

Abstract
Objective. Dendritic cells (DC) contribute to autoimmune disease progression and pathogenesis. Mature DC have been reported to secrete high mobility group box protein (HMGB-1), a novel inflammatory cytokine, via p38 mitogen-activated protein kinase (MAPK) activation. We investigated whether DC are involved in progression of autoimmune diseases followed by secretion of HMGB-1 via p38 MAPK activation in a lupus-prone mouse model.
Methods. FR167653, a specific inhibitor of p38 MAPK, was given orally from 3 months of age in MRL-Faslpr mice. Cultured DC, treated with or without FR167653, were stimulated with tumor necrosis factor-α.
Results. Inhibition of p38 MAPK led to a reduction in the number of CD11c-positive cells, including those with the mature phenotype, in the diseased kidney and spleen, which resulted in improvement of kidney pathology in MRL-Faslpr mice. The number of CD11c-positive cells in circulation was also reduced. HMGB-1 protein and transcripts detected in the diseased kidney, and the number of cells dual-positive for HMGB-1 and CD11c, were reduced by inhibition of p38 MAPK. Maturation of cultured DC and increased cytokines, including HMGB-1, in the supernatant were inhibited by FR167653 treatment. These results suggest that DC are involved in the progression of autoimmune kidney diseases in MRL-Faslpr mice followed by HMGB-1 secretion via p38 MAPK activation.
Conclusion. Our results indicated that DC secrete HMGB-1 via p38 MAPK activation to participate in autoimmunity in MRL-Faslpr mice.
CD11c-positive dendritic cells (DC) are among the most potent antigen-presenting cells. Interaction between DC and T cells is essential for initiation of immune responses. Recent studies indicated that mature DC also influence proinflammatory responses1,2. Immature DC located in peripheral tissues can be induced to undergo phenotypic maturation by a variety of cytokines, such as tumor necrosis factor-α (TNF-α) and interleukin 1β (IL-1β) through activation of some signal transduction pathways. In particular, p38 mitogen-activated protein kinase (MAPK) has been shown to play a critical role in the maturation of DC3,4. DC have been reported to play an important role in the pathogenesis and progression of autoimmune diseases, including kidney diseases5–9.
High mobility group box protein (HMGB-1), which is a DNA-binding nuclear and cytosolic protein, shows inflammatory properties10. Recently, HMGB-1 has been classified as an alarmin, a group of proteins that provide early warning signals to activate innate and adaptive immune systems in response to infection or tissue injury11,12. HMGB-1 has been reported to be involved in the inflammatory processes in autoimmune diseases, including rheumatoid arthritis13,14 and Sjögren’s syndrome15. HMGB-1 is secreted from mature DC as well as activated monocytes/macrophages (Mφ) in response to inflammatory stimuli, such as lipopolysaccharide (LPS) and proinflammatory cytokines10. Necrotic cells have also been reported to release HMGB-1. Released HMGB-1 induces activation of DC via p38 MAPK activation16,17.
The autoimmune disease in MRL/MPJ-lpr/lpr (MRL-Faslpr) mice resembles human systemic lupus erythematosus, characterized by dysregulation of both cellular and humoral immunity18. This strain is particularly valuable as a model in which to investigate the pathogenesis of autoimmune kidney injury. Previously, we reported that inhibition of phosphorylation of p38 MAPK ameliorated kidney injury, resulting in prolonged survival in MRL-Faslpr mice, followed by reduction of cytokine/chemokine production in the kidneys19.
We hypothesized that DC are involved in the progression of autoimmune diseases followed by production of HMGB-1 via p38 MAPK activation. In this study, we show that FR167653 (denoted FR), a p38 MAPK inhibitor, reduced the number of CD11c-positive cells as well as HMGB-1 in the diseased kidney. p38 MAPK inhibition also reduced HMGB-1 excretion from activated DC in vitro.
MATERIALS AND METHODS
Mice
MRL-Faslpr mice were obtained from Charles River Japan Inc., Atsugi, Japan. FR (32 mg/kg/day) dissolved in drinking water was administered orally from 3 months of age. Mice in any group were sacrificed at 6 months of age. Ten MRL/MpJ++ (MRL++) mice were used as negative controls. All procedures in the animal experiments complied with the Guideline for the Care and Use of Laboratory Animals of Kanazawa University.
Histopathology. Kidney
Kidneys were either snap-frozen in OCT compound for cryostat sectioning or fixed in 10% neutral-buffered formalin. Formalin-fixed tissue was embedded in paraffin, and 4-μm sections were stained with periodic acid-Schiff and evaluated by a light microscopy.
Antibodies
The following primary antibodies were used for immunostaining; biotin or fluorescein isothianate (FITC)-conjugated hamster antimouse CD11c antibody, FITC-conjugated hamster anti-mouse CD80 antibody, phycoerythrin (PE)-conjugated mouse anti-mouse I-Ak antibody (BD Biosciences, San Diego, CA, USA), FITC-conjugated rat anti-mouse CC chemokine receptor (CCR7) antibody (Abcam, Cambridge, UK), and rabbit anti-mouse HMGB-1 antibody (BD Biosciences). The secondary antibodies for immunofluorescence staining were streptavidin-Per-CP-Cy5.5 conjugate (BD Biosciences). Rabbit anti-phosphorylated p38 MAPK polyclonal antibody (Santa Cruz Biotechnology, Santa Cruz, CA, USA) was used to detect phosphorylated p38-positive cells.
Immunohistochemical examination
Tissues for immunoperoxidase staining were snap-frozen in OCT (Miles Scientific, Naperville, IL, USA) and stored at –80° C. CD11c and HMGB-1-positive lesions were examined using monoclonal antibodies (10 μg/ml, ×1000, respectively). These lesions were identified using the avidin-biotin complex technique or the Envision method (Dako, Glostrup, Denmark). In addition, phosphorylated p38 MAPK was evaluated within the kidneys. The presence of phosphorylated p38 MAPK was detected as described19.
The positive area of CD11c and HMGB-1 was evaluated from the whole area of cortex and outer medulla in the complete sagittal kidney section, and expressed as a percentage/glomeruli or visual field using MacScope version 6.02.
Dual staining
To determine the phenotypes of CD11c-positive cells, dual-labeling immunofluorescence immunohistochemistry was performed. Briefly, sections were first incubated overnight with biotin-or FITC-conjugated anti-mouse CD11c antibody. After rinsing in phosphate buffered saline, streptavidin-Per-CP-Cy5.5 conjugate was added for biotin-conjugated antibody. Then, FITC-conjugated anti-mouse CCR7 or FITC-conjugated anti-mouse CD80 antibody, or PE-conjugated anti-mouse I-Ak antibody was added, and the sections were incubated overnight. Adobe Photoshop (Adobe Systems, San Jose, CA, USA) was used for image handling, and the 3-color channels were handled separately.
Flow cytometric analysis
A single-cell suspension was obtained from spleens as described20. Following cell isolation, red blood cells were lysed using FACS lysis solution (BD Biosciences). Heparinized blood was collected from the tail vein of the mice. FITC-or allophycocyanin (APC)-conjugated hamster anti-mouse CD11c antibody, FITC-conjugated hamster anti-mouse CD80 antibody, biotin- or PE-conjugated mouse anti-mouse IAk antibody, and APC-conjugated rat anti-mouse CCR7 antibody (Biolegend, San Diego, CA, USA) were used for detection of cell-surface markers. To detect phosphorylated p38 MAPK, cells were fixed and permeabilized using BD Phosflow™ following the manufacturer’s protocol (BD Biosciences). The cells were then stained with PE-conjugated antiphospho-p38 MAPK antibody (BD Biosciences). The frequencies of cells expressing cell-surface markers were determined by analyzing a minimum of 10,000 cells using FlowJo software (Tree Star, Palo Alto, CA, USA).
Real-time reverse transcription-polymerase chain reaction (RT-PCR)
To determine transcripts of HMGB-1, total RNA was extracted from the whole kidneys. Reverse transcription was performed as reported19. Quantitative real-time RT-PCR were performed on the ABI Prism 7900HT sequence detection system (Applied Biosystems, Foster City, CA, USA) using 384-well microtiter plates. Reactions were performed in a total volume of 20 μl, containing 1 μl of cDNA sample, TaqMan Gene Expression Assays, and TaqMan Universal PCR Master Mix (Applied Biosystems), using the universal temperature cycles. Assay identifiers of TaqMan Gene Expression Assays were Mm00801666_m1 for murine HMGB-1 and Mm00446953_m1 for murine β-glucuronidase (GUS). The mRNA expression in each sample was finally described after correction with GUS expression.
Cell culture and treatment
Bone marrow-derived DC were prepared as described21,22. Briefly, bone marrow cells were suspended at 2 × 105 cells/ml in 10-mm dishes with RPMI-1640 medium supplemented with 10% fetal bovine serum with 20 ng/ml recombinant mouse granulocytemacrophage colony-stimulating factor (GM-CSF; R&D Systems, Minneapolis, MN, USA) in a humidified atmosphere of 5% CO2/95% air at 37° C. On Day 3, another 10 ml of this medium was added. On Days 6 and 8, half the culture supernatant was collected and centrifuged. The cell pellet was resuspended in 10 ml of fresh medium containing 20 ng/ml GMCSF and returned to the original plate. On Day 10, CD11c cells were purified using MACS beads (Miltenyi Biotec, Auburn, CA, USA) following the manufacturer’s protocol. Purified CD11c-positive cells were harvested for 24 h with or without FR167653 (1 × 10−6 M) in the presence of TNF-α (20 ng/ml; R&D Systems).
Detection of cytokines in supernatants of cultured DC by ELISA
HMGB-1 levels in the supernatants of cultured DC were determined using an HMGB-1 ELISA kit (Shino-Test, Sagamihara, Japan). IL-12 levels and IL-23 levels were analyzed using BD OptEIA™ mouse IL-12 (p40) ELISA Set and mouse IL-23 (p19/p40, IL23) ELISA Ready-Set-Go (eBioscience, San Diego, CA, USA), respectively. All analyses were performed according to the respective manufacturer’s protocol.
Statistical analyses
The data represent means ± SEM. Statistical significance was determined by ANOVA and Kruskal-Wallis analyses.
RESULTS
Frequency of CD11c-positive cells, including mature phenotype, was increased according to disease activity in spleens of MRL-Faslpr mice
Flow cytometric analysis was performed to determine whether CD11c-positive cells contribute to the pathogenesis of MRL-Faslpr mice. The frequency of CD11c-positive cells was determined in splenocytes of MRL-Faslpr and MRL++ mice. The frequency of CD11c-positive cells was increased during disease progression (Figure 1). Moreover, the frequency of MHC class II, CD80, CCR7-positive mature phenotype was also increased in the spleens of MRL-Faslpr mice (Figure 1).

The frequency of CD11c-positive cells was analyzed in splenocytes of MRL-Faslpr and MRL++ mice. The frequency of CD11c-positive cells was increased during disease progression. Moreover, the frequency of MHC class II, CD80, CCR7-positive mature phenotype was also increased in the spleen of MRL-Faslpr mice. Isotype control is shown in each analysis. The data are representative of more than 3 experiments.
p38 MAPK activation contributed to maturation of CD11c-positive splenocytes
To determine the effect of p38 MAPK signaling on the maturation of DC, phosphorylation of p38 MAPK was analyzed by flow cytometry (Figure 2a). p38 MAPK-activated CD11c-positive cells included cells with the CCR7- and MHC class II-positive mature phenotypes (Figure 2b). In contrast, the frequencies of these mature phenotypes were reduced in the phosphorylated p38 MAPK-negative CD11c-positive population (Figure 2c).
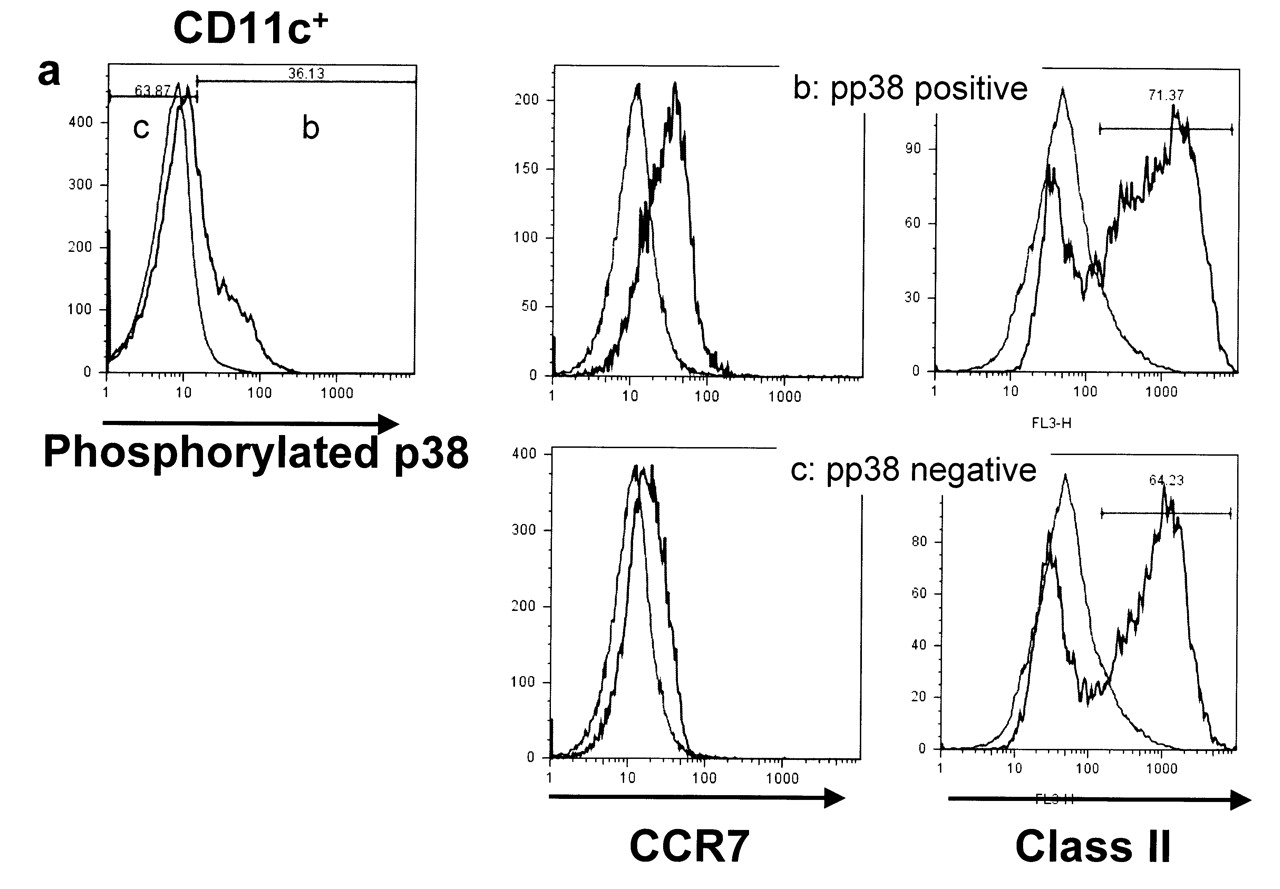
(a) To determine the influence of p38 MAPK signaling on maturation of dendritic cells, phosphorylation of p38 MAPK was analyzed by flow cytometry. (b) p38 MAPK-activated CD11c-positive cells contained CCR7- and MHC class II-positive mature phenotypes. (c) In contrast, the frequencies of these mature phenotypes were reduced in the phosphorylated p38 MAPK-negative CD11c-positive population. Isotype control is shown in each analysis. Data are representative of more than 3 experiments.
Inhibition of p38 MAPK decreased infiltration of CD11c-positive cells, including those with mature phenotype, in kidneys of MRL-Faslpr mice
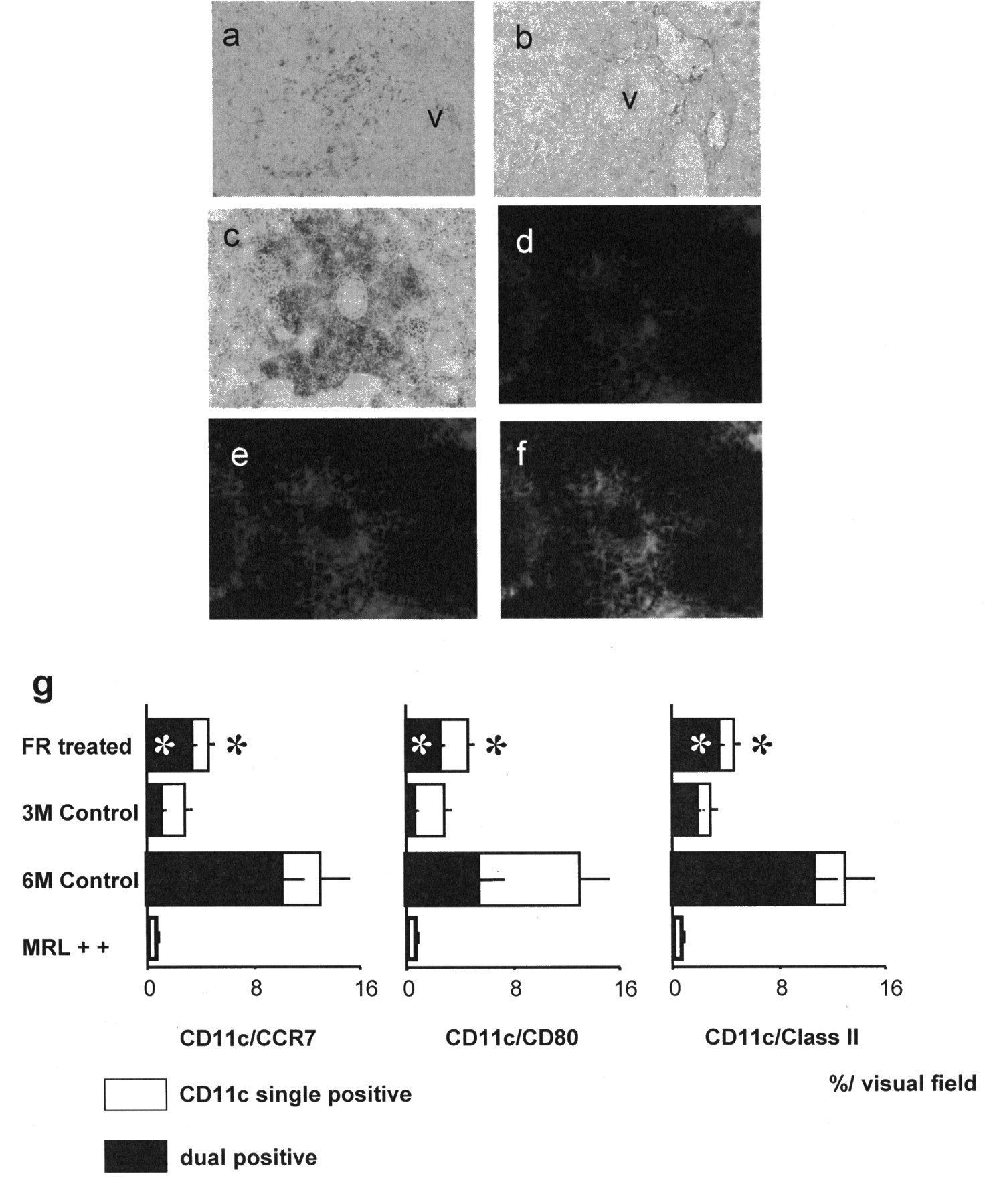
Phosphorylated p38-positive cells were detected in perivascular lesions (Figure 3a), and were decreased by administration of FR (Figure 3b), suggesting that p38 MAPK activation is involved in autoimmune kidney injury. Kidney pathology in untreated mice consisted of severe proliferative glomerulonephritis and interstitial/perivascular damage. In contrast, renal pathological changes were attenuated but not abrogated in FR-treated MRL-Faslpr mice, as compared to those of MRL++ mice19. CD11c-positive cells were detected in perivascular and periglomerular lesions in diseased kidneys (Figure 3c, 3d). The area of CD11c-positive cells, expressed as percentage/visual field, was examined by computer analysis. At 6 months of age, CD11c-positive cells were decreased to 4.6% ± 0.6% in treated mice, as compared to 13.0% ± 3.6% in untreated MRL-Faslpr mice (Figure 3c, 3g). To determine the presence of mature DC, dual immunofluorescence immunohistochemistry labeling for CD11c (Figure 3d) and CCR7 (Figure 3e) was performed. In untreated MRL-Faslpr mice, cells dual-positive for CD11c and CCR7 were detected in perivascular lesions at 6 months of age (Figure 3f). In treated mice, the area of cells dual-positive for CD11c and CCR7 (Figure 3g) was decreased in comparison to untreated MRL-Faslpr controls (3.3% ± 0.2%, 10.7% ± 3.2%, respectively; p < 0.05). Moreover, the numbers of CD80- or MHC class IIpositive CD11c-positive cells were also decreased in the FRtreated group (Figure 3g).

(a) Phosphorylated p38-positive cells were detected in perivascular lesions in untreated MRL-Faslpr mice. (b) The number of phosphorylated p38-positive cells was decreased by inhibition of p38 MAPK. This result indicated that FR167653 inhibited p38 MAPK activation. (c, d) CD11c-positive cells were detected in perivascular and periglomerular lesions in the diseased kidneys. (e) CCR7-positive cells were located in the same lesions as CD11c-positive cells. (f) Cells dual-positive for CD11c and CCR7 were detected in perivascular lesions. Kidney pathology was determined by staining with the indirect avidin-biotinylated peroxidase complex method and dual-labeled immunofluorescence immunohistochemistry method (magnification × 200). (g) The number of CD11c-positive cells, expressed as percentage/visual field, was determined by computer analysis. In treated mice, the numbers of CD11c-positive cells and cells dual-positive for CD11c and CCR7 as well as CD80/class II were significantly decreased as compared to untreated 6-month-old control MRL-Faslpr mice. *p < 0.05 compared to untreated 6-month-old control MRL-Faslpr mice. FR: FR167653; v: vessel.
Inhibition of p38 MAPK decreased mature CD11c-positive cells in spleens of MRL-Faslpr mice
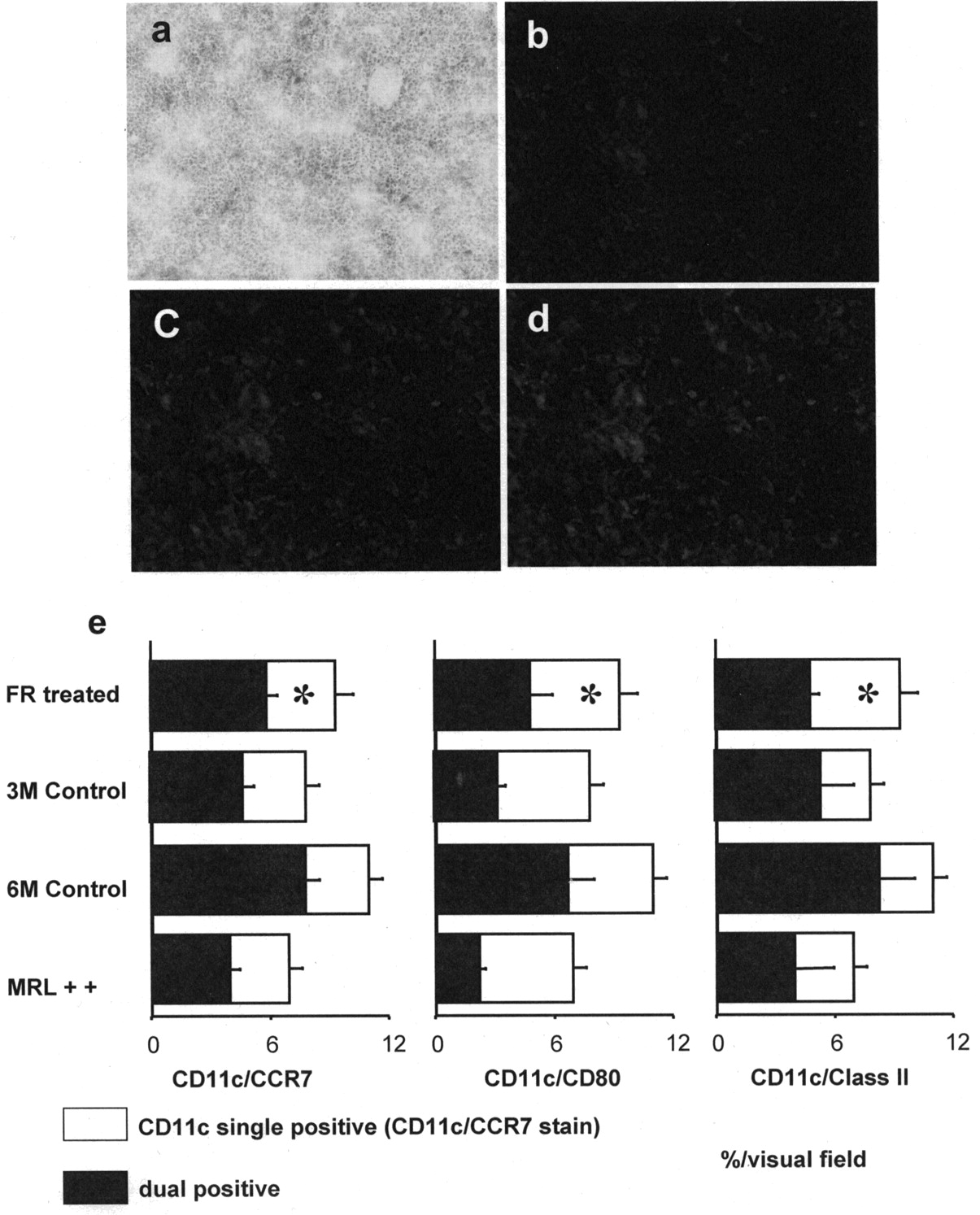
CD11c-positive cells were located in the T cell areas in the spleens of MRL-Faslpr mice at 6 months of age (Figure 4a, 4b). In treated MRL-Faslpr mice, CD11c-positive cell numbers tended to be reduced, but the difference was not significant in comparison to untreated controls (9.5% ± 1.7%, 10.9% ± 1.1%, respectively; Figure 4a, 4e). The area of cells with the mature phenotype dual-positive for CD11c (Figure 4b) and CCR7 (Figure 4c) was decreased as compared to untreated MRL-Faslpr mice at 6 months of age (7.9% ± 0.4%, 5.8% ± 0.3%, respectively, p < 0.05; Figure 4d, 4e). Moreover, the numbers of CD80- or MHC class II-positive CD11c-positive cells were also decreased in the FR-treated group (Figure 4e).

(a, b) CD11c-positive cells were detected in the T cell area in spleens from MRL-Faslpr mice. (c) CCR7-positive cells were located in the same lesions as CD11c-positive cells. (d) Cells dual-positive for CD11c and CCR7 were detected in the T cell area. Spleen pathology was determined by staining with the indirect avidin-biotinylated peroxidase complex method and dual-labeled immunofluorescence immunohistochemistry method (magnification: a × 100; b–d × 200). (e) In treated MRL-Faslpr mice, the number of CD11c-positive cells tended to decrease, but the differences compared to untreated mice were not significant. The numbers of mature dendritic cells dual-positive for CD11c and CCR7 as well as CD80/class II were significantly decreased compared to untreated MRL-Faslpr mice at 6 months of age. *p < 0.05 compared to untreated 6-month-old control MRL-Faslpr mice. FR: FR167653.
CD11c-positive cells were decreased by p38 MAPK inhibition in the circulation
Flow cytometric analysis was performed to evaluate the effect of the inhibition of p38 MAPK on circulating CD11c-positive cells. CD11c-positive cells were detected at a level of 9.9% ± 0.4% at 6 months of age. In mice treated with FR, CD11c-positive cell numbers in hemolyzed blood were reduced to 6.3% ± 0.6% in comparison to untreated MRL-Faslpr controls at 6 months of age (Figure 5a, 5b).

(a) The percentage of CD11c-positive cells was analyzed by flow cytometry; results of representative experiments are shown. (b) In treated mice, the number of CD11c-positive cells in the circulation were significantly decreased compared to untreated 6-month-old control MRL-Faslpr mice. *p < 0.05 compared to untreated 6-month-old control MRL-Faslpr mice. FR: FR167653.
HMGB-1-positive area was reduced by p38 MAPK inhibition in kidneys
Immunohistochemical analysis was performed to investigate involvement of HMGB-1 in kidneys of MRL-Faslpr mice. The HMGB-1-positive area was expressed as the percentage/glomeruli or visual field by computer analysis. High levels of HMGB-1 expression were noted within the glomeruli as well as infiltrates of mononuclear cells in the interstitium (Figure 6a). HMGB-1-positive area was also detected in perivascular lesions in the diseased kidneys (Figure 6b). The percentages of HMGB-1-positive area were 11.3% ± 2.0% in glomeruli and 28.3% ± 3.5% in interstitium in untreated MRL-Faslpr mice at 6 months of age. Glomerular and interstitial HMGB-1-positive areas were reduced in treated MRL-Faslpr mice as compared to untreated controls (6.0% ± 1.0% and 13.3% ± 1.5%, respectively; Figure 6c, 6d). In addition, HMGB-1 staining was also detected in the T cell area of spleens from MRL-Faslpr mice, and this was reduced by p38 MAPK inhibition (data not shown).

(a) High levels of HMGB-1 expression were observed within the glomeruli as well as mononuclear cell infiltrates in the interstitium. Kidney pathology was determined by staining with the indirect avidin-biotinylated peroxidase complex method (original magnification × 400). (b) HMGB-1-positive area was also detected in perivascular lesions in the diseased kidneys (original magnification × 200). (c, d) The percentage of HMGB-1-positive area was significantly decreased in glomeruli as well as the interstitium in FR-treated MRL-Faslpr mice as compared to untreated 6-month-old control MRL-Faslpr mice. (e) mRNA levels of HMGB-1 in the whole kidney were analyzed using real-time RT-PCR. In the kidneys of FR-treated MRL-Faslpr mice, HMGB-1 transcript levels were reduced as compared to untreated 6-month-old control MRL-Faslpr mice. (f) CD11c-positive cells and HMGB-1-positive area were colocalized in perivascular lesions in the diseased kidneys at 6 months of age. Some CD11c-positive cells were also positive for HMGB-1 (arrowheads). Kidney pathology was determined by staining with the indirect avidin-biotinylated alkaline phosphatase method for CD11c, with the indirect Envision peroxidase complex method for HMGB-1 (original magnification × 400). (g) Numbers of dual-positive cells in the diseased kidney were significantly decreased by inhibition of p38 MAPK. (h) In the spleen, cells dual-positive for CD11c and HMGB-1 were located in the T cell area (original magnification × 100). (i) Number of dualpositive cells in the spleen of treated MRL-Faslpr mice was also reduced compared to untreated 6-month-old control MRL-Faslpr mice. *p < 0.05 compared to untreated 6-month-old control MRL-Faslpr mice.
HMGB-1 transcript levels were reduced in kidneys from treated MRL-Faslpr mice
Real-time RT-PCR was used to determine the mRNA levels of HMGB-1 in the whole kidney. In kidneys of treated MRL-Faslpr mice, HMGB-1 transcript levels were reduced compared to untreated controls (Figure 6e).
Both CD11c-positive cells and HMGB-1-positive areas were colocalized in diseased kidney and spleen from MRL-Faslpr mice
It has been reported that mature DC secrete HMGB-1 via p38 MAPK activation16,17. Dual-labeling immunohistochemistry was performed to examine the effects of FR on the expression of HMGB-1 on CD11c-positive cells. Both CD11c-positive cells and HMGB-1-positive areas were colocalized in perivascular lesions in the diseased kidney at 6 months of age (Figure 6f). Some CD11c-positive cells were also positive for HMGB-1. The numbers of dual-positive cells were significantly decreased by inhibition of p38 MAPK compared to untreated controls (0.3% ± 0.0% and 0.8% ± 0.1%, respectively; p < 0.05; Figure 6g). In the spleen, cells dual-positive for CD11c and HMGB-1 were located in the T cell area (Figure 6h). The number of dual-positive cells was also reduced in the spleen of MRL-Faslpr mice compared to untreated controls (3.1% ± 0.1%, 4.2% ± 0.3%, respectively; p < 0.05; Figure 6i).
Maturation of DC was inhibited by administration of FR167653
In GM-CSF-derived bone marrow DC, stimulation with TNF-α increased the frequency of phosphorylated p38 MAPK-positive CCR7-positive phenotype as compared to control DC (42.0% ± 0.7%, 33.0% ± 1.6% in MRL-Faslpr mice; 66.3% ± 6.1%, 46.8% ± 6.2% in MRL++ mice, respectively; p < 0.05). In contrast, inhibition of p38 MAPK signaling decreased the frequency of phosphorylated p38 MAPK-positive CCR7-positive DC as compared to TNF-α-treated DC in each group (21.9% ± 3.2% in MRL-Faslpr mice, 31.5% ± 0.3% in MRL++ mice; p < 0.05; Figure 7a, 7b, 7c). In addition, FR treatment also decreased the number of phosphorylated p38 MAPK-positive class II-positive DC as compared to TNF-α-treated DC (29.1% ± 1.5%, 43.4% ± 1.5% in MRL-Faslpr mice; 35.8% ± 1.3%, 58.3% ± 13.4% in MRL++ mice, respectively; p < 0.05; Figure 7d).
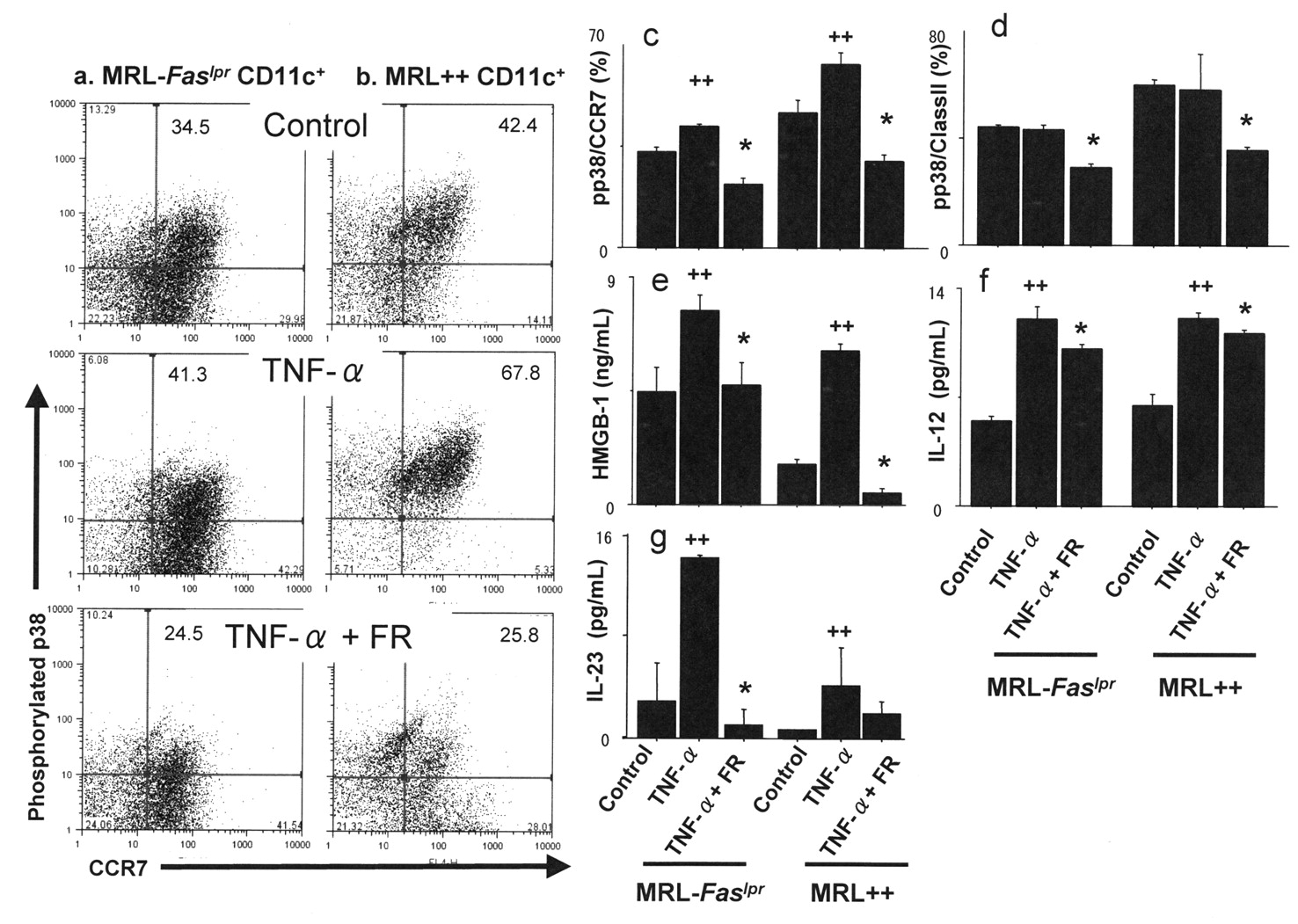
(a–d) Stimulation with TNF-α increased the frequency of phosphorylated p38 MAPK-positive CCR7-positive phenotype compared to control dendritic cells. Inhibition of p38 MAPK signaling decreased the frequency of phosphorylated p38 MAPK-positive CCR7-positive dendritic cells as well as phosphorylated p38 MAPK-positive class II-positive dendritic cells. (e, f, g) In cultured dendritic cells, TNF-α stimulation enhanced HMGB-1, IL-12, and IL-23 levels in the supernatants. Increased concentrations of these cytokines were decreased by treatment with FR167653. ++p < 0.05 compared to controls; *p < 0.05 compared to TNF-α-stimulated group.
Concentrations of HMGB-1, IL-12, and IL-23 in supernatants of cultured DC were reduced by FR167653 treatment
In cultured DC, TNF-α stimulation enhanced HMGB-1, IL-12, and IL-23 levels in the supernatants. However, the enhanced concentrations of these cytokines were reduced by treatment with FR in cultured DC from MRL-Faslpr mice as well as MRL++ mice (Figure 7e, 7f, 7g).
DISCUSSION
Our results suggested that DC are involved in the progression of autoimmune kidney diseases in MRL-Faslpr mice, followed by secretion of HMGB-1 via p38 MAPK activation. In the diseased kidney in particular, the numbers of CD11c-positive cells, including those with the mature phenotype, were reduced in treated mice. Interestingly, the number of CD11c-positive cells in circulation was decreased in accord with disease activity. HMGB-1 protein and transcripts were detected in the diseased kidney, and both were decreased by administration of FR167653. Moreover, CD11c-positive cells were located adjacent to the HMGB-1-positive area in the diseased kidney as well as the spleen. The number of dual-positive cells was reduced by inhibition of p38 MAPK. Maturation of DC and increased HMGB-1, IL-12, and IL-23 concentrations, induced by stimulation with TNF-α, were inhibited by FR167653 treatment in vitro. Therefore, we considered DC were involved in progression of autoimmune diseases in MRL-Faslpr mice, followed by production of cytokines, including HMGB-1, via p38 MAPK activation.
It has been reported that immature DC differentiate into mature DC, which show upregulation of CCR7 to migrate to regional lymph nodes via secondary lymphoid-tissue chemokine/CCR7 interaction23,24. It has been established that mature DC migrate toward regional lymph nodes, where they interact with T cells. Previous studies indicated that maturation of DC induces autoimmune responses in mouse models25–27. These reports suggested that mature DC may play an important role in the pathogenesis of autoimmune diseases.
Our findings indicated that the p38 MAPK pathway is critical for the maturation of immature DC. Similarly, stimuli such as TNF-α, LPS, or contact sensitizers induced maturation of DC via p38MAPK activation3. In our experiment, it should be noted that the inhibition of p38 MAPK reduced the number of mature DC in the diseased kidney and spleen. In support of this, the blockade of p38 MAPK has been shown to inhibit maturation of DC3. p38 MAPK has been reported to regulate the expression of CCR7 and migration of maturing DC to macrophage inflammatory protein-3β and secondary lymphoid chemokines28. Further, DC from peripheral blood and spleen in a murine lupus model were shown to be chemoattracted toward secondary lymphoid chemokines29. Moreover, in acute lung inflammation models, mature DC were detected in the injured lung30. In this study, anti-secondary lymphoid chemokine antibody injection increased the number of DC within the injured lung, thus exacerbating pulmonary inflammation. These observations may explain our findings that the inhibition of p38 MAPK prevents not only maturation but also migration of DC from peripheral organs, resulting in improving autoimmune kidney diseases in our model.
p38 MAPK is involved in regulation of the expression of many proinflammatory cytokines31. Therefore, reduction of mature DC would partly be due to the reduction of microenvironmental stimuli, such as proinflammatory cytokines. Moreover, reduction of the total number of DC could reduce the number of cells with the mature phenotype. In addition, we noted that DC in circulation were reduced in accord with disease activity. These findings suggest that inhibition of p38 MAPK activation induces systemic reduction of DC in MRL-Faslpr mice.
Recent studies showed that HMGB-1, which is released or secreted from necrotic cells, activated Mφ, and mature DC, possesses inflammatory properties10. HMGB-1 acts as an alarmin, a class of molecules that activate antigen-presenting cells and enhance the development of immune responses11,12. It has been reported that HMGB-1 is expressed in various tissues, such as synovial tissue, from patients with rheumatoid arthritis14, liver tissues from hepatitis models32, and some tumor cells10. Our findings suggest that HMGB-1, which may be secreted from mature DC through p38 MAPK activation, plays a role in the pathogenesis of autoimmune kidney diseases. Mature DC secrete HMGB-1, which is crucial for the signal transduction events that maintain DC maturation and for the control of T cell function16. Further, HMGB-1 induced phenotypic maturation of immature DC. Messmer, et al reported that HMGB-1 induced DC maturation and secretion of proinflammatory cytokines, such as IL-1β and TNF-α17. In that report, maturation of DC induced by HMGB-1 was shown to be dependent on p38 MAPK activation, using the p38 MAPK inhibitor. These results raise the possibility that HMGB-1 contributes to the maturation of DC in an autocrine-paracrine loop.
In addition, the stimulation of TNF-α induced the secretion of IL-12 and IL-23 as well as HMGB-1 in vitro, which was reduced by inhibition of p38 MAPK. DC have been reported to promote production of IL-17 by CD4-positive T cells via secretion of IL-12 and IL-2333. These results raise the possibility that DC play an important role in activation of the Th17 pathway in this model. However, more detailed studies are needed to determine the association between DC and the Th17 pathway in this model.
We observed that CD11c-positive cells are involved in autoimmune kidney injury in MRL-Faslpr mice. Our results indicated that dendritic cells secrete HMGB-1 through p38 MAPK activation to participate in autoimmunity in this model.
Footnotes
- Accepted for publication September 5, 2008.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}