhaps other arachidonic acid metabolites, may mediate some of the CCL20-dependent effects on target gene expression29,30. If so, then the number of potential targets of inflammation increases dramatically, because PGE2, through at least 4 distinct receptor systems, controls a very large proteome in FLS including some 800 membranal, cytosolic, microsomal, mitochondrial, and nuclear proteins regulated17,31,32. In addition, a phosphoproteomic profile of PKA/Akt substrates in FLS demonstrated that 300 substrates were activated within seconds and included adaptor/scaffolding proteins, adhesion molecules, apoptosis regulators, cell cycle regulators, chaperones, cytoskeletal proteins, G-protein regulators, protein kinases and phosphatases, RNA processing and binding proteins, and transcriptional and translational regulators33.
These findings suggest a contributing role of CCL20 in cartilage degradation and place the chemokine at a higher rung in the cytokine/chemokine hierarchy than previously thought. Moreover, increased expression of CCR6 has also been confirmed in tissue biopsies from RA subchondral bone25 and RA synovial B cells9. While anti-TNF-α therapy has been a staple among the biologics used to treat patients with RA when standard disease-modifying antirheumatic drug therapy has shown poor responsiveness, its use has been associated with lymphoma, reactivation of latent infections, lupus-like syndromes, and cardiovascular complications33. Considering this evidence, we suggest a contributing role of CCL20 in the pathogenesis of arthritis and can envisage anti-CCL20 and/or anti-CCR6 blockade as potentially useful objectives for pharmacotherapeutic research.
We have shown that in addition to the well described chemoattractant properties of CCL20 for activated T cells, B cells, and dendritic cells in rheumatoid synovium, the chemokine also potently activates resident FLS lining cells to produce critical inflammatory mediators. CCL20, via CCR6, activates critical signaling pathways involved in the control of a large number of genes through both transcriptional and post-transcriptional regulatory mechanisms. Thus, the chemokine may serve as a critical factor assembling the molecular and cellular components orchestrating local immune responses leading to synovitis. The pleiotropic properties of CCL20 and the specificity with which it acts suggest that the chemokine may serve as an excellent drug target for antirheumatic pharmacotherapy.
Footnotes
-
Supported in part by the Canadian Institutes of Health Research grant no. M11557 (J.A. Di Battista) and the Research Council of St. Joseph University, Beirut, Lebanon (N. Alaaeddine).
- Accepted for publication April 9, 2011.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}