Article Text

Abstract
Background: In coeliac disease (CD), the upper bowel lesion is associated with a marked infiltration of the mucosa with Th1 cells secreting interferon γ (IFNγ) and expressing the Th1-associated transcription factor, T-bet. However, the molecular mechanisms which regulate T-bet and promote the Th1 cell response are unknown.
Objective: To examine whether interleukin 21 (IL21), a cytokine that regulates T cell activation, has a role in CD.
Methods: Duodenal mucosal samples were taken from CD patients and normal controls. IL21 and T-bet were examined by real-time PCR and western blotting, and IFNγ was assessed by real-time PCR and ELISA. The effect of blockade of endogenous IL21 on the expression of T-bet was examined in an ex vivo culture of biopsies taken from untreated CD patients. Finally, the role of IL21 in controlling T-bet and IFNγ was also evaluated in cultures of biopsies taken from treated CD patients and cultured with a peptic–tryptic digest of gliadin (PT) in the presence or absence of a neutralising IL21 antibody.
Results: Enhanced IL21 RNA and protein expression was seen in duodenal samples from untreated CD patients. Blockade of IL21 activity in biopsies of untreated CD patients reduced T-bet and IFNγ secretion. Stimulation of treated CD biopsies with PT enhanced IL21 expression, and neutralisation of IL21 largely prevented PT-driven T-bet and IFNγ induction.
Conclusions: IL21 is overproduced in the mucosa of CD patients, where it helps sustain T-bet expression and IFNγ production.
Statistics from Altmetric.com
In active coeliac disease (CD) the upper bowel lesion is associated with a marked accumulation of polarised Th1 cells, making large amounts of interferon γ (IFNγ).1 2 Neutralisation of endogenous IFNγ prevents the gliadin-mediated morphological changes in ex vivo organ culture of treated CD biopsies.3 These observations, together with the demonstration that activation of Th1 cells in fetal gut explants results in villous atrophy and crypt cell hyperplasia, suggest that Th1-type cytokines play an important role in the CD-associated mucosal damage.4 5 Nonetheless, it is not yet clear how Th1 cells are induced and maintained in the intestine of CD patients.
We have shown that T-bet, a member of the T-box family of transcription factors, critical to Th1 polarisation, is highly expressed in the inflamed gut of CD patients.6 Additionally, immunohistochemistry revealed expression of T-bet in mucosal CD4+ and CD8+ T cells in CD.7 Although, T-bet is rapidly induced when T cells recognise antigens in the presence of IFNγ, and we previously showed that IFNγ enhances T-bet expression in normal mucosal T cells,6 the exact molecular mechanisms that induce and stabilise T-bet in CD remain to be determined.
Interleukin 21 (IL21) is a recently identified cytokine produced by activated CD4+ T cells,8 and by natural killer (NK) T cells following in vitro stimulation via CD3 and CD28, or activation with α-galactosylceramide.9 IL21 biological functions are mediated by a heterodimeric receptor, formed of a unique subunit, designed IL21R, and the common γ-chain subunit, shared with IL2, IL4, IL7, IL9, IL13 and IL15.10–12 IL21R is expressed by both immune and non-immune cells, and ligand binding to IL21R activates the JAK family protein tyrosine kinases JAK1 and JAK3, leading to the activation of signal transducer and activator of transcription (Stat) 1, Stat3 and, to a lesser degree, Stat4, Stat5 and Stat6, depending on the cell types studied.13
Several studies have documented the ability of IL21 to regulate the production of cytokines by T cell subsets. However, conflicting results have been reported, depending on the cell type analysed. Studies in murine systems have shown that IL21 is mostly made by Th2 cells, and exposure of naïve T cells to IL21 results in a decreased production of IFNγ, linked to the repression of the expression of another T-box transcription factor, eomesodermin.14 In contrast, analysis of IL21 mRNA expression by human blood Th1 and Th2 cells showed IL21 only in Th1 cells.15 IL21 also enhances the expression of Stat4 and T-bet, and stimulates the production of IFNγ in human T cells.8 12 In agreement with this, we have recently shown that IL21 is overproduced in patients with Crohn’s disease,16 another Th1-associated gastrointestinal disease driven by IL12 and IL23. Moreover, blockade of IL21 activity in cultures of Crohn’s disease lamina propria mononuclear cells reduces the expression of p-Stat4 and T-bet, and the production of IFNγ,16 thus suggesting that IL21 may be part of a positive feedback loop that contributes to amplify Th1 cell responses.
We have therefore sought to determine the role of IL21 in CD. We first examined IL21 in the duodenal mucosa of patients with CD and controls. Secondly, using the ex vivo organ culture of duodenal explants, we examined whether IL21 positively controls the expression of T-bet and IFNγ.
METHODS
Patients and controls
Biopsies from the distal duodenum of 43 patients with untreated CD (aged from 22 to 54 years) were obtained during upper gastrointestinal endoscopy. The histopathological diagnosis was based on typical mucosal lesions with crypt cell hyperplasia, villous atrophy and increased number of intraepithelial lymphocytes (IELs). All untreated CD patients were positive for antiendomysial (EMA) and antitransglutaminase antibodies at the time of diagnosis. Duodenal biopsies were collected at the time of diagnosis from all these patients. Biopsies were also obtained from 12 treated CD patients (aged from 29 to 42 years), who were in clinical and histological remission, and negative for EMA and antitransglutaminase antibodies. Normal controls (n = 36) were under investigation for gastrointestinal symptoms, but had normal histology, no increase in inflammatory cells and were EMA and antitransglutaminase antibody negative. This study received ethical approval.
Organ culture
Duodenal biopsies were taken from untreated CD patients and cultured in RPMI 1640 supplemented with 10% fetal bovine serum (FBS), penicillin (100 U/ml) and streptomycin (100 μg/ml) (all from Sigma-Aldrich, Milan, Italy). Biopsies were either left untreated or treated with a neutralising monoclonal anti-human IL21,17 and/or a neutralising antihuman IL15 (R&D Systems, Minneapolis, Minnesota, USA) or control immunoglobulin G (IgG; final concentration: 10 μg/ml). The biopsies were placed on iron grids in the central well of an organ culture dish, and the dishes placed in a tight container with 95% O2/5% CO2 at 37°C, at 1 bar. Duodenal biopsies were also taken from treated CD patients and controls and cultured with a peptic–tryptic digest of gliadin (PT, 1 mg/ml) in the presence or absence of the anti-IL21 or control IgG as indicated above. After 24 h culture, tissue explants were used for examining IL21, T-bet and IFNγ, while organ culture supernatants were analysed for the content of IFNγ by ELISA.
Lamina propria mononuclear cell (LPMC) isolation
LPMCs were isolated by duodenal biopsies taken from three patients with active CD and three normal controls as previously described,16 and used to purify CD4-positive T lamina propria lymphocytes (T-LPLs). For this purpose, LPMCs were incubated for 30 min at 4°C with magnetically labelled CD4 antibodies (Miltenyi Biotec, Calderara di Reno, Italy) and both CD4+ and CD4– T-LPLs were then collected using the magnetic cell sorting system (MACS, Miltenyi Biotec S.r.l.).
Protein extraction and western blotting
IL21 was analysed using total proteins extracted from biopsies of 14 untreated CD patients, 3 treated CD patients and 14 controls. IL21 was also analysed in duodenal biopsies taken from treated CD patients and controls, and cultured as indicated above. Samples were lysed for 60 min on ice in buffer containing 10 mM HEPES (pH 7.9), 10 mM KCl, 0.1 mM EDTA, 0.2 mM EGTA and 0.5% Nonidet P40, supplemented with 1 mM dithiothreitol (DTT), 10 μg/ml aprotinin, 10 μg/ml leupeptin, 1 mM phenylmethylsulfonyl fluoride (PMSF), 1 mM Na3VO4 and 1 mM NaF (all reagents were from Sigma-Aldrich). Lysates were then clarified by centrifugation at 4°C for 30 min at 12 000 g. Equal amounts of total proteins were fractionated on SDS–polyacrylamide gels. The membranes were blocked with Tris-buffered saline containing 0.05% Tween-20 and 5% non-fat dry milk. IL21, T-bet and β-actin were evaluated as previously described.6 16 Computer-assisted scanning densitometry (Total lab, AB.EL, Rome, Italy) was used to analyse the intensity of the immunoreactive bands.
RNA extraction, cDNA preparation and real-time PCR
Analysis of IL21 transcripts was performed by real-time PCR using total RNA extracted from duodenal biopsies of 12 untreated CD patients and 12 normal controls. RNA samples extracted from the same 12 untreated CD patients were also evaluated for T-bet in order to ascertain if the expression of these two molecules is correlated in CD. IL21 was also evaluated in purified CD4+ and CD4– T-LPLs. RNA was extracted by using TRIzol reagent according to the manufacturer’s instructions (Invitrogen, Milan, Italy). A constant amount of RNA (1 μg/sample) was retro-transcribed into cDNA, and 1 μl of cDNA/sample was then amplified using the following conditions: denaturation 1 min at 95°C, annealing 30 s at 60°C for both IL21 and T-bet, and at 62°C for β-actin, followed by 30 s of extension at 72°C. IL21 and T-bet were evaluated using commercially available TaqMan probes (Applied Biosystems, Foster City, California, USA). Real-time PCR was performed using the IQ SYBR Green Supermix (Bio-Rad Laboratories, Milan, Italy). β-actin (FWD: 5′-AAGATGACCCAGATCATGTTTGAGACC-3′ and REV: 5′-AGCCAGTCCAGACGCAGGAT-3) was used as an internal control.
Determination of IFNγ by ELISA
IFNγ was evaluated in supernatants of organ cultures of duodenal biopsies using a sensitive ELISA kit (R&D Systems), and values were expressed as pg/μg of total proteins.
Statistical analysis
Differences between groups were compared using the Student t test, ANOVA and Wilcoxon tests.
RESULTS
IL21 expression is increased in untreated CD mucosa
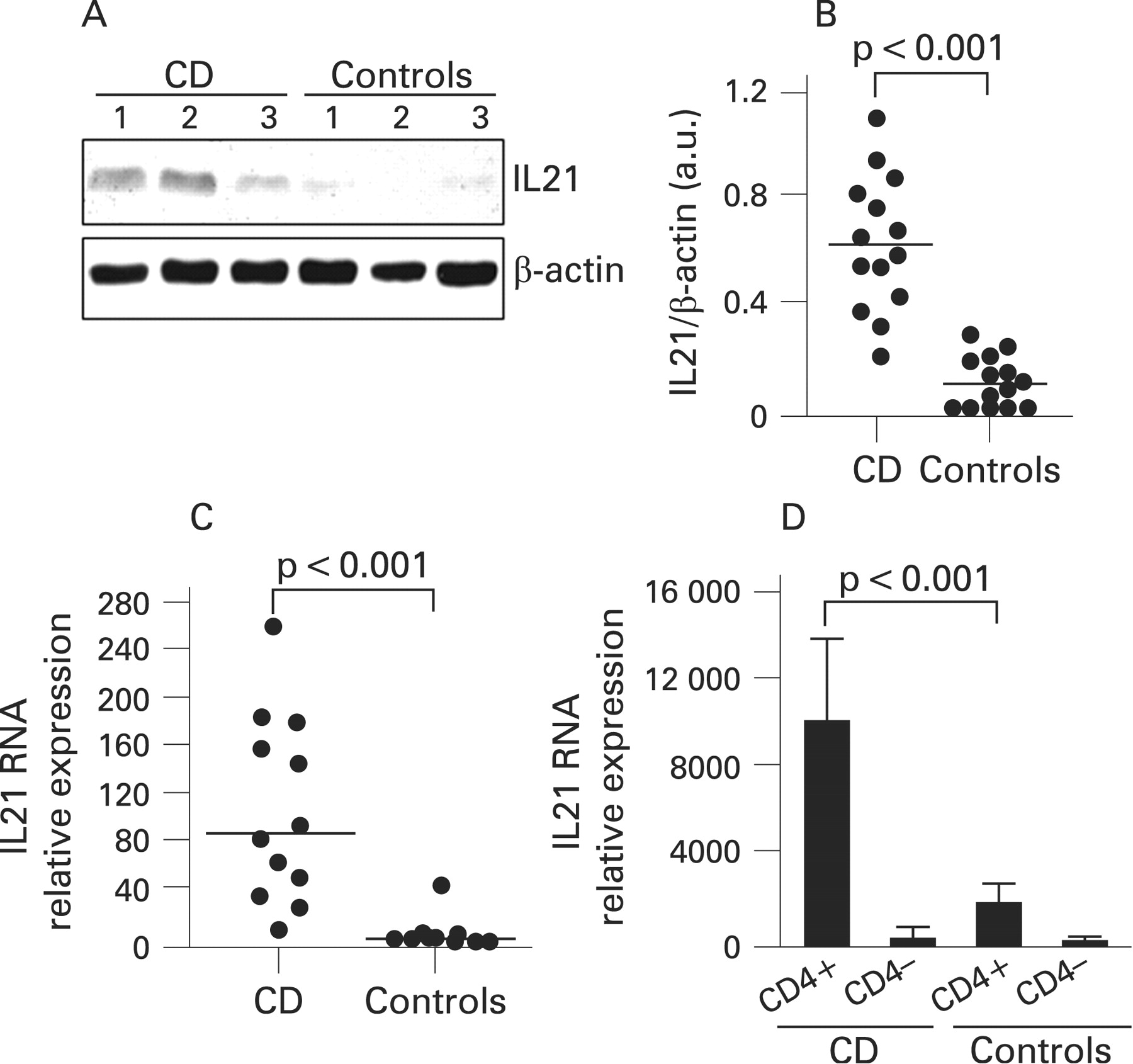
Immunoreactivity for IL21 protein was seen in all untreated CD patients and 9/14 control samples. There was a clear increase in the intensity of the IL21 bands in CD compared with controls (fig 1A). IL21 expression quantitated by densitometry and normalised by β-actin expression was significantly increased in biopsies taken from untreated CD patients as compared with controls (fig 1B, p<0.001). Biopsies taken from treated CD patients showed the same levels of IL21 as controls (not shown).
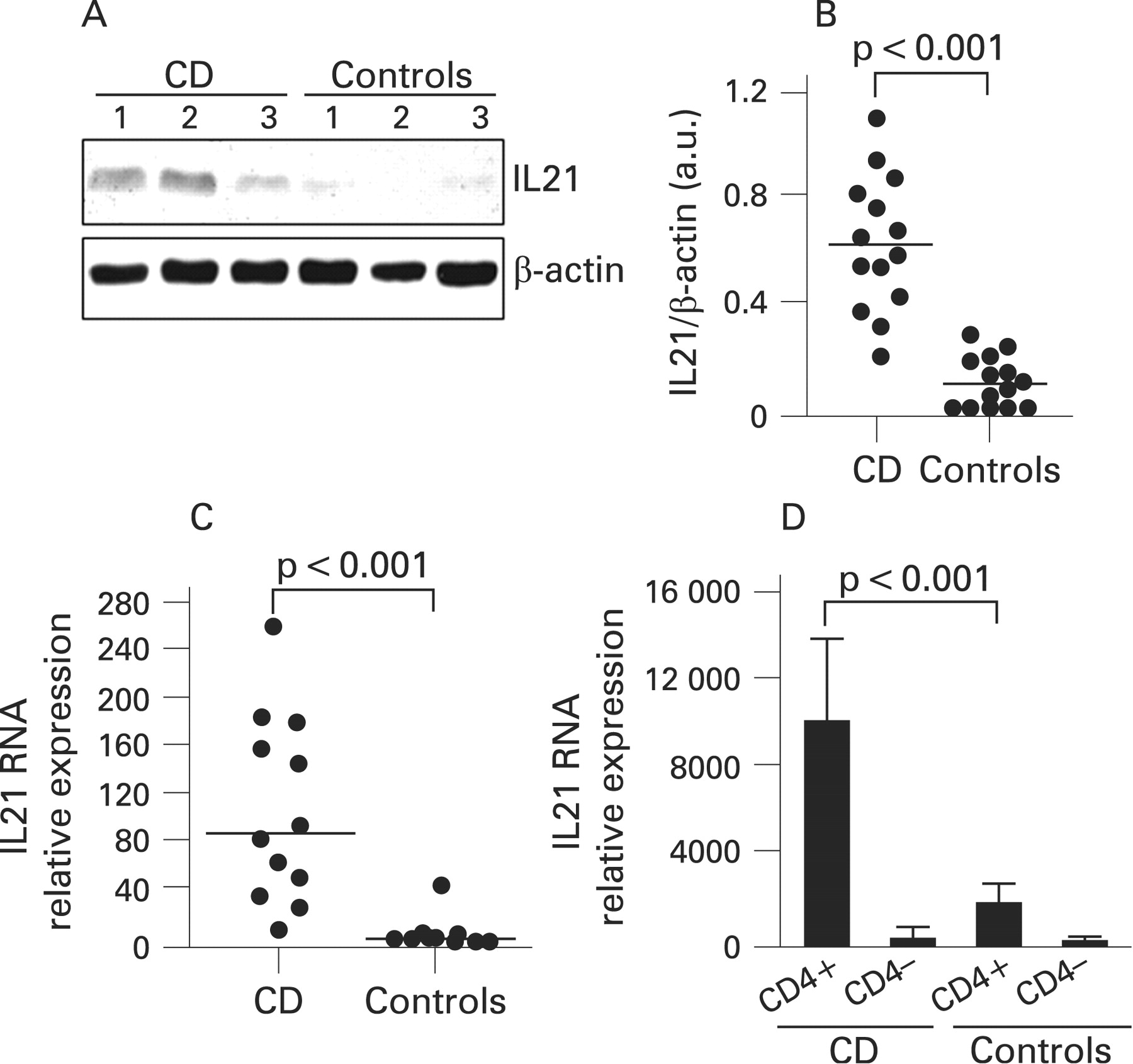
Mucosal samples from untreated CD patients and controls were assessed for IL21 RNA transcripts by real-time PCR. Figure 1C shows that IL21 RNA expression was significantly higher in CD patients than in controls (p<0.001). In order to exclude that differences reflected the higher number of mucosal T lymphocytes in CD, IL21 RNA expression was also assessed in CD4+ and CD4–T-LPLs isolated from CD and normal biopsies. As shown in fig 1D, CD4+ T-LPLs from CD patients contained a higher amount of IL21 RNA than that seen in cells isolated from controls. In contrast, IL21 was barely detectable in CD4– T-LPLs from CD patients and controls (Fig 1D).
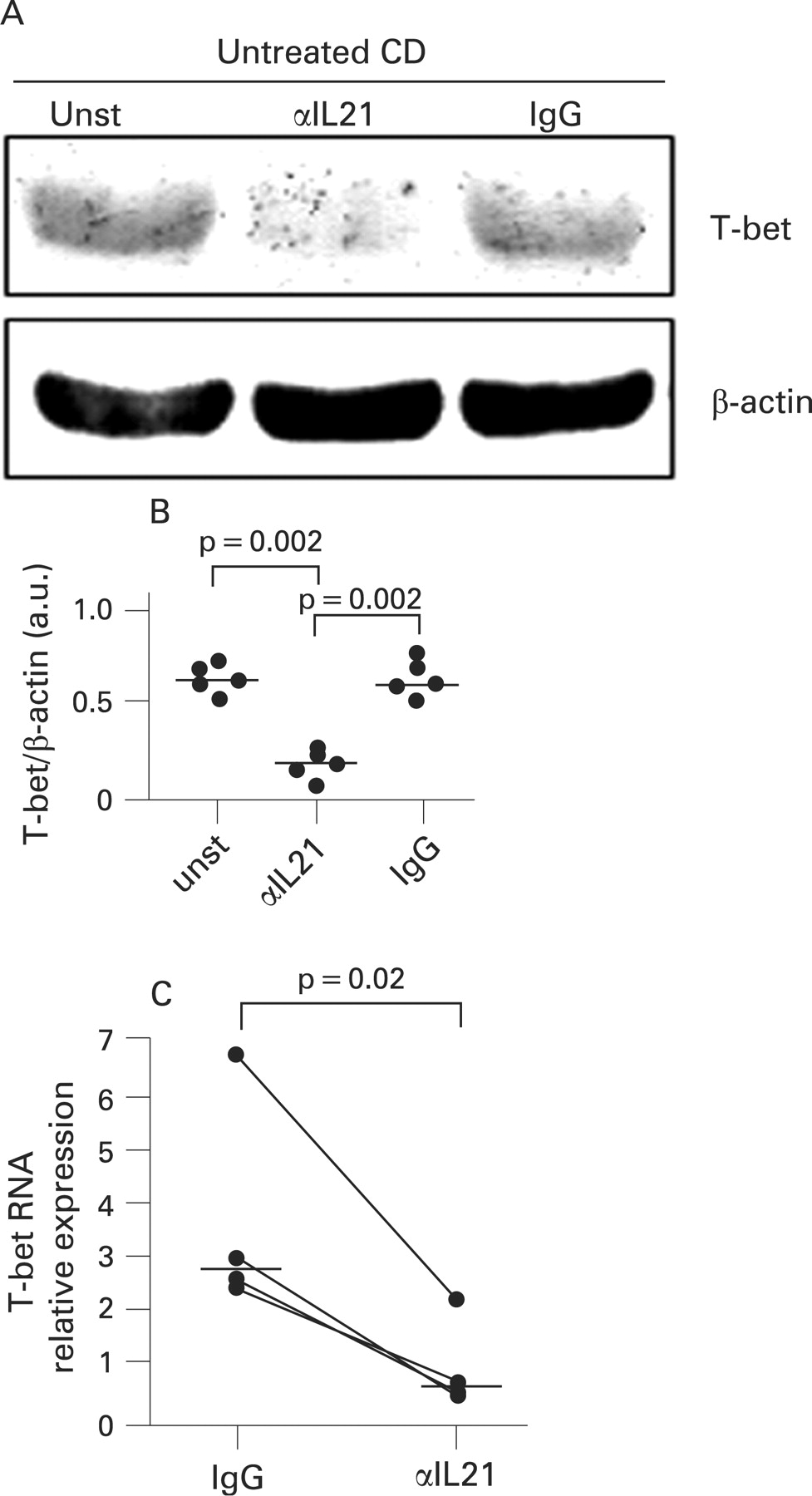
Blockade of IL21 results in a reduced expression of T-bet and IFNγ in organ cultures of duodenal explants of untreated CD patients
We next carried out ex vivo organ cultures and examined whether blocking endogenous IL21 reduced T-bet in duodenal biopsies of active CD patients. Duodenal biopsies taken from untreated CD patients were cultured with or without a neutralising anti-IL21 antibody or control antibody for 24 h and then analysed for T-bet by western blotting. T-bet was clearly detectable in biopsies cultured with medium alone, and was downregulated by anti-IL21 but not by the control antibody (fig 2A). Densitometric analysis of western blots confirmed that blockade of endogenous IL21 resulted in a significant inhibition of T-bet expression (fig 2B, p = 0.002). Treatment of biopsies from untreated CD patients with the anti-IL21 antibody also resulted in a significant inhibition of T-bet RNA expression (fig 2C, p = 0.01).
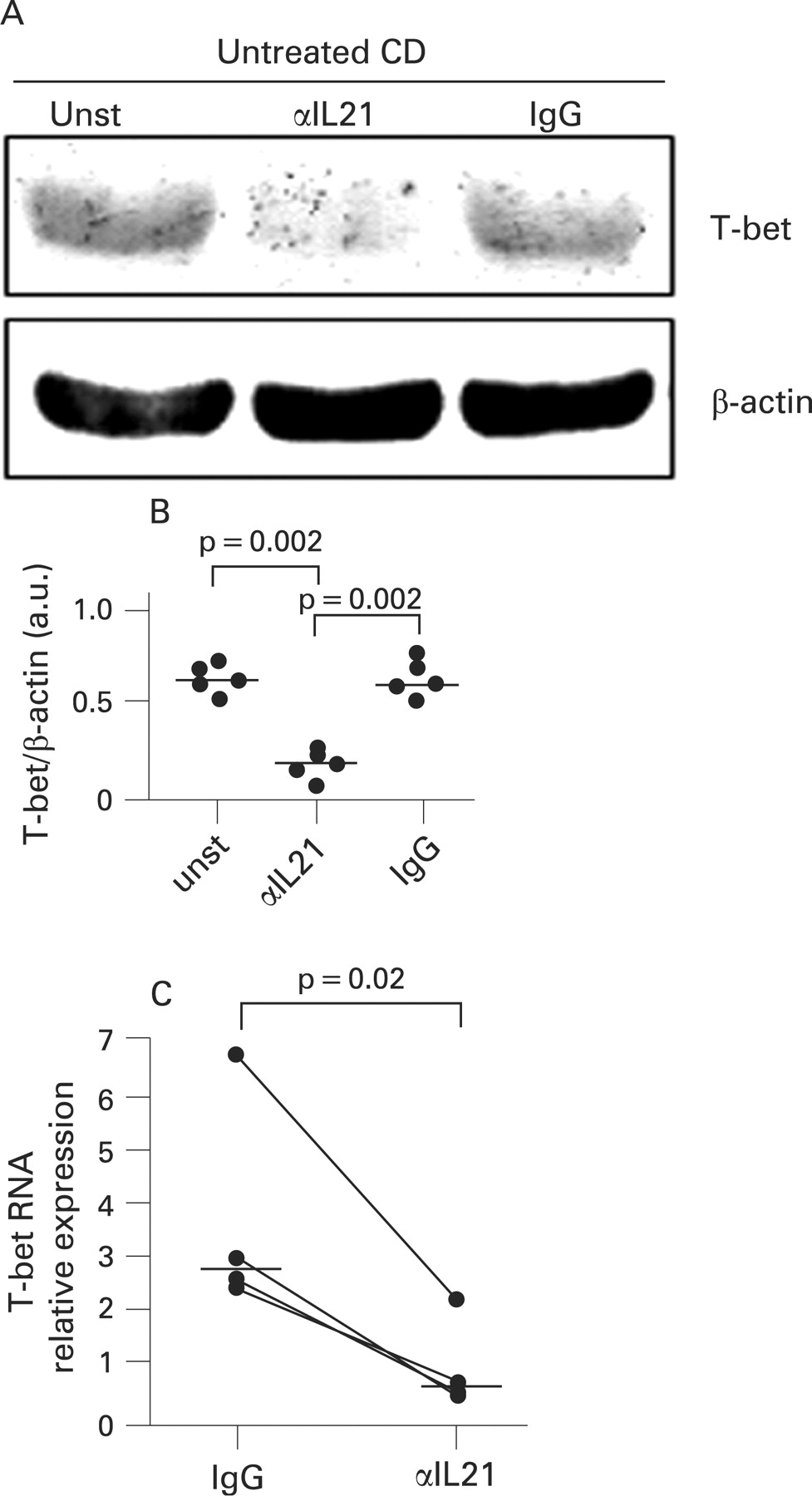
Since T-bet is a master regulator of the Th1 cell response,18 we also examined whether the anti-IL21-mediated downregulation of T-bet was associated with a decrease in IFNγ expression. The anti-IL21 but not control antibody significantly reduced both the secretion and the RNA content of IFNγ (fig 3A, B, p = 0.002).

The ability of IL21 to amplify Th1 cell responses can be further enhanced by IL15, a cytokine produced in excess in the gut of patients with active CD.19 20 Therefore, in subsequent experiments, we examined the effect of blocking IL21 and IL15 antibodies on the expression of Th1-associated markers in CD mucosa. The addition of either anti-IL21 or anti-IL15 antibody to ex vivo cultures of CD mucosal explants reduced T-bet and IFNγ RNA transcripts. In particular, when compared with IgG-treated explants, inhibition of both T-bet and IFNγ RNA was more pronounced in cultures treated with anti-IL15 (72% and 85%, respectively) than those treated with anti-IL21 (60% and 75%, respectively). Moreover, the simultaneous treatment of explants with anti-IL21 and anti-IL15 did not cause any further inhibition in T-bet and IFNγ in comparison with that seen in cultures treated with anti-IL15 alone (not shown).
In CD, gliadin-induced IL21 sustains T-bet and IFNγ expression
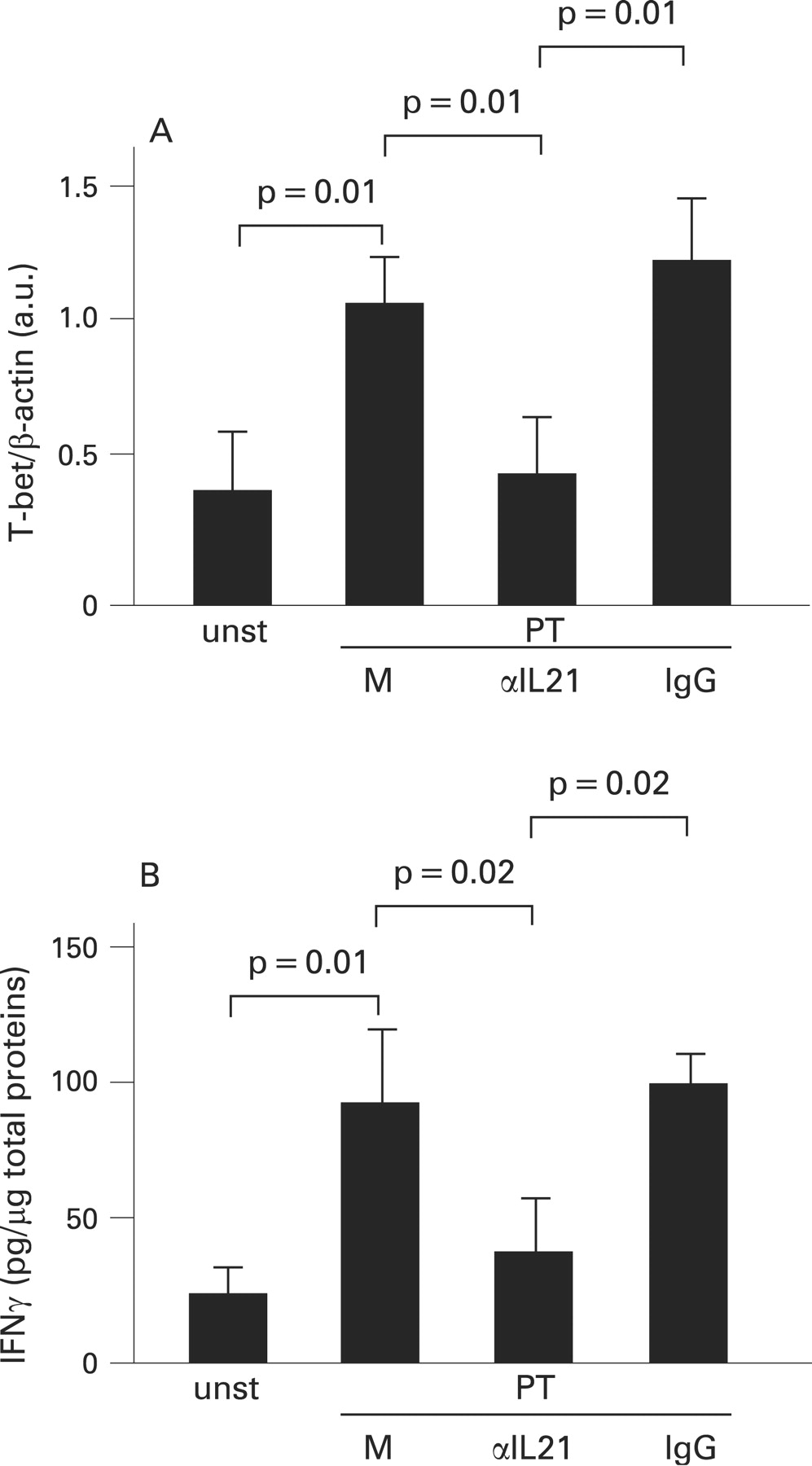
We next stimulated biopsies of treated CD patients and normal controls with PT for 24 h, and then examined the level of IL21 protein by western blotting. In explants from treated CD biopsies, but not controls, stimulation with PT enhanced IL21 (fig 4). We then examined whether blockade of IL21 inhibited the PT-mediated induction of T-bet. For this purpose, biopsies from treated CD patients were stimulated with PT in the presence or absence of anti-IL21 or control antibody. PT enhanced T-bet expression, and this effect was significantly reduced by anti-IL21 but not by control antibody (fig 5A, p = 0.01). Consistently, anti-IL21 significantly inhibited PT-driven IFNγ secretion by CD mucosal explants (fig 5B, p = 0.02).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
In active CD mucosa IL21 positively correlates with T-bet expression
Overall, the above results suggest that IL21 positively controls T-bet, thereby enhancing the Th1-mediated cell response in the duodenal mucosa of patients with untreated CD. To confirm this hypothesis further, we examined whether the expression of IL21 RNA correlates with the content of T-bet transcripts in CD. IL21 and T-bet transcripts were evaluated in biopsies taken from 12 untreated CD patients by real-time PCR. There was a clear direct relationships in individual samples between the expression of IL21 RNA and the amount of T-bet transcripts (r2 = 0.822, p<0.001).
DISCUSSION
In this study we show for the first time that in the mucosa of patients with untreated CD there is enhanced expression of IL21, at both the RNA and protein level. Densitometry analysis of western blots and real-time PCR assays revealed that levels of IL21 in CD samples were >4 times higher than that in controls. In contrast, IL21 was expressed at the same level in treated CD patients and normal controls, thus indicating that upregulation of IL21 in CD is dependent on the gluten-driven active inflammation. This was confirmed by the demonstration that stimulation of treated CD biopsies with gliadin enhances IL21 expression.
Since its discovery, IL21 production has been attributed to activated CD4+ T cells, and there is evidence that conditions inducing high levels of IL21 are those that best mimic CD4+ T cell activation.8 Consistent with these observations, we here show that, in CD mucosa, IL21 is expressed at a high level by CD4+ T-LPLs, while IL21 RNA was barely detectable in CD4– T-LPLs. At the same time, the demonstration that IL21 transcripts are more pronounced in CD4+ T-LPLs isolated from CD patients than controls suggests that the upregulation of IL21 in CD biopsies does not simply reflect the increased infiltration of the mucosa with CD4+ T-LPLs.
The mechanisms that lead to the overexpression of IL21 in CD remain to be elucidated. Studies with human blood T cells have shown that IL21 is preferentially made by Th1 cells, thus suggesting that factors that drive Th1 cell responses can positively regulate IL21 synthesis.13 15 In line with this, we previously showed that IL21 is upregulated in other Th1-mediated gastrointestinal inflammatory diseases, such as Crohn’s disease16 and Helicobacter pylori-related gastritis,21 and that, in Crohn’s disease, IL21 production is enhanced by IL12.16 In this context, it is however noteworthy that IL12 is not produced in CD,22 suggesting the existence of alternative pathways of induction of IL21. This is in line with the demonstration that IL21 production is distinct between NK T cells from different organs. In particular, IL21 production by liver-derived NK T cells seems to be dependent on the presence of IL12, whereas thymus- and spleen-derived NK T cells produce IL21 even in the absence of IL12.9
A major feature of active CD is the marked infiltration of the small intestinal mucosa with gluten-reactive T cells which overexpress T-bet and produce high levels of IFNγ.1 2 How Th1 cells are generated in CD remains unknown. A possibility is that naive T cells recruited from the blood are differentiated in situ along the Th1 pathway in response to factors that preferentially induce T-bet. It is also possible that Th1 cells primed in the intestinal secondary lymphoid organs seed the mucosa of CD patients, where locally produced molecules enhance T-bet expression and expand the gluten-specific Th1 response. Data of the present study suggest that IL21 may accomplish this function, because blockade of IL21 activity in mucosal explants of untreated CD patients reduces T-bet and leads to a diminished production of IFNγ. Moreover, stimulation of biopsies from treated CD patients with anti-IL21 prevents the gliadin-driven induction of T-bet and IFNγ. Although there is evidence that IL21 may control in vitro the survival of immune cells under specific circumstances,13 it is unlikely that the effect of anti-IL21 on the T-bet expression may be due to a cytotoxic effect on mucosal cells, because this antibody does not promote T cell apoptosis in vitro (personal unpublished observations). We previously showed that IFNγ positively regulates T-bet expression in CD mucosa through a process that involves JAK2/Stat molecules.6 Since IL21 is able to enhance IFNγ production, it is conceivable that some of the effects of IL21 on T-bet induction are mediated by IFNγ. We would, however, like to point out that both T-bet and IFNγ were examined in organ cultures of whole biopsies and not in purified cell types. As both these proteins can be expressed by several cell types (e.g. CD4+ and CD8+ T cells, NK cells, NK T cells), we think that it would be biologically relevant to examine T-bet and IFNγ in single mucosal cell types to ascertain if induction of T-bet by IL21 directly leads to IFNγ production. Unfortunately, we were not able to purify sufficient T, NK and NK T cells from pinch biopsies to carry out mechanistic studies. Together, our data suggest, however, that, in CD mucosa, optimal IL21 activity is necessary to enhance T-bet, thus amplifying and stabilizing the committed Th1 cell phenotype. These results confirm and expand on previous published studies showing that IL21 is not able to trigger Th1 cell differentiation but it can enhance ongoing Th1 cell responses. For example, Strengell et al showed that IL21 enhances IFNγ gene expression in activated human T cells and promotes the transcription of Th1-associated genes, including T-bet.15 The same group also documented a synergistic effect of IL21 and IL15 or IL21 and IL18 on IFNγ mRNA synthesis and production,19 whereas Kasaian et al demonstrated that IL21 cooperates with IL2 and IL15 in promoting IFNγ synthesis.23 Finally, we previously showed that IL21 enhances T-bet and p-Stat4 expression and sustains IFNγ synthesis in gut mucosal lymphocytes of patients with Crohn’s disease.16
In addition to its role in the regulation of Th1 cell differentiation, IL21 can maintain chronic inflammation and/or favour tissue damage by promoting the recruitment of immune cells within the inflamed tissue,17 the expansion of autoreactive T cells24 and the synthesis of extracellular matrix metalloproteinases.25
Acknowledgments
This work received support from the “Fondazione Umberto di Mario”, Rome, the Broad Medical Research Program Foundation (Grant No. IBD-0154R) and Giuliani SpA, Milan, Italy.
REFERENCES
Footnotes
Competing interests: None.
Ethics approval: This study received approval from the University Tor Vegata of Rome ethical committee.