Article Text

Abstract
Background Recent evidence has highlighted a potential role of interleukin 1β (IL-1β) in systemic sclerosis (SSc). NLRP1 provides a scaffold for the assembly of the inflammasome that promotes the processing and maturation of pro-IL-1β. In addition, NLRP1 variants were found to confer susceptibility to autoimmune disorders.
Objective To study a possible association of the NLRP1 rs6502867, rs2670660 and rs8182352, rs12150220 and rs4790797 with SSc in the European Caucasian population.
Methods NLRP1 single nucleotide polymorphisms were genotyped in 3227 individuals comprising a discovery set (870 SSc patients and 962 controls) and a replication set including individuals from Germany (532 SSc patients and 324 controls) and Italy (527 SSc patients and 301 controls), all individuals being of European Caucasian origin.
Results Conditional analyses revealed a significant association for the NLRP1 rs8182352 variant with both anti-topoisomerase-positive and SSc-related fibrosing alveolitis (FA) subsets under an additive model: p=0.0042, OR 1.23 (95% CI 1.07 to 1.41) and p=0.0065 OR 1.19 (95% CI 1.05 to 1.36), respectively. Logistic regression analysis showed an additive effect of IRF5 rs2004640, STAT4 rs7574865 and NLRP1 rs8182352 risk alleles on SSc-related FA.
Conclusions Our results establish NLRP1 as a new genetic susceptibility factor for SSc-related pulmonary fibrosis and anti-topoisomerase-positive SSc phenotypes. This provides new insights into the pathogenesis of SSc, underlining the potential role of innate immunity in particular in the FA-positive SSc subphenotype, which represents a severe subset of the disease.
Statistics from Altmetric.com
Several lines of evidence have recently raised the paradigm of the contribution of innate immune mechanisms to autoimmunity. The concept of innate immunity as a primitive system, being non-specific in most aspects of both recognition and response, has been transformed by the identification of the membrane-associated Toll-like receptors and the cytosol-expressed Nod-like receptors (NLR).1 ,2 The NLR family, pyrin domain containing 1 (NLRP1), is a member of the NLR family, which are cytoplasmic proteins that sense endogenous microbial products and metabolic stresses, thereby stimulating innate immunity. NLR associated with other proteins form multiprotein cytoplasmic complexes, the so-called inflammasomes. NLRP1 provides a scaffold for the assembly of the inflammasome that activates caspases 1 and 5, which subsequently promote the processing and maturation of the inflammatory cytokines, pro-interleukin 1β (IL-1β), IL-18 and IL-33.3 ,4
Some studies have highlighted a potential role of IL-1β in fibrotic disorders.5 ,6 In systemic sclerosis (SSc), the prototypic fibrotic connective tissue disorder, IL-1β has been reported to induce the fibrogenic phenotype of SSc fibroblasts and to increase strikingly in vitro the production of extracellular matrix by these cells.7 ,8 SSc is a chronic autoimmune disease with a complex pathogenesis driven by a combination of genetic risk factors and environmental events that lead to a break in immunological self-tolerance and systemic fibrosis.9 ,10 Two of the SSc susceptibility genes recently identified play a pivotal role in innate immunity: IRF5 and STAT4.11,–,14 In addition, other genes involved in innate immunity have been found to be associated with various autoimmune or auto-inflammatory disorders. NOD2 have thus been associated with a risk of Crohn's disease.15 Mutations of NLRP3 (also known as cryopyrin) were reported to cause three Mendelian dominant inflammatory disorders.16 Finally, NLRP1 has been found to confer a risk for extended autoimmune/inflammatory disorders17,–,19 and more recently to autoimmune Addison's disease and type I diabetes.20 ,21
Taking into consideration: (1) the potential role of IL-1β in the SSc fibrosing process; (2) the identification of SSc susceptibility genes encoding proteins involved in innate immunity; and (3) common genetic variants shared by different autoimmune diseases, we hypothesised that the previously identified NLRP1 risk variants may be involved in the susceptibility of SSc.
Patients and methods
Study population and study design
We performed a case–control association study including a replication step. The discovery sample included 870 SSc patients (83.6% women with a mean age (±SD) of 57.2±12.8) and 962 controls coming from the French network (82.8% women with a mean age (±SD) of 53.8±9.7).11 ,12 ,22 ,23 The replication step included individuals from Germany: 532 SSc patients (84.2% women with a mean age (±SD) of 56.1±8.3) and 324 controls (81.8% women with a mean age (±SD) of 56.3±7.1), and Italy: 527 SSc patients (86.1% women with a mean age (±SD) of 53.6±10.9) and 301 controls (80.2% women with a mean age (±SD) of 53.8±9.4). All SSc patients were classified according to LeRoy's cutaneous subtypes24 and phenotypically assessed as recommended.25 ,26 The characteristics of SSc patients are summarised in table 1.
Characteristics of the SSc patients in the three European populations investigated
The control groups consisted of healthy unrelated individuals who were ethnically matched to the SSc cases (all individuals were of European Caucasian ancestry). All local institutional review boards approved the study, and written informed consent was obtained from all subjects in the study. SSc patients were tested for antinuclear antibodies using indirect immunofluorescence and HEp-2 cells as antigen substrate (Antibodies, Davis, California, USA). Specific SSc antibodies were systematically assessed; anticentromere antibodies were determined by their distinctive indirect immunofluorescence pattern on HEp-2 cells. Anti-topoisomerase I (Topo) antibodies were determined by counter immunoelectrophoresis. Regarding lung fibrosis, fibrosing alveolitis (FA) was defined as the presence of typical features on chest high-resolution CT, which was collected for all SSc patients.
Genotyping
We selected the following five single nucleotide polymorphisms (SNP) for which convincing association was reported with vitiligo and/or autoimmune diseases: rs6502867, rs2670660 and rs8182352, rs12150220 and rs4790797.17 ,18 ,20 ,21 The five NLRP1 SNP were genotyped using a competitive allele specific PCR system (Kaspar genotyping; KBioscience, Hoddesdon, UK) as previously described.11 ,12 ,22 ,23 The average genotype completeness for both NLRP1 rs2670660, rs6502867, rs8182352, rs12150220 and rs4790797 polymorphisms was 98% for the SSc and the control samples. The accuracy was greater than 99%, according to duplicate genotyping of 10% of all samples using the Taqman SNP genotyping assay-allelic discrimination method (Applied Biosystems, Foster City, California, USA).
Statistical analysis
Statistical analyses were performed using the R computer package software (version 2.6.0). The level of significance for all the tests corresponds to a type I error rate α=5%. Tests for conformity with Hardy–Weinberg equilibrium were performed using a standard χ2 test (1 degree of freedom). Many methods for addressing the problem of multiple comparisons invoke a conservative approach, namely a standard Bonferroni correction, which assumes the independence of all tests performed. In many association studies, markers are not independent because they are in linkage disequilibrium (LD), and so a standard Bonferroni correction would be overly conservative.27 Therefore, we applied a correction for multiple testing for all the tests performed for a ‘generating hypothesis step’ when comparing SSc subgroups and controls (10 phenotypic subsets) in the ‘discovery’ sample. p Values remaining significant after this adjustment for multiple testing are indicated in the tables and termed ‘padj’ in the text. In the case of a defined hypothesis for any particular SSc phenotype detected in the discovery sample, we thereafter tested it in the relevant SSc samples (German and Italian sets) without correction for multiple testing. All the OR are provided with their 95% CI. Individual association analyses of NLRP1 SNP with SSc were performed by comparing cases and controls with a Fisher's exact test on genotype distribution. The corresponding OR were assessed using a standard logistic regression analysis with the homozygous genotype for the most frequent allele, in the control population, taken as the reference. The same procedure was applied in subgroups stratified according to SSc phenotypes.
Finally, we investigated whether the NLRP1 rs8182352 variant was statistically associated with a specific SSc subset. The main association concerned both SSc pulmonary fibrosis and Topo-positive subsets; so we performed multivariate logistic models including NLRP1 rs8182352, the Topo-positive status and the SSc diffuse cutaneous subset in order to assess the respective independent effect of these three genetic and phenotypic factors on pulmonary fibrosis.
Meta-analysis on NLRP1 rs8182352
The analysis of the combined data including the three populations was performed: (1) by the calculation of homogeneity of OR among cohorts using Breslow–Day and Woolf Q methods; (2) by calculation of the pooled OR under a fixed-effects model (Mantel–Haenszel meta-analysis) or random effects (DerSimonian–Laird) when necessary assessed by logistic regression analysis genetic effects under three modes of inheritance: additive, dominant and recessive. The meta-analysis was performed using logistic regression analysis adjusted for the three populations investigated.
Power calculation
Power calculations are driven through an asymptotic non-central χ2 approach as previously described.28 The discovery set of 870 SSc cases and 962 controls provides a power greater than 80% to detect the association between SSc and the NLRP1 rs8182352 risk allele with an OR of 1.30 at the 5% significance level.
Results
Discovery set
Genotype frequencies were in Hardy–Weinberg equilibrium in the control population for all the NLRP1 SNP investigated.
Single marker analysis
The case–control association study performed in the discovery sample (n=870 SSc patients and 962 controls) did not reveal any association between the five NLRP1 SNP and the SSc population as a whole. When SSc subphenotypes were queried we observed a significant increase in the frequency of the rare NLRP1 rs8182352 C allele in the Topo-positive SSc subset compared with the controls: padj=0.047, OR 1.35, 95% CI 1.09 to 1.66). Analysis of the genotype distributions also revealed a strong increase in frequency of the homozygous CC genotype in SSc patients compared with controls: padj=0.038, OR 1.83, 95% CI 1.21 to 2.76 (table 2). We also observed a strong increase in frequency of both the rs8182352 C risk allele and the homozygous CC genotype in the subset of SSc patients having FA, however, it did not reach statistical significance (table 2).
Genotype and allele distribution of the NLRP1 rs8182352 SNP in SSc patients and controls in the French Caucasian population
Regarding the NLRP1 rs2670660 SNP, we observed an increased frequency of the rare allele in both Topo-positive and FA-positive SSc subsets, however, not reaching statistical significance after correction for multiple testing. In agreement with the previously reported LD analysis, NLRP1 rs2670660 and rs8182352 were found to be in relatively high LD (r2=0.71). Conditional analyses between rs2670660 and rs8182352 revealed a significant association restricted to the NLRP1 rs8182352 risk allele (p=0.014) (table 2 and supplementary file 1, available online only).
No association between SSc and the NLRP1 rs6502867, rs12150220 and rs4790797 SNP was detected with any of the subphenotypes investigated (supplementary file 1, available online only). In agreement with the previously reported LD analysis, NLRP1 rs2670660 and rs8182352 were found to be in LD (r2=0.709), whereas rs6502867 was not in LD with both rs2670660 and rs8182352 (r2=0.057 and 0.056, respectively) (figure 1). The pairwise correlation coefficient found rs12150220 in LD with rs2670660 (r2=0.59) and rs8182352 (r2=0.56), whereas rs6502867 and rs4790797 were found to belong to distinct LD blocks (supplementary figure 1, available online only).
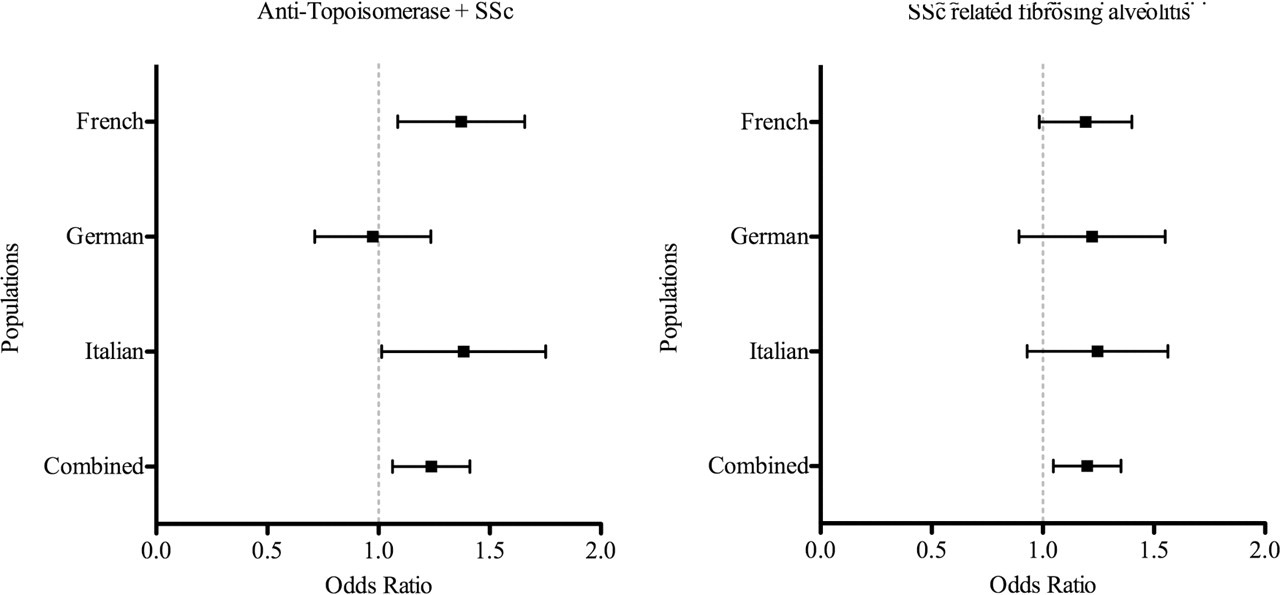
Association between NLRP1 rs8182352 and systemic sclerosis (SSc) in the European Caucasian population: meta-analysis.
Replication set
Following the results obtained in the discovery set, we selected the NLRP1 rs8182352 SNP to be investigated in Topo-positive SSc subset. Genotype frequencies of rs8182352 were in Hardy–Weinberg equilibrium in the control population. No association was detected in the German population. Conversely, the association between NLRP1 rs8182352 and the Topo-positive SSc subset was replicated in the Italian population (table 3). Taking into account the striking differences in frequency of the NLRP1 rs8182352 risk allele when comparing SSc FA-positive with the controls and an elevated OR (1.41 for the homozygous risk genotype) for a given complex disease, although not reaching significance, we decided to follow this association in the replication sample. We assume that the lack of significance in the first set relates to an insufficient statistical power, and considering the critical weight of FA on SSc morbidity and mortality, was another argument to follow the association. Interestingly, as observed in the French population we detected a strong increase in frequency of the risk allele in the FA-positive SSc subset in both German and Italian populations; however, not reaching statistical significance (table 3).
Association study of NLRP1 rs8182352 with SSc in both German and Italian Caucasian populations
Meta-analysis
The combinability test according to the Breslow–Day method showed no significant differences among them. Therefore, we performed a meta-analysis using the Mantel–Haenszel test under fixed effects.
The NLRP1 rs8182352 SNP was found to confer susceptibility to both Topo-positive SSc and SSc-related FA, under an additive model: pooled OR 1.23 (95% CI 1.07 to 1.41) and 1.19 (95% CI 1.05 to 1.35), respectively. These results are detailed in table 4 shown in figure 1 and supplementary data, available online only.
Meta-analysis of the NLRP1 rs8182352 variant in the European Caucasian population
Intra-cohort analyses found that the NLRP1 rs8182352 C allele and CC genotype frequencies were significantly increased in Topo-positive SSc compared with Topo-negative SSc: 50.4% versus 46.3%, p=0.025 and 24.8% versus 21.6%, p=0.027, respectively. In addition, both the C allele and CC genotype were increased in FA-positive compared with FA-negative SSc: 49.7% versus 46.2%, p=0.046 and 25.4% versus 20.9%, p=0.041, respectively. As FA is strongly linked to the presence of anti-topoisomerase,29 we next performed multivariate logistic regression analysis including NLRP1 rs8182352 and Topo and FA status in order to assess the respective independent effect of NLRP1 rs8182352 on pulmonary fibrosis and/or Topo-positive subsets. Regarding the association of NLRP1 rs8182352 with the FA adjusted for Topo status and with Topo adjusted for FA, no association was detected: OR 1.14 (95% CI 0.97 to 1.34), p=0.11 and OR 1.13 (95% CI 0.95 to 1.34), p=0.17, respectively. Multivariate logistic regression analysis adjusted for the dcSSc status found similar results (data not shown). Therefore, NLRP1 contribute to both Topo-positive and FA-positive SSc phenotypes, which are strongly linked.
Joint effects of the IRF5 rs20046640, STAT4 rs7574865 and NLRP1 rs8182352 risk alleles on SSc-related FA susceptibility
As we have recently reported that carrying at least three IRF5 rs2004640/STAT4 rs7574865 risk alleles is an independent marker of SSc-related FA,12 we decided to investigate the potential joint effects of the risk alleles of NLRP1, IRF5 and STAT4 in the French sample of SSc patients and controls using an additive logistic regression model. When gene–gene interaction between NLRP1 and IRF5 and STAT4 was queried, no evidence for dominance or interactions was observed between IRF5 rs2004640, STAT4 rs7574865 and NLRP1 rs8182352 risk alleles using multiple logistic regression analysis ruling out epistatic effects. We observed an additive effect of risk alleles on the FA-positive SSc susceptibility. The OR for FA-positive SSc were 1.82, 95% CI 1.25 to 2.65; p=0.001 for carriers of three risk alleles among these three SNP, 2.47, 95% CI 1.62 to 3.75; p<0.0001 for carriers of four risk alleles, and 2.91, 95% CI 1.58 to 5.36; p<0.0001 for carriers of five or six risk alleles.
Intracohort analysis comparing FA-positive and FA-negative subsets revealed that SSc patients carrying at least four IRF5, STAT4, NLRP1 risk alleles were at increased risk of FA. The OR for FA-positive SSc were 1.68, 95% CI 1.05 to 2.67; p=0.019 for carriers of four risk alleles, 2.03, 95% CI 1.03 to 4.04; p=0.033 for carriers of five or six risk alleles.
In these settings, figure 2 shows OR for FA-positive SSc individuals carrying three, four, five or six risk alleles, using the individuals with no or one risk allele as reference. The OR increases as a function of the number of risk alleles in a multiplicative manner, with a 1.33-fold increase in OR for each additional risk allele.

{kind=link}
{kind=link}
Additive effect of IRF5 rs2004640, STAT4 rs7574865 and NLRP1 rs8182352 risk alleles on systemic sclerosis (SSc)-related fibrosing alveolitis susceptibility.
Discussion
In this study our aim was to test for association of NLRP1 with SSc. We first detected in the discovery set an association between NLRP1 rs2670660 and rs8182352 with SSc Topo-positive and FA subsets and Topo-positive subgroup, respectively. Conditional analysis suggested that the association with this region could best be explained by the NLRP1 rs8182352 variant. Indeed, we next investigated the NLRP1 rs8182352 polymorphism in two European Caucasian populations. The association was replicated in the Italian population but not in the German. The weak sample size of the German sample may partly account for this unexpected finding. However, some data showed that FA may also occur as a limited cutaneous disease and this can also account for the discrepancy between the association with Topo or FA.30 Nonetheless, the meta-analysis that is corrected for population stratification further strengthens this association and supports its soundness, showing that the NLRP1 rs8182352 rare allele acts with a risk effect, under an additive model on both SSc-related pulmonary fibrosis and Topo-positive subsets. Multivariate analysis revealed that NLRP1 was associated with both SSc subsets, illustrating the high correlation of Topo positivity and SSc-related pulmonary fibrosis.
We then investigated the NLRP1 rs8182352 SNP, a polymorphism with no predicted functional role. Both allele and genotype frequencies were in agreement with those previously reported in Caucasian populations,17 ,18 supporting the accuracy of this genotyping. The restricted association of NLRP1 rs8182352 with both FA-positive and Topo-positive SSc subsets suggests that this variant could contribute to a disease-specific phenotype, and may be regarded as a marker of SSc severity, lung disease being actually the primary cause of SSc-related death.31 No other NLRP1 variant investigated was found to be associated with SSc; notably, the non-synonymous NLRP1 rs12150220 polymorphism, reported to confer susceptibility to autoimmune diseases.20 ,21 Interestingly, this risk variant is located in the same NLRP1 LD block tagged by rs8182352.20 Therefore, taking into account that the putative functional role of the NLRP1 rs8182352 SNP is unknown, re-sequencing of the LD block tagged by rs8182352 may be essential in order to identify one or multiple SSc causal variants. In addition, future functional analyses of the candidate NLRP1 causal variants are mandatory to obtain a better understanding of the contribution of NLRP1 in the pathogenesis of SSc.
NLRP1 is believed to mediate inflammation and autoimmunity partly due to its ability to form the inflammasome that plays a pivotal role in the activation of pro-IL-1β.32 Several lines of evidence have raised the contribution of IL-1β in SSc pathogenesis, including an increased expression of IL-1β by fibroblasts detected in skin biopsies, high IL-1β serum concentrations and overexpression of IL-1β and IL-1ra genes in bronchoalveolar lavage fluid of SSc patients compared with controls.33,–,36 Nevertheless, it remains to be determined whether IL-1β may induce the fibrotic process in the lung or contribute to the chronicity and severity of involvement. In addition, it has previously been reported that silica exposure, which is a well-known SSc environmental factor, leads to the local release of IL-1β, and may contribute to abnormal connective tissue deposition by stimulating fibroblast proliferation.37,–,39
Of most interest, we found an additive effect of the rare variants of these genes on SSc-related FA, with a 1.33-fold increase in OR for each additional risk allele. Furthermore, intra-cohort analysis revealed that SSc patients carrying at least five IRF5, STAT4, NLRP1 risk alleles were at a twofold increased risk of having FA. The identification of IRF5, STAT4 and NALP1 as new SSc-related pulmonary fibrosis genetic susceptibility factors raises the question of the contribution of innate immunity in SSc pathogenesis and in the fibrotic process, notably through IL-1β and the type I interferon pathway. However, this model, observed in the French Caucasian population, needs to be replicated in other populations to be validated.
In the present study we show for the first time that the NLRP1 rs8182352 variant plays a significant role in SSc-related FA and SSc Topo-positive susceptibility. These findings were obtained through large samples and replication in an independent cohort. The fact that genotyping provided similar frequencies in both control groups underlines the consistency of the reported results. Intracohort analyses revealed an additive effect of NLRP1 with IRF5 and STAT4 risk variants on SSc-related pulmonary fibrosis susceptibility. These data provide new insights into the pathogenesis of SSc, underlying the pivotal key role of innate immunity at least in the FA SSc subphenotype.
Acknowledgments
The authors would like to thank Dr Joëlle Benessiano (Centre de Ressource Biologique, Hôpital Bichat Claude-Bernard, Paris France) and staff members of the Établissement Français du Sang for their assistance and Mrs Carole Desbas for expert secretarial assistance. The authors also thank Professor Bernard Grandchamp and Dr Nadem Soufir (Hôpital Bichat Claude-Bernard) for assistance in setting up the French Caucasian control sample.
References
Supplementary materials
Web Only Data
Files in this Data Supplement:
{kind=link}
Footnotes
-
Funding ANR (grant R070994KS), SFR, ASF, GFRS, INSERM. The German Network for Systemic Sclerosis was funded by the German Federal Ministry for Education and Research (grant nos 01 GM 0310 and 01 GM 0634 to IM, GR, NH and IT).
-
Competing interests None.
-
Patient consent Obtained.
-
Ethics approval This study was conducted with the approval of the Comité de Protection des Personnes of Cochin Hospital, Paris, France.
-
Provenance and peer review Not commissioned; externally peer reviewed.