

Abstract
Objective. Osteopontin (OPN) is an extracellular matrix glycoprotein that has been recognized as a potential inflammatory cytokine. The function of OPN is modulated by protease digestion, and a thrombin-cleaved form of OPN is involved in the pathogenesis of various inflammatory disorders. We examined thrombin-cleaved OPN products in synovial fluid from patients with rheumatoid arthritis (RA) and osteoarthritis (OA).
Methods. Synovial fluid samples were obtained from knees of 20 patients with RA and 111 patients with OA. Thrombin-cleaved OPN product was determined using Western blotting. Levels of thrombin- cleaved and full-length OPN in synovial fluid were determined by ELISA. Synovia were analyzed by immunohistochemistry using an antibody specific to the thrombin-cleaved form.
Results. Immunoblotting showed the presence of thrombin-cleaved OPN in synovial fluid from patients with RA and OA. ELISA results showed no difference between concentrations of full-length OPN in the synovial fluid of RA and OA patients; however, thrombin-cleaved OPN concentrations in RA synovial fluid samples were roughly 30-fold higher compared with OA samples (p < 0.001). Synovial fluid concentrations of thrombin-cleaved OPN in RA did not correlate with C-reactive protein levels. Immunohistochemistry of the synovium showed stronger reactivity in RA than in OA samples.
Conclusion. Local generation of thrombin-cleaved OPN was increased in RA joints. Thrombin-cleaved OPN may be a useful biochemical marker of RA.
Rheumatoid arthritis (RA) is a chronic inflammatory disease characterized by pathologic changes in joints, including synovitis, cartilage degradation, subchondral bone erosion, and alterations in cellular immune responses. Proinflammatory cytokines, including interleukin 1β (IL-1β), IL-6, and tumor necrosis factor-α (TNF-α), which are produced mainly by macrophages, are involved in the development of synovitis1–3.
Recent studies have shown that osteopontin (OPN), an extracellular matrix glycoprotein, is a potential inflammatory cytokine4–6 and modulates a variety of pathological conditions7,8. OPN is also expressed in synovial tissue and cartilage from patients with RA and osteoarthritis (OA)9,10, suggesting an involvement in the pathogenesis of inflammatory arthritis11.
OPN is a highly phosphorylated and sulfated glycoprotein, with a molecular weight of roughly 32 kDa. Variable sizes up to 75 kDa have been reported7. It is expressed in bone-forming cells, as well as in mesenchymal cells from the uterus, placenta, kidney, and nervous system12. During inflammation and wound-healing, OPN is also expressed by cells related to both innate and adaptive immunity, such as activated T lymphocytes, macrophages, and resident fibroblasts5,13. OPN contains several cell adhesive domains, including an arginine-glycine-aspartate (RGD)-containing domain that interacts with cell-surface integrins αvβ3, αvβ1, and αvβ5. Proteolytic modification of OPN by thrombin cleavage reveals cryptic binding sites for α9β1 and α4β1 integrins, preferentially expressed by neutrophils and by monocytes and lymphocytes, respectively4,7,14,15. The newly exposed binding sites within OPN, SVVYGLR in humans and SLAYGLR in mice and rats, promote adhesion and migration of leukocytes and neutrophils through these alternative sites in an RGD-independent manner7,15 (Figure 1). The presence of the thrombin-cleaved form of OPN is well correlated with various inflammatory disease activities16.

In terms of RA pathogenesis, inflammatory cells such as macrophages, neutrophils, and lymphocytes are infiltrated into the diseased synovium. Moreover, activation of the coagulation cascade in RA synovial fluid, resulting in increased thrombin activity, has been found17. It is conceivable that the thrombin-cleaved form of OPN participates in the development of arthritis18. The purpose of our cross-sectional study was to quantify this form of OPN in synovial fluid from patients with RA and OA and to compare these levels between diseases.
MATERIALS AND METHODS
Patients and samples
All patients with RA had knee joint arthritis and fulfilled the revised criteria for RA of the American College of Rheumatology (ACR)19. All patients with OA fulfilled the ACR clinical and radiological diagnostic criteria20. Synovial fluid samples were obtained at the time of surgery or before intraarticular injection of hyaluronic acid from 20 patients with RA and 111 patients with OA. Synovial fluid was centrifuged at 15,000 × g for 15 min, and the supernatants were stored at –80°C until analyzed.
The group with RA consisted of 16 women and 4 men with a mean age of 66.9 years and a mean body mass index (BMI) of 22.5 kg/m2. The group with OA consisted of 78 women and 33 men with a mean age of 73.9 years and mean BMI of 23.6 kg/m2. Synovial membrane samples were obtained at the time of surgery from 7 patients with RA and 23 with OA of the knee. C-reactive protein (CRP) and matrix metalloproteinase-3 (MMP-3) were measured in sera as a marker of inflammation. All patients gave informed consent, and the study was approved by the local ethics committee.
Western blot analysis
Immunoblotting of synovial fluid was done to examine the presence of the N-terminal half of thrombin-cleaved osteopontin (OPN N-half). Synovial fluid was examined from 3 OA patients, 2 RA patients, and one control patient with a meniscus tear. From each synovial fluid sample, 0.15 ml was diluted twice with phosphate buffered saline (PBS), added to 20 μl of DEAE Sepharose Fast Flow (GE Healthcare UK Ltd., Buckinghamshire, UK), mixed for 30 min at room temperature, washed 5 times with PBS, then eluted with 1.0 ml of 0.7 M NaCl in PBS. The eluates were diluted twice with 2×sodium dodecyl sulfate (SDS) buffer [4% SDS, 20% glycerol, 125 mM Tris HCl (pH 6.8), 10% 2-mercaptoethanol], boiled, and applied on Western blotting with horseradish peroxidase-labeled anti-human OPN (O-17) rabbit IgG Fab’ (IBL, Gunma, Japan). Antibody (O-17) is raised against synthetic peptides corresponding to the internal sequence of mouse OPN (17LPVKVTDSGSSEEKLY32) and can bind both non-thrombin-cleaved OPN (full-length) and OPN N-half21 (Figure 1).
Enzyme linked immunosorbent assay
An ELISA (IBL) was applied to quantify the levels of non-thrombin-cleaved OPN (OPN full-length) and OPN N-half. For the OPN N-half ELISA, Immuno Module Plates (Nalge Nunc, Rochester, NY, USA) were coated with anti-OPN N-half (34E3) mouse monoclonal antibody (in 0.1 M carbonate buffer, pH 9.5) at 4°C overnight, then blocked with 1% bovine serum albumin in PBS containing 0.05% NaN3 at 4°C overnight. The mouse monoclonal antibody (34E3) specifically reacts to SLAYGLR and SVVYGLR, exposed by thrombin cleavage of mouse and human OPN, respectively. The OPN N-half ELISA system does not recognize full-length OPN and detects OPN after thrombin cleavage21 (Figure 1). Sample and standard proteins were diluted with 1% BSA, 0.05% Tween 20 in PBS, added to each well, and incubated at 37°C for 1 h. After 7 washes with washing buffer (0.05% Tween 20 in phosphate buffer), 100 μl of horseradish peroxidase (HRP)-labeled anti-human OPN (O-17) rabbit polyclonal antibody was added to each well and incubated for 30 min at 4°C. After 9 washes with washing buffer, 100 μl of tetramethyl benzidine buffer as a substrate was added to each well and incubated for 30 min at room temperature in the dark. Color development was stopped by addition of 100 μl of stop solution (1 N H2SO4). Optic density of each sample was measured at 450 nm.
To quantify the levels of non-thrombin-cleaved OPN (OPN full-length), the human osteopontin assay kit (IBL) was used with 2 antibodies (O-17 and 10A16). A mouse monoclonal antibody (10A16) is raised against synthetic peptides corresponding to the internal sequence of human OPN (155KSRSFQVSDEQYPDATDE172) and can bind to OPN full-length but not OPN N-half (Figure 1).
The percentage of N-half was expressed as a percentage (OPN N-half divided by all OPN (OPN N-half plus OPN full-length).
Immunohistochemistry
Expression of thrombin-cleaved OPN in synovial membranes was determined by immunohistochemistry using the avidinbiotin complex method. Synovial membrane samples were fixed in 10% buffered formalin, embedded in paraffin, and cut into 4-μm thick sections. After deparaffinization with xylene and rehydration through a series of graded ethanol solutions, the sections were pretreated in 10 mM citrate buffer, pH 6.0, in a microwave oven for 5 min for antigen retrieval. Sections were treated with superblock solution (Scytek Laboratories, Logan, UT, USA). After washing, sections were incubated in 0.3% H2O2 in methanol for 15 min to block endogenous peroxidase activity, blocked with 2% bovine serum albumin, and incubated with the primary antibody [anti-OPN N-half (34E3) mouse monoclonal antibody] overnight at 4°C. Sections were washed with PBS and reacted with the secondary antibody (biotinylated goat anti-mouse IgG antibody) for 1 h at room temperature. After washing, they were incubated with avidin-peroxidase complex for 30 min. Sections were then developed with diaminobenzidine tetrahydrochloride substrate solution and counterstained with hematoxylin.
The results of immunoreactivity for synoviocytes and subintimal tissues were identified using the point system of Salter22, as follows: For synoviocytes, no staining = 0 points; staining of < 25% of synoviocytes = 1 point; staining of 25%–75% of synoviocytes = 2 points; and staining of > 75% of synoviocytes = 3 points. For subintimal tissues: no staining = 0 points; focal weak staining of subintimal tissues = 1 point; focal strong staining of subintimal tissues = 2 points; and extensive strong staining of subintimal connective tissue = 3 points. The protocol was tested with 3 observers (MH, YN, AS) to standardize the scoring system.
Statistical analysis
The Mann-Whitney U-test was used to determine the differences between values in RA and OA patients. Correlation analyses were performed for serum biomarker levels of RA, including CRP and MMP-3, and synovial fluid levels of OPN N-half in patients with RA using Spearman’s rank correlation test. Correlation between levels of OPN N-half and age and BMI were also estimated. P values < 0.05 were considered significant.
RESULTS
Expression of thrombin-cleaved OPN protein
The antibody O-17 reacted with full-length and N-half OPN with molecular weights of 48 kDa and 30 kDa, respectively, in synovial fluids from patients with OA (Figure 2, lanes 1–3) and RA (Figure 2, lanes 5, 6). A control sample showed no band (Figure 2, lane 4). These results showed that all samples of OA and RA contained full-length OPN, and that the levels of N-half OPN were considerably elevated in patients with RA (Figure 2, lanes 5, 6), although faint bands were rarely detected in OA samples (Figure 2, lane 1).
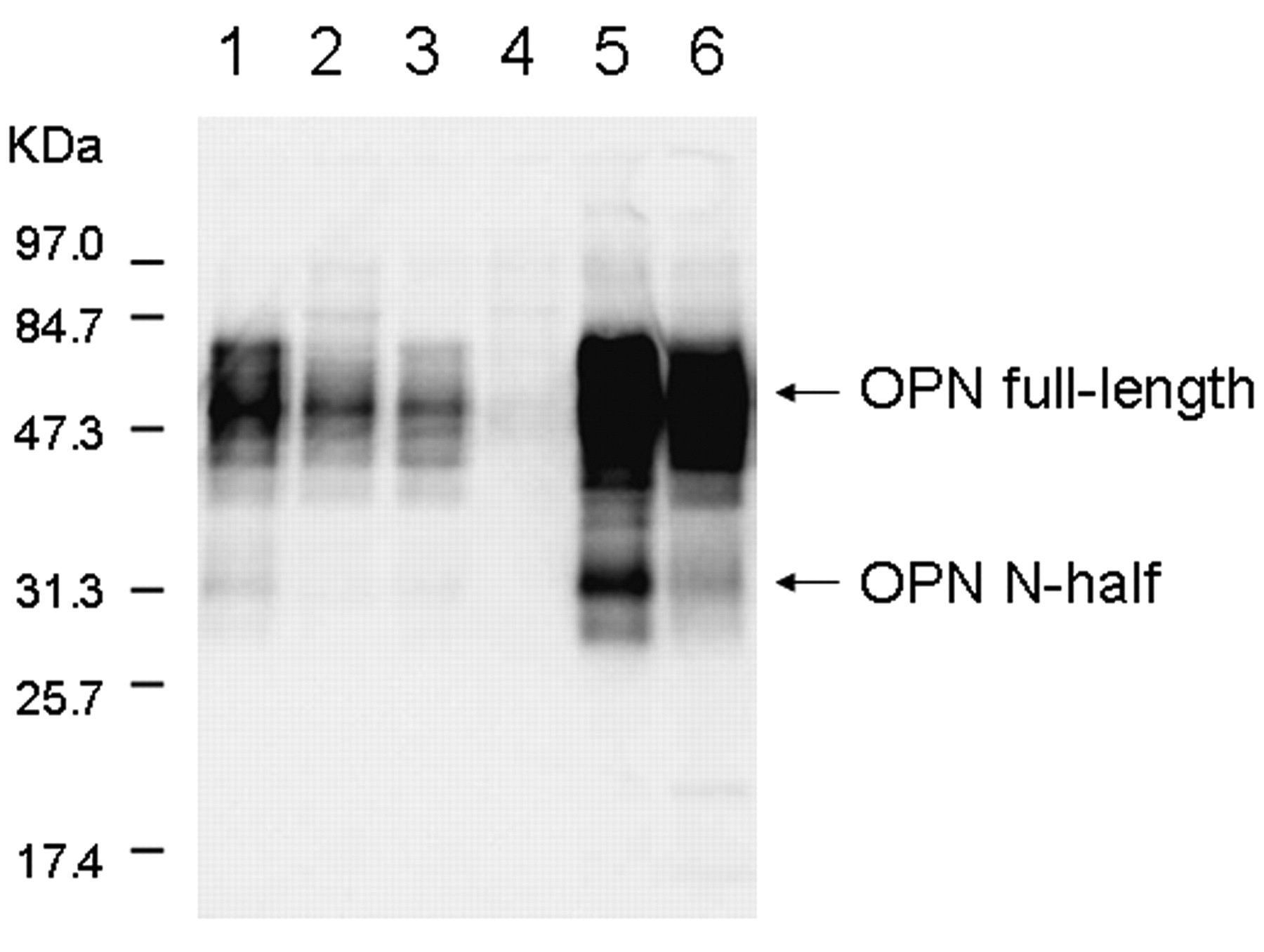
Western blot analyses by HRP-labeled anti-human OPN (O-17) Fab’. Synovial fluid results from patients with OA (lanes 1–3), a control patient (lane 4), and patients with RA (lanes 5, 6) are shown. Synovial fluid from patients with RA contained a considerable amount of thrombincleaved OPN.
Concentrations of full-length and thrombin-cleaved OPN in synovial fluid
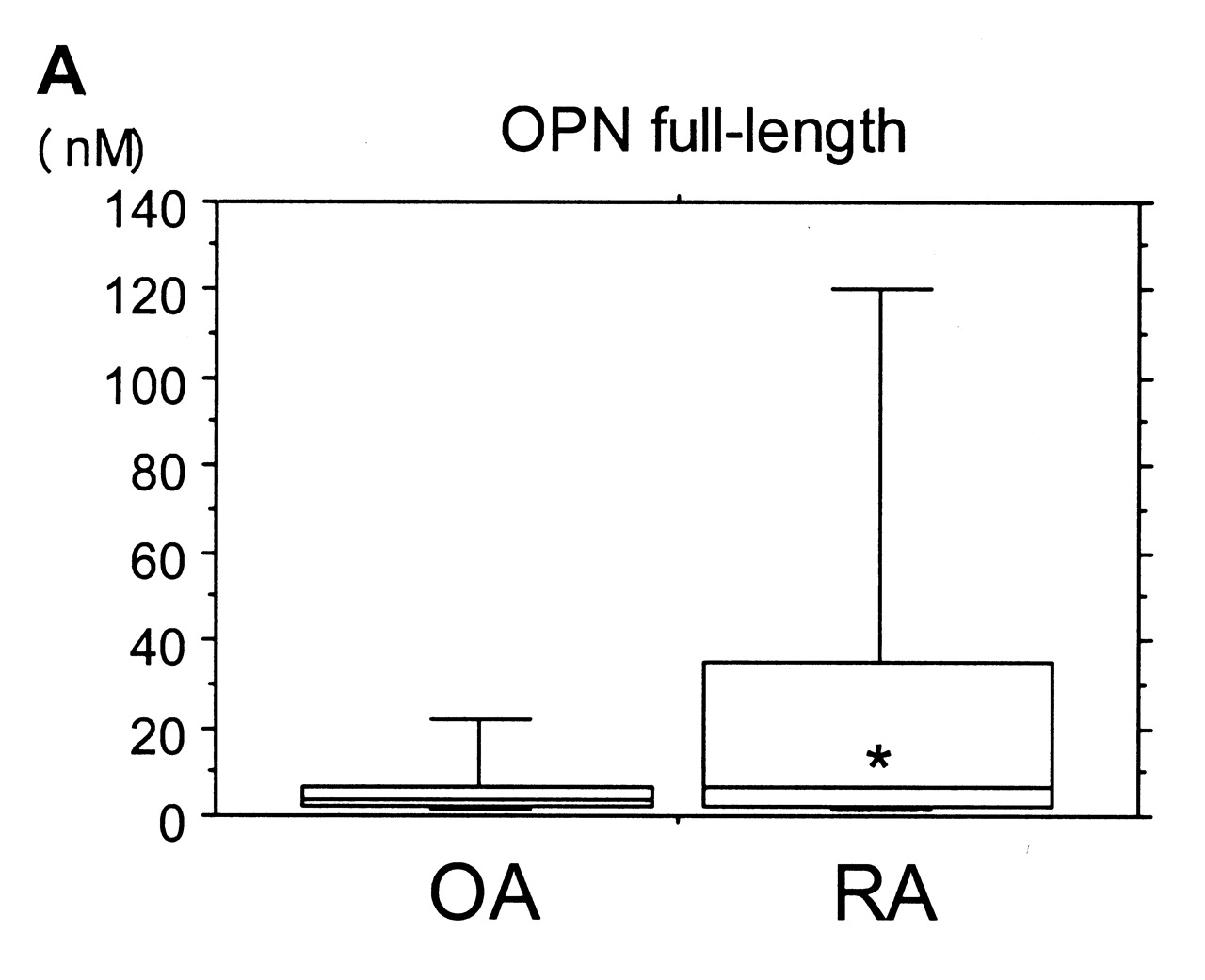
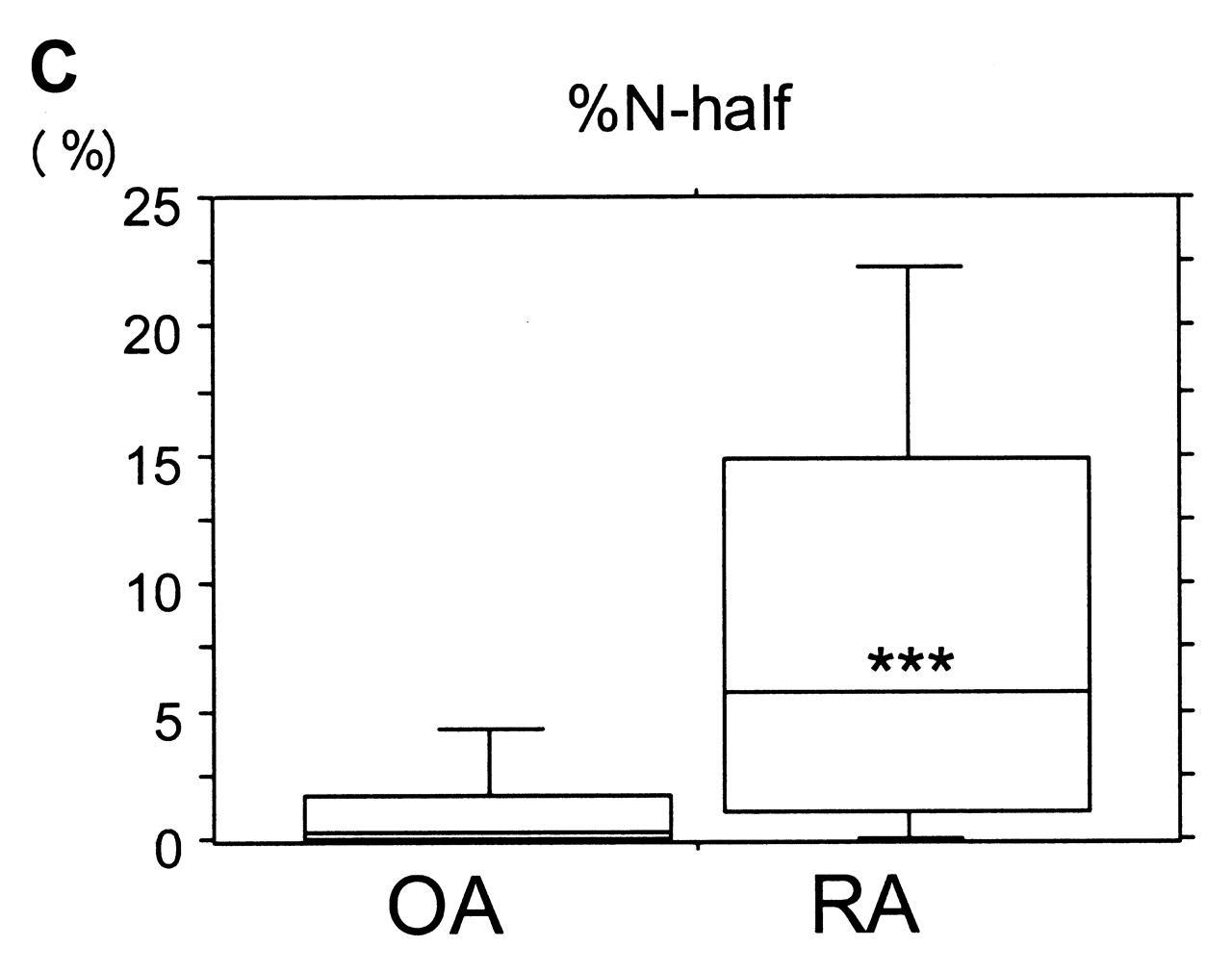
ELISA results showed a tendency for higher levels of full-length OPN in synovial fluid from RA over OA samples, but differences were not significant. In contrast, N-half OPN levels in synovial fluid of RA samples (median 0.376 nM) were roughly 30-fold higher (p < 0.001) compared with OA samples (median 0.013 nM). In addition, percentage of N-half was significantly higher in RA samples than in OA samples (p < 0.001; Figure 3).

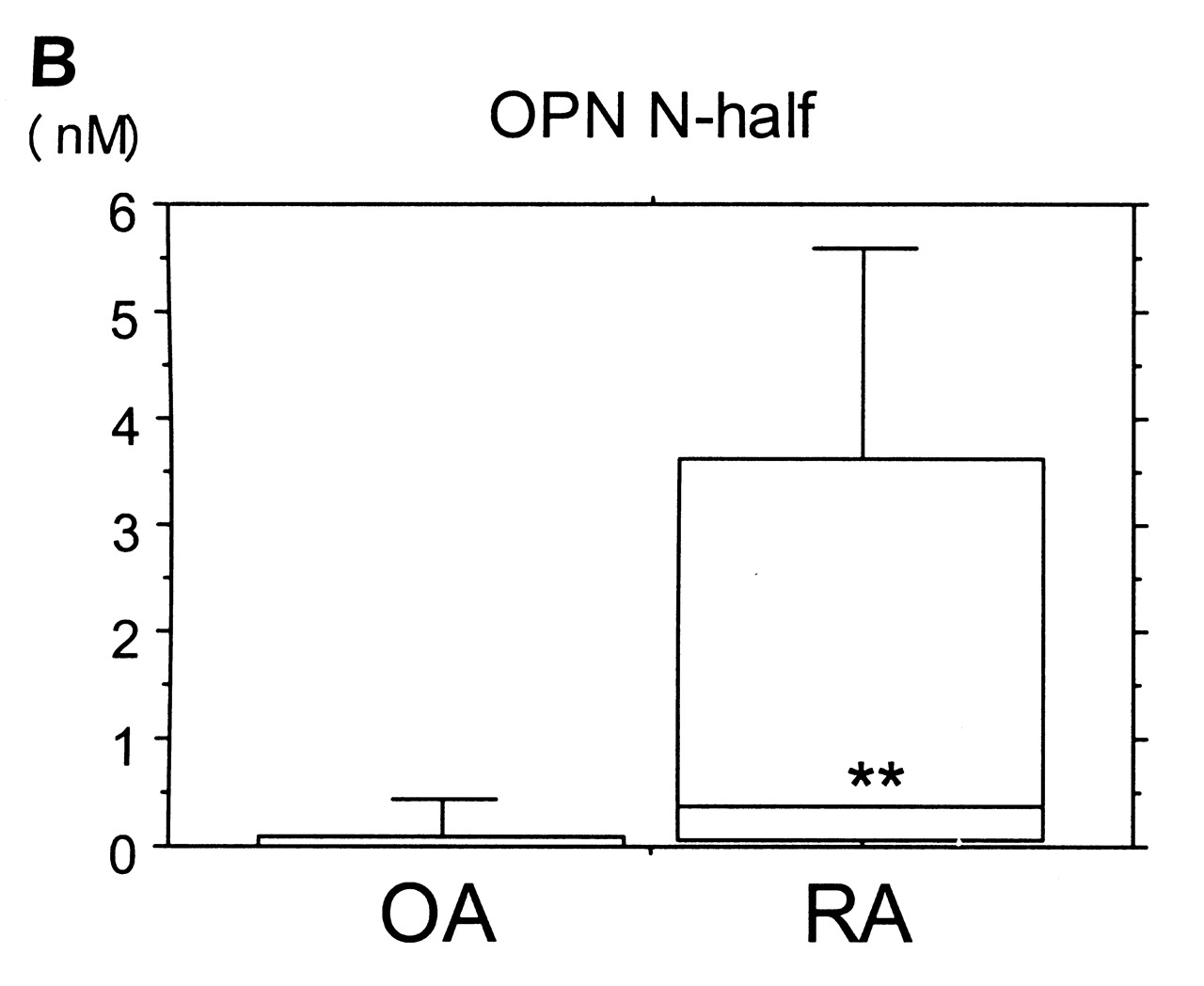

Synovial fluid concentrations of OPN full-length (A) and OPN N-half (B), and percentage of N-half (C). Lower and upper lines in the box represent 25th and 75th percentiles, with the median marked in the box. Bars show range of 10th and 90th percentiles. *p = 0.174, **p < 0.001, ***p < 0.001.
All OA patients had serum CRP levels within the normal range, whereas patients with RA showed elevated serum CRP, with mean level of 2.6 μg/ml. However, no significant correlation was found between serum CRP levels and synovial fluid OPN N-half levels in patients with RA (r = 0.343, p = 0.137). The mean serum MMP-3 level in RA patients was 239 ng/ml. We found no significant correlation between serum MMP-3 and synovial fluid levels of OPN N-half (r = 0.497, p = 0.116). Similarly, synovial fluid levels of OPN N-half did not correlate with patient age (r = –0.029, p = 0.457) or BMI (r = 0.018, p = 0.858).
Localization of thrombin-cleaved OPN in synovial membrane
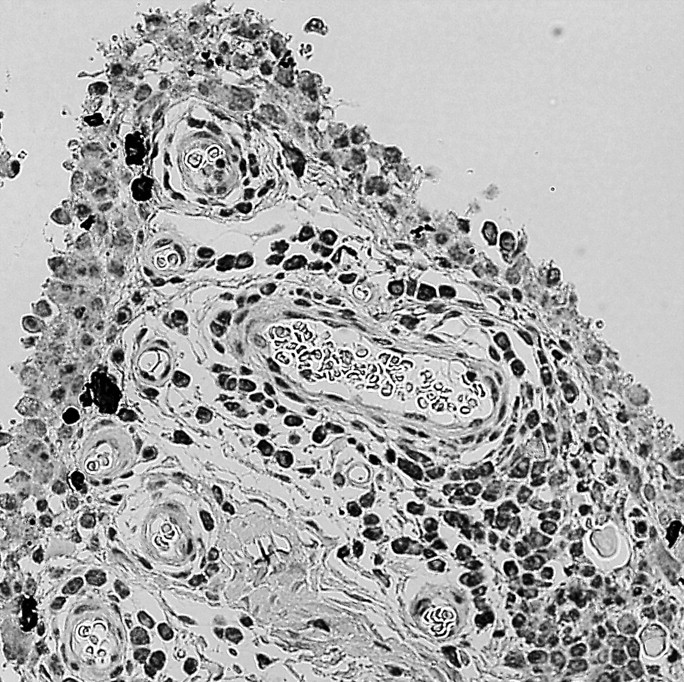
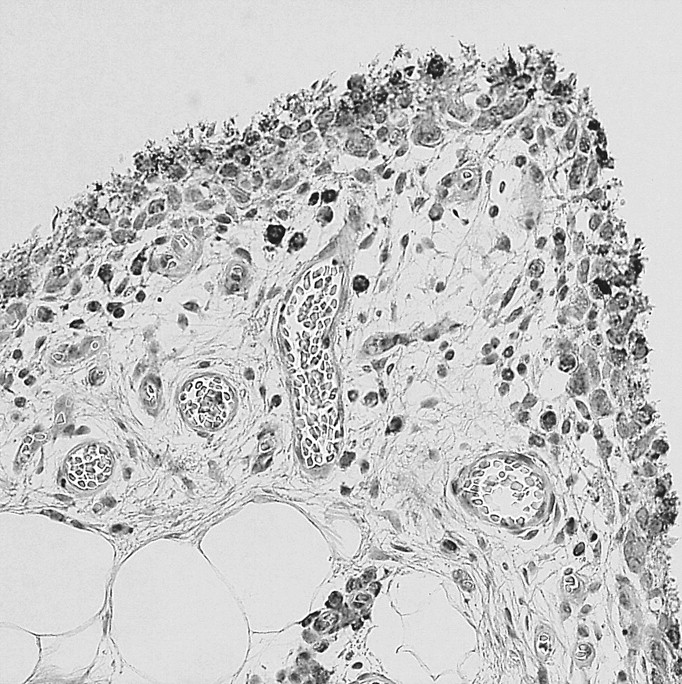
Synovial membrane samples showed positive labeling of OPN N-half in synovial lining cells and subintimal tissues in both RA and OA samples (Figure 4). The mean (± standard deviation) scores for synoviocytes in RA samples and OA samples were 2.0 ± 1.0 and 1.7 ± 0.7, respectively, and immunostaining for OPN N-half in synoviocytes showed no difference between RA and OA samples (p = 0.539). However, expression of OPN N-half was stronger in RA subintimal tissues (2.0 ± 0.6) compared with OA subintimal tissues (1.4 ± 0.5; p = 0.019).


Expression of thrombin-cleaved OPN in synovium. Synovial lining and sublining cells stained for OPN in RA (A) and OA (B). Strong OPN staining was found in RA synovium (original magnification ×200)
DISCUSSION
OPN could be involved in arthritis, which is induced by many proinflammatory cytokines, including IL-1β, IL-6, and TNF-α, and this protein is produced by macrophages in response to the activation by inflammatory stimuli12. Among its multiple properties, OPN, an extracellular matrix glycoprotein, has been shown to act as a T-helper type 1 immunoregulatory cytokine, and has been implicated in inflammatory responses through recruitment of inflammatory cells and augmentation of cytokine expression, including TNF-α and integrins18. OPN plays a pivotal role in the pathogenesis of RA in the mouse model23. Compared with arthritic wild-type mice, OPN-deficient mice are resistant to type II collagen-induced arthritis, showing attenuated joint swelling and cartilage destruction, chondrocyte apoptosis, and synovial angiogenesis11. Administration of an antibody directed against the SLAYGLR sequence, exposed by thrombin cleavage of murine OPN, has been shown to inhibit synovitis, bone erosion, and inflammatory cell infiltration in arthritic joints of animal models18.
The microvascular network of the RA synovium has an increased permeability to plasma proteins, and morphological abnormalities of the microvasculature are well described24,25. Inflammatory changes in vessel permeability and bleeding from fragile capillaries in the hyperplastic synovium may allow plasma thrombin to enter the joint space. Vascular endothelial growth factor (VEGF) induces mRNA expression encoding αvβ3 integrin subunits and OPN, an αvβ3 ligand, in dermal microvascular endothelial cells26. Concomitant induction of both the receptor and ligand promotes endothelial cell migration. OPN N-half may be involved in functional changes of microvasculature and inflammatory cell infiltration. Gattorno, et al27 reported that a positive correlation between OPN and VEGF concentrations was found in synovial fluid in juvenile idiopathic arthritis. In addition, OPN expression in the lining layer correlated with the number of vessels present in the areas underlying the sublining layer27. We need to study morphometric analysis of vessel walls and their expression of VEGF and cellular infiltration around the vessels. In addition, OPN is an autocrine/paracrine migratory and adhesive factor for tissue macrophages28. Macrophages in the synovium show expression of OPN9. OPN and VEGF are often expressed in close proximity to each other during angiogenesis26,29. Synovial macrophage-like cells produce thrombin as well as the components of extrinsic coagulation cascade in situ, promoting activation of coagulation pathways at biologically meaningful rates. Through its mitogenic effects, thrombin may elicit a cellular healing response inappropriate to the chronic inflammatory condition of the rheumatoid joints. It is capable of both causing the proliferation of synoviocytes and stimulating angiogenesis17,30. A study has revealed that thrombin concentration was higher in synovial fluid of patients with RA than in patients with OA31. Thrombin receptor-positive cells were present in rheumatoid synovia, but osteoarthritic and normal synovia contained few cells expressing thrombin receptors32.
Results of our present immunohistochemical study revealed that thrombin-cleaved OPN is highly expressed in the rheumatoid synovium at both synovial lining cells and subintimal tissues. Petrow, et al9 reported a similar expression pattern of intact OPN and mRNA by immunohistochemistry using monospecific affinity-purified rabbit antibody against human OPN and in situ hybridization, whereas Ohshima, et al33 found that the expression was predominantly in synovial lining cells using a mouse monoclonal IgG anti-human OPN antibody, 4C1. Petrow, et al9 showed no significant differences of OPN levels in synovial fluid between patients with RA and those with OA using a human OPN enzyme immunometric assay kit (Assay Designs, Ann Arbor, MI, USA). In contrast, Ohshima, et al33 reported that non-thrombin-cleaved synovial fluid OPN levels of RA patients were significantly higher than those of OA patients. They used 2 distinct sandwich ELISA systems, which can detect only non-thrombin-cleaved OPN and both nonthrombin- cleaved and thrombin-cleaved OPN. For detecting only non-thrombin-cleaved OPN, a rabbit polyclonal antihuman OPN antibody, OPN 1, was used for a coating antibody and a mouse monoclonal anti-human OPN antibody, OPN 3, was used for a detecting antibody. For detecting both non-thrombin-cleaved and thrombin-cleaved OPN, another rabbit polyclonal anti-human OPN antibody, OPN 5, was used for a coating antibody, and OPN 1 was used for a detecting antibody generated by their laboratory34. One possible reason for the difference in these results may be the molecular fragility of OPN. OPN can easily be cleaved into 2 fragments by thrombin, because the thrombin cleavage site is present in the OPN molecule. We used a novel ELISA system that we developed21; the system can measure only thrombin-cleaved OPN directly.
Limitations of our study include the cross-sectional design of the trial and the small RA patient sample, and lack of analysis of serum levels of thrombin-cleaved OPN. However, to our knowledge, this is the first report to measure the synovial fluid levels of thrombin-cleaved OPN in RA directly, and we have demonstrated that levels of thrombin-cleaved OPN are elevated in RA compared with OA. A recent study demonstrated that neutralizing antibody against the cryptic epitope of OPN, which is exposed by thrombin cleavage, could be a future therapeutic choice for patients with RA16.
Our study showed that local generation of thrombin-cleaved OPN was increased during rheumatic disease, indicating that the cleaved form may be a useful biochemical marker of RA. Further investigations are needed regarding the relationship between synovial fluid levels of thrombin-cleaved OPN levels and serum levels of thrombin-cleaved OPN, as well as progression of RA.
Acknowledgments
The authors thank Dr. N. Kinoshita for contributing to the study design.
Footnotes
- Accepted for publication September 30, 2008.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}