

Abstract
Objective. STAT3 and 4 are, among other factors, critical for the interleukin 12 (IL-12)-mediated Th1 response, for transfer of IL-23 signals, and for survival and expansion of Th17 cells. We investigated the association of STAT3 and STAT4 polymorphisms with serologically distinct subgroups of rheumatoid arthritis (RA).
Methods. A total of 41 single-nucleotide polymorphisms (SNP) within STAT3 and STAT1-STAT4 loci were investigated in a Swedish cohort of 2043 RA cases and 1115 controls. Nine of the associated SNP were tested in a Spanish cohort of 1223 RA cases and 1090 controls.
Results. Fourteen SNP in the STAT3 and STAT1-STAT4 loci were associated with anticitrullinated protein antibody (ACPA)-negative RA in the Swedish cohort. Three of the SNP in STAT4 and 2 SNP in STAT3 remained associated with ACPA-negative RA after considering the Spanish results. In addition, rs7574865 and rs10181656, in STAT4, were associated with ACPA-positive RA in the Swedish study. One of these SNP, rs7574865, showed a similar pattern of the association in serologically distinct subgroups of RA in a metaanalysis of all 7 published studies.
Conclusion. Our findings suggest that variants in STAT genes may contribute differentially to susceptibility to RA in seropositive and in seronegative patients.
Rheumatoid arthritis (RA) is an autoimmune disease in which genetic and environmental factors are involved1. It can be divided into 2 clinically relevant subgroups by the presence of autoantibodies to citrullinated proteins/peptides (ACPA). ACPA-positive and ACPA-negative RA are different in severity, with ACPA-negative RA generally being a milder disease. Most of the genetic variants associated with RA are specific to ACPA-positive RA [e.g., HLA-DRB1 shared epitope (SE) alleles, PTPN22, C5/TRAF-1] and only a few have no such discrimination or are restricted to ACPA-negative RA2,3,4. Especially striking is the case of HLA-DRB1 SE alleles, which notably increase the risk for autoantibody-positive RA in most European white populations5, but have no effect or a very modest one for autoantibody-negative RA. This finding is consistent with the idea that development of autoantibodies, in RA and in other autoimmune diseases, results from a loss of tolerance, which is tightly connected to T cell function and MHC class II specificity. However, the details of these mechanisms remain unclear. Specifically, it is not clear how similar clinical phenotypes may arise in serologically distinct subgroups of RA and with different HLA background.
STAT3 and STAT4 are important transcription factors for interleukin 12R (IL)-12R/IL23 receptor signaling and for mediating signals during Th1 and Th17 cell differentiation, among other pathways and cell functions. STAT4 has been shown to be associated with several autoimmune diseases with significant production of autoantibody, such as RA and systemic lupus erythematosus (SLE)6,7. However, there is no consistency from genetic association studies whether STAT4 is associated with ACPA-positive or negative RA8,9,10,11,12. STAT3 variants are associated with susceptibility to Crohn disease13, multiple sclerosis14, and with ankylosing spondylitis15 (all autoimmune diseases without much influence on autoantibody production), but not with RA. We investigated whether there is an association between STAT3 and STAT4 genetic variants and ACPA-positive or ACPA-negative RA.
MATERIALS AND METHODS
Study population
The Epidemiological Investigation of Rheumatoid Arthritis (EIRA) is a population-based case-control study of incident cases of RA. Controls were randomly selected from the Swedish national population registry and were matched to the patients for age, sex, and residential area, as described16. In our current study, 2418 patients (71.2% women) and 1393 controls (74% women) recruited between 1996 and 2007 served as a discovery cohort. The median age of the patients was 51 years (range 39–63 yrs) and 64% of the patients were ACPA-positive.
For the Spanish study, recruitment of 1223 patients (76% women) and 1090 controls (58.2% women, among 1010 individuals with available information) was done from 2001 to 2010 in 13 Spanish hospitals, as described17. All samples from hospitals in previous studies were excluded from metaanalysis to avoid duplication of data. The median age of patients was 49 years (range 38–58 yrs) and 68.1% were ACPA-positive (ACPA status available for 1125 patients). Patients from both studies fulfilled the American College of Rheumatology 1987 criteria18. The studies were done with approval from the regional ethical committees of the Stockholm area and of Galicia and informed consent was obtained from all participants.
Measurement of ACPA levels
The level of ACPA for samples included in the EIRA study was determined using Immunoscan-RA Mark2 ELISA (Euro-Diagnostica, Arnhem, The Netherlands) as described19. Samples with antibody levels above 25 U/ml were regarded as positive according to instructions provided by the manufacturer. For the Spanish study, ACPA levels were determined using the EDIA ACPA Kit (Euro-Diagnostica). The cutoff level was 5 U/ml, following manufacturer’s instructions.
Genotyping
The majority of patients and controls in the EIRA study population (2043 patients with RA and 1115 matched controls) have been genotyped for > 300,000 single-nucleotide polymorphisms (SNP) using the Illumina HumanHap300 array (Illumina Inc., San Diego, CA, USA) and the details have been published20. Available genotyping data for STAT3 and STAT4 were extracted from this genome-wide association study (GWAS). We also included an additional 50 kb in the upstream region of the STAT4 gene to investigate the STAT1 gene. In the GWAS, 34 SNP were available for STAT3 and STAT1-STAT4 loci (Figure 1A). Next, a fine-mapping analysis (stage 1) of STAT3 and STAT4 with 37 SNP (not included in the GWAS) was performed on the part of the EIRA study consisting of samples from 451 patients with RA and 269 healthy matched controls (Figure 1B). These 37 SNP were selected for genotyping because they were in close proximity to the genes of interest and had a minor allele frequency in the white population ≥ 5%. The results were combined with GWAS data for more accurate definition of blocks of linkage disequilibrium (LD). This information was used to select 7 additional SNP not available in the GWAS and not included in stage 1 for a second round of fine-mapping (stage 2) in the whole EIRA material (2418 patients, 1374 controls; Figure 1C). Genotyping was done using iPLEX Gold chemistry of Sequenom’s MassArray platform (San Diego, CA, USA) for stage 1 and TaqMan allelic discrimination assays (Applied Biosystems, Foster City, CA, USA) for stage 2. The genotyping call rate was above 97% for all SNP at stage 2. The genotyping data from fine-mapping stage 2 were analyzed together with GWAS data.

Various steps of genotyping of the STAT1-STAT4 and STAT3 loci. A. Thirty-four SNP genotyped in the Swedish cohort were extracted from a genome-wide association study (GWAS). B. Thirty-seven additional SNP were genotyped on a fraction of the Swedish cohort (fine-mapping stage 1) and these results were combined with GWAS data and used as a guideline in the selection of 7 SNP for the second round of fine-mapping (C; stage 2). The results from fine-mapping stage 2 were combined with GWAS data and 9 tag SNP were selected and genotyped in the Spanish cohort (D). LD: linkage disequilibrium.
Out of 14 SNP associated with ACPA-negative RA (690 patients, 1076 controls) in the EIRA study, as a result of analyses of stage 2 data combined with the GWAS, 9 tag SNP were selected for evaluation in the Spanish study (Table 1, Figure 1D). Polymerase chain reaction (PCR) amplification was done with the KAPA2G Fast HotStart kit (Kapa Biosystems, Woburn, MA, USA) and PCR products were purified by Exo-SAP digestion with Exonuclease I (Epicentre, Madison, WI, USA) and shrimp alkaline phosphatase (GE Healthcare, Barcelona, Spain). Subsequently, single-base extension reactions with the SNaPshot Multiplex kit (Applied Biosystems) were done following the manufacturer’s instructions, and the genotyping call rate was 99.9%.
Associations in Swedish and Spanish rheumatoid arthritis cohorts that were negative for autoantibodies to citrullinated proteins/peptides.
Statistical analysis
Association analyses for single marker and haplotypes were performed using the HaploView 4.0 software. The LD between the SNP was estimated using D’ and r2. Frequency distribution of all SNP genotyped in the Swedish cohort and the Spanish cohort was in agreement with Hardy-Weinberg equilibrium (p > 0.05). After combining GWAS and fine-mapping stage 2 genotyping data, individuals with < 99% successful genotypes for all SNP were removed from analysis. The chi-square test was used to compare allele frequencies between patients and controls, and OR with 95% CI were calculated. Bonferroni correction for multiple comparisons was used, based on the number of SNP included in the analysis and the number of groups (ACPA-positive and ACPA-negative). The difference in the number of controls from GWAS and the number of controls in Table 1 for the Swedish cohort (1115 and 1076, respectively) is due to removing 39 individuals from the analysis as genetic outliers based on principal components analysis with SNP data from the Illumina 300K chip4. In the final analysis (Table 1) the power of the Spanish study to detect an effect with an OR of 1.49 was 74% for the SNP with the lowest minor allele frequency (rs3024935).
Metaanalysis of current results was done using the Mantel-Haenszel method for combining OR under a fixed model of all the STAT SNP showing significant association with RA in the EIRA study and investigated in the Spanish study (n = 9). All SNP passed the test for homogeneity of effects in a recessive model except for rs6751855, which passed the test for homogeneity in a dominant model (p > 0.05).
Literature search (PubMed and ISI Web of Knowledge) was performed with the terms “rheumatoid arthritis,” “STAT4,” and “polymorphism.” Studies with a white study population and available ACPA status and genotype frequencies for rs7574865 (the most commonly investigated SNP in STAT4) were selected. Five publications were identified for ACPA-negative RA and 6 for ACPA-positive RA (the most recent article from January 2011)8,9,10,11,12,21. The following information was extracted from each report: ethnicity, ACPA status, allele frequencies for the T-allele of rs7574865, and the number of cases and controls. The 2 cohorts from the current study were also included in the metaanalysis (Figure 2A, 2B).
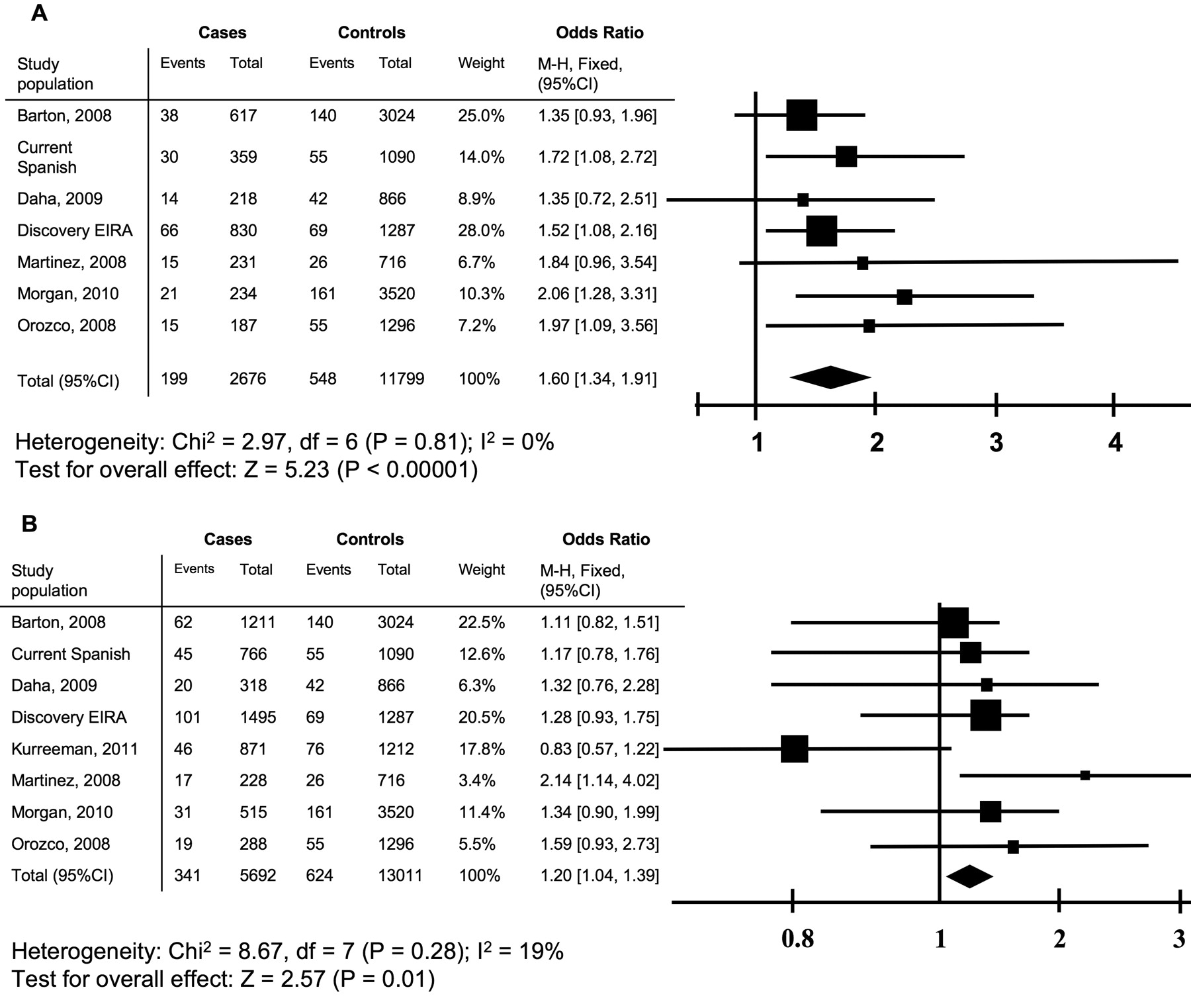
Metaanalysis of case control studies for risk of RA measured by OR for rs7574865 (STAT4). A combined OR was calculated in a recessive model among (A) patients negative for autoantibodies to citrullinated proteins/peptides (ACPA; n = 2676) relative to controls (n = 11799), and (B) in ACPA-positive patients (n = 5692) relative to controls (n = 13011). For each study, OR is represented by a square and the combined estimate is presented as a diamond. M-H: Mantel-Haenszel; EIRA: Epidemiological Investigation of Rheumatoid Arthritis study.
RESULTS
Association in the STAT1-STAT4 locus
We performed association analyses for 210 kbp of the STAT1-STAT4 locus at chromosome 2q32.2 with SNP selected from the EIRA GWAS (n = 29) and with genotype data from de novo genotyping done in stage 2 (n = 5). The analyses were done separately in ACPA-positive and ACPA-negative patients with RA. Five SNP located in the STAT4 (rs11889341, rs7574865, rs8179673, rs10181656 and rs3024935), 1 SNP in the intergenic region (rs6751855), and 2 SNP located in STAT1 (rs10173099, rs12693591) were associated with ACPA-negative RA (Table 1) as a result of combining data from GWAS and stage 2 (Figure 3A). rs12693591 remained significant after Bonferroni correction for multiple testing. Additionally, SNP rs7574865 (OR 1.13, 95% CI 1.0–1.29) and rs10181656 (OR 1.15, 95% CI 1.02–1.31) in STAT4 were also associated with ACPA-positive RA using an allelic model (Table 2). However, these latter differences in allelic frequency were nonsignificant after correction for multiple testing. Haplotype-based association analysis did not reveal any additional information (data not shown).
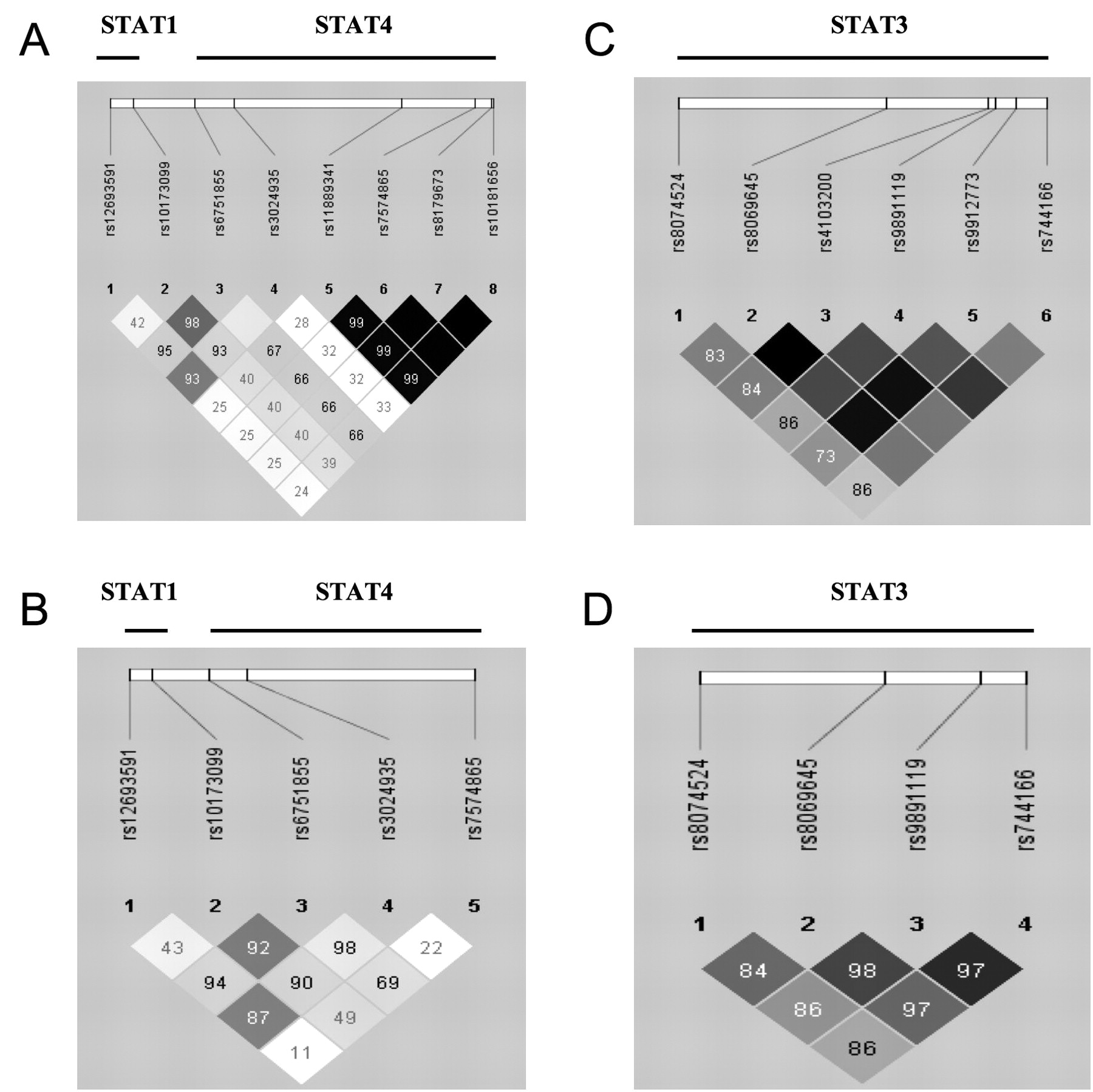
Linkage disequilibrium map showing D-prime values and r2 for the genotyped SNP in STAT1/STAT4 loci (A, B) and STAT3 loci (C, D) in the Swedish cohort (A, C) and in the Spanish cohort (B, D). The figure was generated using HaploView 4.0 software.
Associations in Swedish and Spanish rheumatoid arthritis cohorts that were positive for autoantibodies to citrullinated protein/peptides.
Five of the associated SNP in the EIRA samples were tested in an independent Spanish cohort of RA cases and controls for replication (Figure 3B). Only rs7574865 showed association with ACPA-negative RA in this population (Table 1). The same SNP showed a trend toward association with ACPA-positive RA (p = 0.073). Two of the other 4 SNP, rs12693591 and rs6751855, were still significantly associated with ACPA-negative RA in the combined analysis of the EIRA and the Spanish samples. No association with ACPA-positive RA was detected in the Spanish samples with any of the SNP in this locus (Table 2).
We performed Mantel-Haenszel metaanalysis to summarize all information available for the STAT4 SNP rs757865 including the results from our current study and previous reports8,9,10,11,12,21. In total, 2676 and 5692 ACPA-negative and ACPA-positive patients with RA were included in each metaanalysis, respectively. The results demonstrated association with both subgroups of patients with RA (p < 0.00001, p = 0.01 for ACPA-negative and ACPA-positive patients, respectively). The strength of association was similar, as shown by the overlapping 95% CI of the OR, although there was a trend to a stronger association with the ACPA-negative patients than with the ACPA-positive ones (pooled OR 1.60, 95% CI 1.34–1.91, and OR 1.20, 95% CI 1.04–1.39, respectively; Figure 2A, 2B).
Association at the STAT3 locus
Our investigation of the STAT3 locus at chromosome 17q21.31 in the EIRA samples was based on 7 SNP with genotypes obtained either from our previous GWAS (n = 5) or from fine-mapping stage 2 (n = 2): rs9891119, rs744166, rs8069645, rs9912773, rs8074524, rs4103200, and rs17405722. This analysis resulted in relatively robust association hits for all tested variants (Figure 3C), except for rs17405722, with ACPA-negative RA. Three of the associated SNP remained significant after correction for multiple testing (Table 1). Notably, we did not find association of these genetic variants with ACPA-positive RA (Table 2).
To replicate our findings, 4 of the associated STAT3 SNP (rs9891119, rs8069645, rs744166, and rs8074524) were genotyped in the Spanish cohort (Figure 3D). None of the tested SNP demonstrated significant association in the replication cohort. However, the difference between cases and controls for 2 SNP, rs9891119 and rs744166, was coherent with what we have found in the EIRA cohort, and metaanalyses demonstrated significant association for the combined results in the absence of heterogeneity (OR 1.29, 95% CI 1.05–1.60, and OR 1.35, 95% CI 1.12–1.63, respectively; Table 1). No association with ACPA-positive RA was detected in the replication cohort for the STAT3 locus (Table 2) and haplotype analysis did not add further information to the single-marker analysis (data not shown).
DISCUSSION
Introduction of ACPA assays for detection of ACPA in individuals with RA contributed greatly to our understanding of the disease. The majority of the RA-associated loci found to date are specific for ACPA-positive RA (e.g., PTPN22, C5/TRAF-1). Only a few loci have shown predominant association with ACPA-negative RA2,22,23. We found that STAT3 and STAT4 are 2 additional loci associated with ACPA-negative RA.
To our knowledge, this is the first time genetic variants in STAT3 have been investigated in RA, although this locus has been shown to be associated with Crohn disease13, multiple sclerosis14, and ankylosing spondylitis15. In our study, we found suggestive evidence for association of STAT3 with ACPA-negative RA in the EIRA study and in a combined analysis, but the association cannot be considered validated because it was not replicated in the Spanish cohort. Lack of replication could be ascribed to low statistical power because direction of differences between cases and controls was the same in the 2 sample collections.
STAT4 is a gene that has been investigated frequently for many complex diseases including multiple studies in RA6,7,24. The SNP that has been more thoroughly studied is rs7574865. However, previous reports regarding association of rs7574865 with the ACPA subgroups of RA are contradictory6,7,8,9,10,11,12. Because of these discrepancies, we performed metaanalysis on our results with publications where the association of rs75748645 with ACPA subgroups of RA has been investigated in whites. Three additional white study populations are included in this metaanalysis, compared with 2 previously published metaanalyses for this SNP25,26, which gives us higher power and increased confidence in the summary results.
Further, previous metaanalysis demonstrated that the observed effect was similar for rs7574865 in ACPA-positive and ACPA-negative RA, whereas in our current study we found that there is an indication of stronger effect size of the STAT4 SNP rs7574865 in the ACPA-negative RA group compared to the ACPA-positive group. This difference in effect might possibly reflect the heterogeneity of RA. However, even using metaanalysis and a high number of cases, we did not reach a statistically significant difference between effects of 2 subgroups of disease, an issue that should be explored further.
Similar to rs7574865, 1 other SNP (rs10181656) was associated with both subgroups of RA. This SNP is in the same LD block as rs7574865 (D’ = 1; r2 = 0.99) and has been reported to be in association with SLE27, early-onset type 1 diabetes28, and RA6.
Some other SNP in the STAT1-STAT4 locus that we investigated were exclusively associated with ACPA-negative RA. Two of the SNP, in the intergenic region and in STAT1, were significantly associated with ACPA-negative RA in the combined analysis of the Swedish and Spanish cohorts. Therefore, these results raise the possibility that there are SNP in this locus associated with the generality of the patients with RA, such as rs7574865 and SNP in tight LD with it, and other SNP associated only with ACPA-negative RA, such as rs12693591 and rs6751855. However, these associations need to be confirmed in additional studies because they were found only in the EIRA samples.
We found that STAT4 rs7574865 is associated with both ACPA-positive and ACPA-negative RA, with an indication of stronger effect size in ACPA-negative RA, whereas other SNP in the STAT1-STAT4 locus and in STAT3 show association only with ACPA-negative RA. Therefore, genetic variation in these genes that are from the same lymphoid signaling pathway could contribute in different ways to RA pathogenesis. It is to be expected that mechanistic studies will reveal the specific details of these complex associations. However, independent confirmation of our results regarding some of the associations is still required.
Acknowledgment
We thank the patients for their contribution to the study, and Eva Jemseby, Gull-Britt Almgren, and Julia Boström for the administration of biomaterial. Members of the Rheumatoid Arthritis Network and Coordinated Project contributing samples, data, or help in analysis to the Spanish study include Eva Perez-Pampin and Manuel Calaza from Hospital Clinico Universitario de Santiago de Compostela; Arturo Rodriguez de la Senra from Hospital Santa Creu e San Pau, Barcelona; Luis Carreño from Hospital Gregorio Marañon, Madrid; Juan Cañete from Hospital Clinic, Barcelona; Benjamin Fernandez-Gutierrez from Hospital Clinico San Carlos, Madrid; Javier Narvaez from Hospital de Bellvitge, Barcelona; Beatriz Joven from Hospital 12 de Octubre, Madrid; Alejandro Balsa from Hospital La Paz, Madrid; Jose Luis Marenco from Hospital Valme, Seville; and Ana Ortiz from Hospital de la Princesa, Madrid.
Footnotes
-
Supported by the Combine project from Swedish Vinnova. The Rheumatoid Arthritis Network and Coordinated Project was established with support from the Instituto de Salud Carlos III (ISCIII, Spain) grants G03/152 and PI041513. Genotyping of the Network samples was funded by grant PI080744 from the ISCIII to Dr. Gonzalez.
- Accepted for publication May 7, 2012.








{kind=link}
{kind=link}
{kind=link}