

Abstract
Objective. Systemic sclerosis (SSc) is a life-threatening autoimmune disease characterized by chronic fibrosis of the skin and internal organs. Connective tissue growth factor (CTGF) is believed to be a primary mediator of chronic fibrosis. We assessed the possible association between 7 single-nucleotide polymorphisms (SNP) in the CTGF gene and scleroderma in a French population (registration number 2006/0182).
Methods. We conducted a case-control study with 241 scleroderma patients and 269 controls. Seven SNP were genotyped using the TaqMan system. Univariate and multivariate analyses were performed. In silico electrophoretic mobility shift assay (EMSA), and reverse transcriptase polymerase chain reaction analyses were done to assess the effect of the SNP on CTGF gene expression.
Results. The frequency of the rs9399005TT genotype was significantly lower in SSc patients than in controls. This association remained significant after adjustment for gender. An association was detected between the rs9399005 and the diffuse and limited cutaneous forms. Multivariate analysis between SSc patients and controls taking into account all 7 SNP and sex revealed that only sex and the rs9399005 SNP were associated with disease. DNA analysis by EMSA indicated that the T allele bound nuclear factors that were also bound by the C allele. The binding affinity was higher for the T allele. Analysis of the human database and experiments with human hepatocyte cell line indicated the existence of an alternative transcript containing the rs9399005 polymorphism in its 3’UTR region. In silico analysis indicated that this polymorphism may alter the structure of CTGF messenger RNA.
Conclusion. These findings suggest that CTGF gene polymorphisms may contribute to susceptibility to scleroderma.
- SCLERODERMA
- CONNECTIVE TISSUE GROWTH FACTOR
- POLYMORPHISM
- DISEASE SUSCEPTIBILITY
- ELECTROPHORETIC MOBILITY SHIFT ASSAY
Systemic sclerosis (SSc) is a connective tissue disorder characterized by microvascular abnormalities, autoimmunity and fibrosis, affecting skin and internal organs including lungs and digestive tract. SSc is characterized by deposition of collagen and other extracellular matrix substances in connective tissue, leading to high morbidity and mortality. SSc is a complex, polymorphic, and heterogenous disorder. It is believed to be the consequence of external triggers acting upon a genetically susceptible host. Transforming growth factor beta (TGF-ß) may be central to the initiation of the chronic fibrosing disorder1,2, although its role in the maintenance of the fibrotic phenotype in SSc remains unclear. Unlike TGF-ß, expression of the connective tissue growth factor (CTGF, also known as CCN2), an autocrine stimulator released in response to TGF-ß, correlates well with the extent of skin sclerosis and the severity of pulmonary fibrosis in this disease3. These observations suggest that CTGF is involved in SSc pathogenesis. Moreover, CTGF messenger RNA (mRNA) is constitutively expressed and strongly induced after TGF-ß stimulation in skin specimens from sclerotic SSc lesions, compared to healthy skin control4,5. CTGF is a profibrotic protein that induces proliferation, collagen synthesis, and chemotaxis of fibroblasts. Its presence is associated with excessive matrix protein deposits6, due at least in part to its action on a CTGF response element in the type I collagen (COL1A2) gene promoter7. Subcutaneous injection of TGF-ß into newborn mice results in only a transient fibrotic response, while co-injection of TGF-ß and CTGF results in sustained fibrosis8,9. This work in animal models suggests a 2-step process of fibrosis in SSc: first, TGF-ß initiates the wound healing response and then CTGF promotes sustained and chronic deleterious fibrosis in a TGF-ß-independent manner5. Thus, constitutive overexpression of CTGF by fibroblasts is expected to contribute directly to chronic persistent fibrosis, the characteristic feature of the scleroderma phenotype. A genome-wide association study for regions of SSc susceptibility, performed with a Choctaw Indian population of North America with high incidence of SSc, revealed potential SSc-associated regions. One of them was located at 6q23–27, where the CTGF gene maps10. Association studies between CTGF and SSc have been recently performed that lead to conflicting results. Associations were found with the polymorphism rs6918698 (–945C/G) located in the promoter region in 2 studies11,12. However, these associations were not replicated in other studies13,14. We therefore investigated whether genetic variation of CTGF could confer susceptibility to SSc. We genotyped polymorphisms in this gene and tested for association with SSc in a French European Caucasian population. Associated polymorphism was functionally characterized.
MATERIALS AND METHODS
Patients and controls
We included 510 consecutive unrelated subjects, comprising 241 SSc patients classified according to the LeRoy’s criteria15 and 269 healthy controls, all of French Caucasian origin. Scleroderma patients were recruited by the Internal Medicine Departments in Marseilles (111 patients) and Lille (130 patients). Healthy controls were recruited at the Blood Bank Centre (volunteer blood donors), or included by the Clinical Investigation Centre of the Mediterranean University. They did not suffer from autoimmune disease. Controls were matched by age (± 10 years) and ethnicity to patients. The Ethics Committee of the Mediterranean University approved the study and all patients and controls gave written informed consent for all procedures (Direction Générale de la Santé, registration number 2006/0182).
Blood samples and DNA preparation
We collected 5 to 15 ml of blood on lithium heparin and stored it at –20°C. Genomic DNA was extracted from total blood using a phenol-chloroform method and an Autogen NA2000 (Genework; Pont-à-Mousson, France)16.
Genotyping
Genotype studies were performed using the TaqMan system (allelic discrimination using the 5’nuclease assay, Applied Biosystems, Courtaboeuf, France) with an ABI Prism 7900 Sequence Detector (Applied Biosystems), according to the manufacturer’s instructions. Replication of 50 samples in a double-blind protocol confirmed 100% reproducibility. Seven single nucleotide polymorphisms (SNP) were genotyped: rs6918698, rs1931002, rs9493150, rs12526196, rs12527705, rs9399005, and rs12527379 (Figure 1A). These SNP were chosen for several reasons: (1) SNP rs6918698 (G-945C) mapping in the promoter of the gene was previously studied by Fonseca, et al11 and found to be a functional SNP-associated with susceptibility to SSc; (2) rs1931002, rs9493150, rs9399005, and rs12527379 were selected as Tag SNP according to HapMap Data on the Caucasian reference population. The tag SNP were selected on a region covering the CTGF gene ± 10 kb, according to HapMap. For this selection, correlation groups were defined according to the r2 values. The cutoff value was 0.8; and (3) the rs12526196 and rs12527705 were added to cover the distal 3’ region of the gene (Figure 1A).
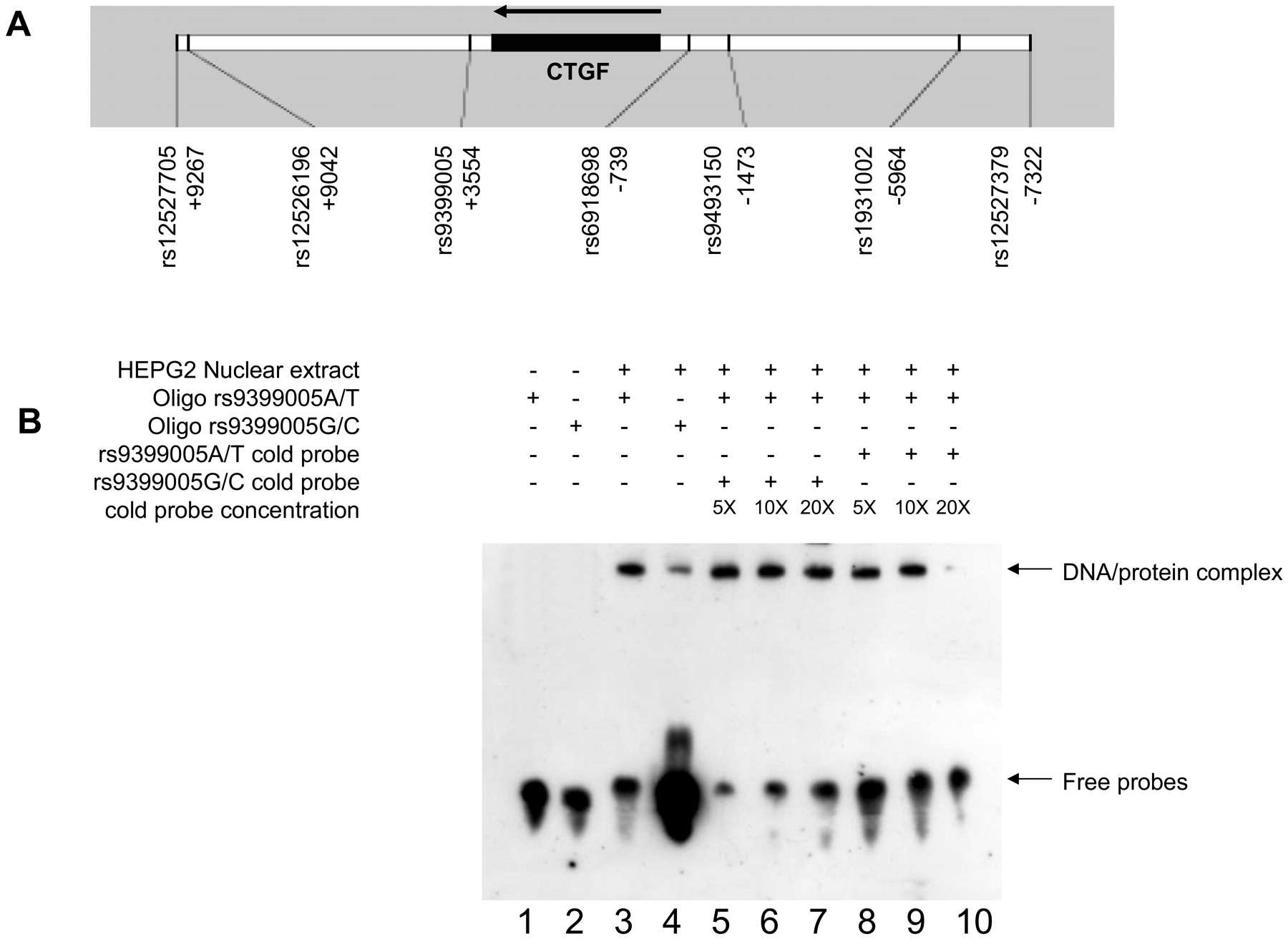
Electrophoretic mobility shift assay (EMSA) for the rs9399005 polymorphism. A. Physical map of the region indicating the 7 polymorphisms tested and their position according to the transcription start point. B. EMSA were performed with or without nuclear extract from a stimulated hepatocyte cell line. In reactions done without nuclear extract, no DNA/protein complex is detected (lines 1 and 2). In reactions with nuclear extract, the rs9399005 T allele probe resulted in the formation of a DNA/protein complex that was also detected with other probe specific from the C allele (lines 3 and 4). However, the binding efficiency was higher with the T allele (line 3). Some competition reactions were carried out with 100 (5×), 200 (10×), and 400 (20×) fmol of unbiotinylated rs9399005A/T and G/C probes (lines 5 to 10). The unbiotinylated rs9399005G/C probe did not compete with the biotinylated rs9399005A/T probe for binding (lines 5 to 7). The unbiotinylated rs9399005A/T probe partially competed with the biotinylated rs9399005A/T probe (at 20× concentration), as shown in line 10.
Statistical analyses
SPSS software was used for statistical analyses. Chi-squared tests were performed to compare genotype frequencies between cases and controls. Fisher’s exact tests were used as appropriate. Because SSc is an autoimmune disorder preferentially affecting women, and because the control group in our study could not be matched for gender, we performed a stepwise binary logistic regression analysis on the whole population including gender and polymorphisms as covariates. Because of the number of statistical tests performed, and to keep the type I error < 5%, p values were corrected as appropriate according to Nyholt’s procedure for multiple testing correction17,18. Associations remained significant after this correction (Table 3). The effective number (Meff) of independent tests was estimated by using the single nucleotide polymorphism spectral decomposition (SNPSpD) Web site (http://genepi.qimr.edu.au/general/daleN/SNPSpD/; accessed October 23, 2009): the Meff value was estimated to be 5.4 in our study. Main associations detected remained significant after correction. Linkage disequilibrium analysis was done with the haploview software.
Nuclear extract preparation
Nuclear extracts were prepared from a human hepatocyte cell line (HEPG2) stimulated for 1 hour with dexamethasone (1μM) and from human normal dermal fibroblasts cell line (GM0038). The nuclear and cytoplasmic extraction reagents from Pierce were used. Nuclear protein concentrations were determined with the bicinchoninic acid protein assay reagent (Pierce).
Electrophoretic mobility shift assay (EMSA)
Complementary single-stranded oligonucleotides were synthesized commercially; they were designed to span a region extending approximately 10 base pairs (bp) on either side of the variant nucleotide, as follows:
Rs9399005 oligonucleotide forward: CTA ACC TAT AA(C/T) GGC CAG AGA GGT ACA AA. Rs9399005 oligonucleotide reverse: TTT GTA CCT CTC TGG CC(G/A) TTA TAG GTT AG.
Complementary strands were annealed by placing reactions (sense and antisense oligonucleotides) in a boiling water bath for 10 min, then cooling to room temperature. Binding reactions were set up with LightShift Chemiluminescent EMSA Kit (Pierce). Aliquots of 20 fmol of complementary DNA were incubated at room temperature for 20 min with 4 μg of nuclear extract in 10 mM Tris, 50 mM KCl, 1 mM DTT, 2.5% glycerol, 5 mM MgCl2, 50 ng/μl poly d(I-C), 0.05% NP-40, pH 7.5. Reactions were loaded onto an 8% non-denaturing polyacrylamide gel and run for 150 min at 110V. Free DNA and DNA/protein complexes were transferred to nylon N+ membranes by capillary action. Binding was detected according to the manufacturer’s instructions (Pierce).
RNA preparation and cDNA synthesis
Total RNA was extracted using the TRIzol reagent (Life Technologies) and following the manufacturer’s instructions. First strand cDNA was synthesized from 2 μg of total RNA using the High-Capacity cDNA Archive Kit (Applied Biosystems) according to the manufacturer’s protocol. To minimize variations in reverse transcriptase efficiency, all samples from a single experiment were reverse transcribed simultaneously.
Polymerase chain reaction (PCR)
PCR amplifications were carried out in 30 μl reactions containing 100 ng DNA or cDNA, 10 mM Tris-HCl pH = 9, 0.1% Triton X-100, 50 mM KCl, 0.2 mg/ml BSA, 1.5 mM MgCl2, 1 μM of each primer, 1 mM of dNTP and 1.5 U Taq polymerase (Invitrogen). Following the initial denaturation step (94°C 5 min), samples were subjected to 35 cycles of PCR consisting of 94°C for 1 min, annealing temperature for 45 s, and 72°C for 45 s. The primers used for exon3-exon4 amplification were exon 3 forward 5’ AGC AGC TGC AAG TAC CAG TG 3’, exon 4 reverse 5’ CCA GGC AGT TGG CTC TAA TC 3’. The primers used to detect the rs9399005 marker in cDNA were forward 5’ AAG TTC AGA AAC AGA CCT AGA GCA 3’ and reverse 5’ TCA GTC TCC ATT AAC CCT GTT G 3’.
RESULTS
The clinical and immunological characteristics of the patients are presented in Table 1. Patients and controls were from a French European Caucasian population; they were all genotyped for 7 CTGF polymorphisms and the association with SSc was tested for the rs6918698, rs1931002, rs9493150, rs12526196, rs12527705, rs9399005, and rs12527379 SNP, in comparison with controls (Tables 2 and 3). All the SNP were in Hardy-Weinberg equilibrium (Tables 2 and 3).
Demographic data and disease characteristics of patients with SSc (n = 241).
Genotype and allele distributions of the CTGF SNP in patients with SSc and controls.
Frequencies of rs9399005 SNP genotypes and alleles in patients with SSc and controls.
Linkage disequilibrium analysis performed on controls indicated that rs12526196 and rs12527705 markers were strongly correlated (r2 value = 1). No other correlation was detected (0.01 < r2 value < 0.44) between the other polymorphisms, indicating that the genotyped markers were good tag SNP.
A difference in CTGF genotype distribution between SSc patients and controls was observed for only the rs9399005T/C polymorphism (Table 3). No association was detected for the other 6 tested polymorphisms (Table 2). In detail, the rs9399005 TT genotype was significantly less frequent in patients with SSc as a whole (2.5%) than in controls [10.0%; p = 0.001, OR = 0.23, 95% CI (0.09–0.56)]. SSc is a heterogeneous disease and cases are usually classified into 2 cutaneous forms based on the extent of skin fibrosis: diffuse cutaneous and limited cutaneous forms. The prevalence of the TT genotype also differed between both these subgroups of patients and controls: 2.6% of the diffuse cutaneous subgroup vs 10.0% of the control group had the TT genotype [p = 0.001, OR = 0.23, 95% CI (0.09–0.56)]; and 2.6% of the limited cutaneous subgroup and 10.0% of the control group had the TT genotype [p = 0.008, OR = 0.24, 95% CI (0.08–0.69); Table 3]. No significant association was observed between the rs9399005 SNP marker and the serum specific autoantibody profile: uncorrected p value of 0.022 for comparison between patients positive for anticentromere antibodies (ACA) and controls and uncorrected p value of 0.998 for comparison between patients positive for antitopoisomerase I (anti-topo I) and controls.
There was a difference in sex ratio between SSc patients (86.3% women) and control subjects (44% women), thus we included sex as a covariate in analyses. A multivariate analysis including sex and the polymorphism rs9399005T/C as covariates confirmed the association between this SNP and SSc disease [sex: p < 0.001, OR = 8.26, 95% CI (5.26–12.82), and rs9399005T/C polymorphism p = 0.001, OR = 0.19, 95% CI (0.07–0.50)].
We also performed an analysis restricted to the female population of patients and controls. The frequency of the rs9399005 TT genotype was significantly lower among the female patients with SSc (2.9%) than among the female control group (11.5%; p = 0.002; Table 4). The frequency of the rs9399005 TT genotype was 2.8% in the female limited cutaneous subgroup vs 11.5% in the female control group (p = 0.005); there was also a trend for association between this SNP and the female diffuse cutaneous subgroup (p = 0.069).
Frequencies of rs9399005 SNP genotypes in cases vs controls according to gender.
Multivariate regression analysis between SSc patients and controls, taking into account all 7 SNPs and sex, revealed that only sex and the rs9399005 SNP were associated with disease. Being a woman was a risk factor [p < 0.001, OR = 8.26, 95% CI (5.03–13.51)] for SSc, while the rs9399005T/T genotype was protective [p = 0.002, OR = 0.21, 95% CI (0.07–0.57)]. The same multivariate analysis done with a limited cutaneous subgroup gave similar results: again, the rs9399005T/T genotype was protective [p = 0.017, OR = 0.25, 95% CI (0.08–0.78)]. Multivariate analysis on the diffuse cutaneous subgroup gave a trend of association only for the rs9399005 SNP (p = 0.054).
We then analyzed the effect of the rs9399005 polymorphism, in vitro by EMSA with nuclear extracts from dexamethasone-stimulated hepatocytes. A DNA/protein complex was formed with both the rs9399005T allele and the rs9399005C allele. However, the binding efficiency was higher with the rs9399005T allele than with the rs9399005C allele (Figure 1B). Some competition reactions were carried out with 100 (5×), 200 (10×), and 400 (20×) fmol of unbiotinylated rs9399005A/T and G/C probes. The unbiotinylated rs9399005G/C probe did not compete with the biotinylated rs9399005A/T probe for binding. The unbiotinylated rs9399005A/T probe partially competed with the biotinylated rs9399005A/T probe (at 20× concentration, no binding was observed confirming the specificity of the binding/band; Figure 1). We performed the same analysis with the nuclear extract from unstimulated human normal dermal fibroblasts (GM00038). Here again, the binding efficiency was higher with the rs9399005T allele than with the rs9399005C allele (data not shown).
Only one CTGF transcript (NM_001901) was found in the genome database: it is 2358 bp long and has a 1083 bp 3’UTR region. Blast analysis of this reference sequence identified another entry describing a longer CTGF transcript with a longer 3’UTR region (CA413027); the untranslated region is 1604 bp long and contains the rs9399005 polymorphism. However, this sequence has been reported only once. To confirm the existence of this transcript, we extracted mRNA from stimulated HEPG2, treated it with DNase and used reverse transcriptase-polymerase chain reaction to test for CTGF transcripts. The cDNA products were tested with 2 pairs of primers (Figure 2A). First, we tested a primer pair covering exon 3 and exon 4. The sizes of the amplified products from genomic DNA and cDNA were different (381 bp and 250 bp, respectively), indicating the absence of DNA contamination. The second pair of primers flanked a short sequence containing the marker rs9399005. An amplification product of 173 bp was obtained from both genomic DNA and stimulated hepatocytes cDNA (Figure 2A). These results confirmed the existence of an alternative CTGF transcript with a longer 3’UTR.
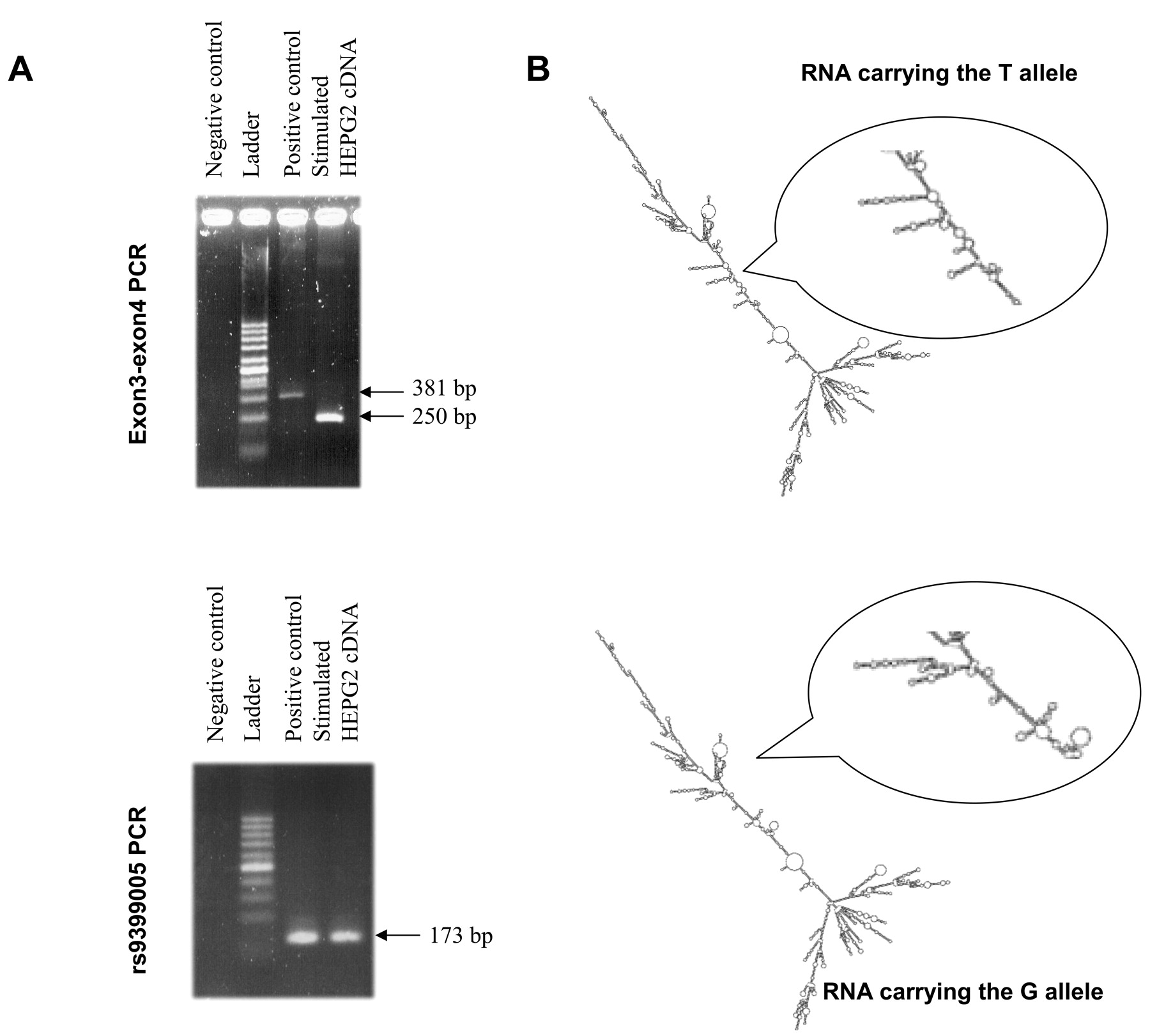
Characterization of the rs9399005 polymorphism. A. Characterization of the different transcripts for the connective tissue growth factor (CTGF) gene. CDNA purified from a stimulated hepatocyte cell line tested by polymerase chain reaction (PCR) with a set of primers amplifies the exons 3 and 4 of the CCN2 gene. A negative control reaction (without template) gave no amplification; a positive control reaction (on genomic DNA) gave a 381 bp amplification product as expected. These primers were also tested on cDNA purified from stimulated hepatocytes. A shorter PCR product was obtained indicating the absence of genomic DNA contamination (250 bp). A second primer set specific for the rs9399005 marker was also tested. Same PCR reactions were done. No amplification products were obtained in the negative reaction. A similar amplification product (173 bp) was obtained either on genomic DNA (positive reaction) or on cDNA purified from stimulated hepatocytes indicating that this region may be a transcript. B. Two-dimensional structure simulation of the CTGF messenger RNA (mRNA) according to the mutation in the rs9399005. The structure of RNA is a major determinant of its function. Successful prediction of RNA tertiary structure is very difficult, but RNA secondary structures can be predicted with relative accuracy and can be biologically useful descriptions. The secondary structure of the longer CTGF mRNA was determined in silico using the Vienna package (http://rna.tbi.uni-vie.ac.at). The same analysis was repeated with the sequences carrying the T and the C alleles. Two different putative secondary structures were defined. These 2 structures present similar energy properties; however, the organization of the 2 central parts differs significantly. A focus on the central region is also presented.
We used the RNAfold program to predict the minimum free energy secondary structure of the long CTGF RNA containing the rs9399005 polymorphism19. Two different structures were predicted (Figure 2B) with similar energies: allelele T: ΔG = –1016.49 kcal/mol; allele G: ΔG = –1018.00 kcal/mol. The 2 predicted secondary structures differed in the central part of the molecules (Figure 2B).
DISCUSSION
Our study shows that the CTGF rs9399005 T/C marker is associated with SSc, and indeed with both diffuse and limited cutaneous forms of the disease. These associations were significant after correction for multiple statistical testing. The sex ratio was different between patients and controls, with women being much more affected by SSc than men, so we tested whether this difference affected the associations identified. Inclusion of sex in the statistical analysis confirmed the association between the rs9399005 marker and SSc.
For women, the CTGF rs9399005 T/C marker was significantly associated with limited forms of the disease (p = 0.005), and there was a trend of association with diffuse forms (p = 0.069). The finding of a trend was probably due to the small number of women presenting the diffuse cutaneous form (n = 60), which is classically the rarer form of SSc. Moreover, the rs9399005 TT genotype was less frequent in women positive for the 2 SSc classical autoantibodies (ACA and anti-topo I) than in control women. Thus, CTGF rs9399005 T/C seems to be a marker of the disease, and is not restricted to a particular subgroup of patients characterized by the extent of skin involvement (cutaneous limited or cutaneous diffuse) or the presence of specific serum autoantibodies (ACA or anti-topo I). Because of the small numbers of patients and to avoid multiple statistical tests, we decided not to test association with other subgroups of patients [for example, patients with pulmonary fibrosis (n = 73) or pulmonary arterial hypertension (n = 26)].
No association was detected between the disease and the other SNP tested (rs6918698, rs1931002, rs9493150, rs12526196, rs12527705, and rs12527379). In particular, no association was detected with the rs6918698 polymorphism (p = 0.62). The same absence of association was found when sex was included as a covariate (p = 0.99). Previous genetic association studies of CTGF in SSc reported discordant results for this marker (also called G-945C11–14). Fonseca, et al11 included 2 study groups, all of United Kingdom origin. In the first group, the authors found an excess of the GG genotype in the SSc group (27.5% of patients) relative to controls (18%, p = 0.03). This was replicated in the independent set (group 2): 32.3% of the SSc patients were GG homozygotes vs 19.9% of the controls (p = 0.001). When the 2 groups were merged, the difference in the frequency of GG homozygotes was statistically significant [30.4% of SSc patients were GG homozygotes vs 19.2% of the controls; p < 0.001, OR = 2.2, 95% CI (1.5–3.2)]. However, Gourh, et al failed to replicate the association between this SNP and SSc in a large cohort of subjects (994 SSc patients and 668 controls), all from North America13; 29% of SSc patients were GG homozygotes vs 29.8% of the controls (p = 0.83). This lack of association was also found by Rueda, et al14. In their study, the GG genotype frequency was not significantly different between cases and controls in 7 independent study populations from Spanish, French, Dutch, German, British, Swedish, and North American origin: the GG genotype frequency was observed between 24.4% and 29.9% in cases, and between 26.6% and 33.7% in controls. Moreover, in our study, the GG frequency in both subgroups (cases and controls) was similar to that observed by Gourh, et al and Rueda, et al13,14.
Insufficient statistical power is one of the possible explanations for these diverse findings, even if this polymorphism is highly informative. However, this is unlikely to be the cause of the discrepancy between the Fonseca and Gourh studies, as they both involved large cohorts of subjects. Fonseca, et al suggest that the disagreement between these studies may be due to different gene-environment interactions in the 2 populations or to differences in the genetic backgrounds (between American and English populations).
According to this second hypothesis, in some populations, the previous associated SNP (rs6918698) may also be in linkage disequilibrium or correlated (r2) with another marker (the functional one) located within or outside the same gene. It is possible that in a different population the associated SNP (rs6918698) is not in linkage disequilibrium or correlated with the causal SNP. In that population, rs6918698 would not be associated with disease but the true disease-associated SNP would be.
The marker highly correlated to the SNP rs6918698, found associated with SSc in the Fonseca study, may be the rs9399005 polymorphism. In our study population these 2 markers are not correlated (r2 = 0.36). Moreover, on the HapMap reference populations, the markers rs9399005 and rs6918698 were genotyped and r2 values are highly variable from one population to the other (0.03 < r2 < 0.74; populations analyzed in HapMap: ASW: African ancestry in Southwest USA, CEU: Utah residents with Northern and Western European ancestry from the CEPH (Centre d’Etude du Polymorphisme Humain; Evry, France) collection, CHB: Han Chinese in Beijing, China, CHD: Chinese in Metropolitan Denver, Colorado, GIH: Gujarati Indians in Houston, Texas, JPT: Japanese in Tokyo, Japan, LWK: Luhya in Webuye, Kenya, MEX: Mexican ancestry in Los Angeles, California, MKK: Maasai in Kinyawa, Kenya, TSI: Tuscans in Italy, and YRI: Yoruban in Ibadan, Nigeria). Thus, in some populations these 2 markers are correlated (r2 > 0.7), while in other populations, the 2 polymorphisms are not correlated (low r2 values), which could explain the difficulty of confirming the association in replicate analysis.
The findings of our genetic association studies involving the CTGF gene suggest an association between the rs9399005 SNP marker of this gene and the disease. EMSA showed that this DNA sequence is involved in a DNA/protein complex, and that the protein binding is more efficient for the T allele. EMSA findings clearly depend on the source of the nuclear extract. Here, we used nuclear extracts from a stimulated HEPG2 that produces CTGF mRNA and confirmed our results on nuclear extracts from human normal skin fibroblasts.
Analysis of the RNA indicated that there are at least 2 alternative transcripts for the CCN2 gene differing by the 3’UTR region. Moreover, the polymorphism rs9399005 may modify the secondary structure of the whole mRNA molecule and may therefore affect stability of the transcript (Figure 2B).
The 3’UTR region can be viewed as a regulatory region required for appropriate expression of many genes20. This region can regulate translation, control the deadenylation and readenylation processes that affect the length of the poly (A) tail and the nuclear export, and can determine subcellular targeting and rates of degradation of mRNA.
A cis-acting determinant, common to both transcripts, acts as a post-transcriptional repressive element21,22. It binds nuclear factors and inhibits gene expression. However, this element alters neither the nuclear export status of cis-linked mRNA nor its intracellular stability, suggesting that there are other regulatory sequences for these functions21.
The rs9399005 polymorphism may affect the transcription or stabilization of CTGF mRNA. We are currently studying patients and controls to determine whether the rs9399005T allele alters the relative abundance of the 2 transcripts.
CTGF is a potential specific target for therapeutic approaches to fibrotic diseases such as SSc. Work with animal models of fibrosis induced by TGF-ß injection have recently shown that it may be possible to inhibit skin fibrosis in vivo by treatment with neutralizing monoclonal antibodies to human CTGF (anti-CTGF antibodies)23. Our findings implicate the CTGF gene in the pathogenesis of SSc, and particularly in the fibrotic process of the disease. Although we have not identified novel diagnostic tools with direct consequences for clinical practice, our work contributes to the understanding of the CTGF pathway in SSc. It consequently provides a basis for further analysis of this SNP in independent cohorts of SSc patients and controls, as replication is needed to validate genetic associations. Modulation of the CTGF pathway can offer new targeted therapeutic strategies to control the progression of fibrosis in SSc.
Acknowledgments
We thank patients and controls for their participation in this study. We thank A. Dessein for helpful discussions and comments on the manuscript.
Footnotes
-
Supported by the Appel d’Offre de Recherche Clinique-Assistance Publique des Hôpitaux de Marseille, by the INSERM (Institut Nationale de la Santé et de la Recherche Médicale), the Groupe Français de Recherche sur la Sclérodermie, and the Clinical Investigation Center.
- Accepted for publication September 18, 2009.








{kind=link}
{kind=link}