Article Text

Abstract
Objectives In the inflamed synovium of patients with rheumatoid arthritis (RA), autoantibodies to citrullinated proteins (ACPA) probably form immune complexes (IC) on deposits of citrullinated fibrin. We showed that in vitro such ACPA-IC activate a pro-inflammatory cytokine response in M-CSF-differentiated macrophages. Our objective was to evaluate how macrophage polarisation influences this response.
Methods CD14-positive monocytes from healthy donors were cultured in the presence of M-CSF, IFN-γ, interleukin (IL)-4 or IL-10. Expression of markers specific for polarised macrophages was analysed by flow cytometry. Their cytokine secretion was prompted by in vitro generated autoantibodies to citrullinated proteins immune complexes (ACPA-IC) and assayed in the culture supernatants.
Results IFN-γ-polarised cells exhibited high levels of CD64 and CD80. Low expression of CD14 and high expression of CD206 characterised the IL-4-polarised cells. Exposure to IL-10 or M-CSF raised the expression of CD14, CD32 and CD163. The two cell types lacked CD80 and exhibited similar expression of CD64, CD200R and CD206. In response to ACPA-IC, the secretion of IL-1β, IL-6 and IL-8 was similar among cells exposed to IFN-γ, IL-4 or IL-10. However, the later cells were associated with the highest IL-1Ra:IL-1β ratio and the lowest tumour necrosis factor (TNF)-α:IL-10 ratio. Conversely, M-CSF-exposed cells secreted the highest levels of pro-inflammatory cytokines, exhibited a high TNF-α:IL-10 ratio and the lowest IL-1Ra:IL-1β ratio.
Conclusions Despite their phenotypic similarity, IL-10-polarised and M-CSF-polarised macrophages clearly differ in their cytokine response to ACPA-IC. M-CSF-polarised cells exhibit the highest pro-inflammatory potential. Since M-CSF is abundant in the RA synovium, therein it probably drives macrophages towards a strong pro-inflammatory cytokine response to the locally formed ACPA-IC.
- Rheumatoid Arthritis
- Ant-CCP
- Cytokines
- Synovitis
- TNF-alpha
Statistics from Altmetric.com
Introduction
Macrophages play crucial roles in tissue homeostasis and immunity.1 ,2 Their extreme phenotypic and functional heterogeneities, that reflect their specialisation in various microenvironments and their plasticity, are conveyed by the now widely used term ‘polarisation’.3–8 Macrophages with different activation states or phenotypes were first classified as ‘M1’ or ‘M2’, the former being involved in inflammation and host defence and the latter having anti-inflammatory or tissue repair functions. This classification was refined, dividing macrophages into three homeostatic functions: host defence (corresponding to M1 ‘inflammatory’ macrophages), tissue repair (M2 ‘wound-healing’ macrophages) and immunoregulation (M2 ‘immunoregulatory’ macrophages). The current consensus is that these polarisation states can be generated in vitro, using interferon (IFN)-γ, interleukin (IL)-4 or IL-10, respectively.9
Besides their vital contribution in health maintenance, macrophages play major deleterious roles in numerous inflammatory diseases.10–12 In rheumatoid arthritis (RA), an increase in the number of macrophages in the synovial tissue (ST) constitutes an early event that correlates with the disease activity13 and the degree of joint erosion.14 This increase probably results both from expansion of resident macrophages and from infiltration by circulating monocytes.10 Sublining macrophage numbers predict responsiveness to therapy, independently of the treatment type.15 ,16 Both in the acute and chronic phases of RA, synovial macrophages become activated and contribute to the disease by secretion of pro-inflammatory cytokines and mediators that drive angiogenesis, infiltration by immune cells and cartilage and bone destruction.17 They constitute a major source of tumour necrosis factor (TNF)-α which is abundantly produced and constitutes a master cytokine in all manifestations of the disease.18
In the ST, immune complexes (IC) containing the tightly RA-associated IgG autoantibodies directed to citrullinated proteins (ACPA) probably promote the secretion of pro-inflammatory cytokines by macrophages. We developed a human in vitro model that allowed demonstrating the Fcγ receptor (FcγR)-dependent inflammatory potential of ACPA.19–21 Macrophages differentiated in vitro from monocytes of healthy donors in the presence of macrophage colony-stimulating factor (M-CSF) secreted TNF-α upon stimulation by autoantibodies to citrullinated proteins immune complexes (ACPA-IC) generated by immunocapture of ACPA from RA sera by in vitro citrullinated human fibrinogen (C-FBG) immobilised on culture wells.19 This well-established ACPA target was chosen because of the conclusively demonstrated frequent presence of citrullinated fibrin in RA synovial membranes.22 ,23 Furthermore, fibrin formation in the ST is an inflammation-dependent phenomenon. As the disease-specific interaction between ACPA and citrullinated fibrin activates pro-inflammatory effector mechanisms, new fibrin deposits are formed which can become new ACPA targets as they are citrullinated by locally expressed peptidylarginine deiminases, thereby fuelling the synovitis-promoting formation of ACPA-IC.24 ,25 Last but not least, we showed that the autoantibodies recognising C-FBG encompass almost all ACPA, that is, those detected in the widely used second-generation anti-cyclic citrullinated peptide antibodies assay (CCP2 assay), meaning that autoantibodies able to interact with this autoantigen are present in a vast majority of the ACPA-positive patients.26–28 Since macrophages can be functionally heterogeneous, the aim of the present study was to evaluate the influence of polarisation on their response to ACPA-IC. Archetypal inflammatory, wound healing or immunoregulatory macrophages were obtained from monocytes of healthy donors by culture in the presence of IFN-γ, IL-4 or IL-10, respectively. Their expression of the subset-specific phenotypic markers recently proposed by Ambarus et al29 was assessed, and their pro-inflammatory and anti-inflammatory cytokine responses to ACPA-IC were compared with each other, as well as with those of the M-CSF-polarised macrophages.
Methods
Monocyte isolation and in vitro polarisation
CD14-expressing monocytes were purified by positive selection from buffy coats of healthy donors as detailed formerly.19 They were then polarised essentially as described by Ambarus et al.29 Briefly, they were incubated for 4 days at 106 cells/mL in perfluoroalkoxy polymer inserts in Iscove's Modified Dulbecco's Medium (IMDM, Gibco) containing 10% fetal calf serum (FCS) (referred to as cIMDM) and supplemented with M-CSF (100 ng/mL), IFN-γ (50 ng/mL), IL-4 (40 ng/mL) or IL-10 (50 ng/mL). Cells cultured in complete Iscove’s Modified Dulbecco’s Medium (cIMDM) alone served as control cells. Macrophages cultured in macrophage–serum free medium (SFM) medium (Gibco) containing 10% FCS (referred to as complete serum free medium (cSFM)) supplemented with M-CSF (100 ng/mL), corresponding to the culture condition used in our initial model,19 were generated for comparison.
Flow cytometry
The expression of surface markers CD14, CD16, CD32, CD64, CD80, CD163, CD200R and CD206 by monocytes and the diversely polarised macrophages was evaluated by flow cytometry (see online supplementary information for details on the used antibodies). Samples were acquired on a FACScan flow cytometer (Becton Dickinson) and data were analysed using FlowJo software (Milteniy Biotec). Expression values correspond to the ratio of the geometric mean fluorescence intensity of the marker, measured on live cells gated using forward scatter (FSC) and side scatter (SSC) characteristics, to that of the isotype control.
Supplemental material
Cell stimulation
Stimulation by ACPA-IC was performed as previously described.19 Briefly, 96-well plates were coated with C-FBG and IC were generated by incubation with the IgG fraction of an ACPA-positive RA serum pool (ACPA+IgG). The IgG fraction of an ACPA-negative RA serum pool (ACPA− IgG) was used as control. Both IgG fractions were used at the same concentration (2.5 mg/mL) and purified as previously described.19 ,20 After washing, each well received 50 000 cells in a serum-free and cytokine-free version of the medium used during polarisation (IMDM or SFM). Alternatively, cells were put into uncoated wells and stimulated by lipopolysaccharide (LPS) (0.5 µg/mL from E. coli strain 055/B5). TNF-α and IL-10 concentrations in 24 h culture supernatants were measured by ELISA (eBiosciences). IL-1β, IL-6, IL-8 and IL-1Ra were evaluated in a multiplexed assay (Procarta Cytokine Profiling Kit) read with a Luminex 100 IS flow cytometer (Luminex). The means of triplicate measurements were taken into account. Cytokine responses after stimulation correspond to secretions specifically prompted by ACPA-IC or by LPS. These were calculated by subtracting background cytokine secretion measured in control conditions, that is, when ACPA− IgG replaced ACPA+ IgG or after culture in medium alone, respectively. For the calculation of cytokine ratios, zero values (values inferior to the cytokine detection limit) were replaced by 1.
Statistical analyses
Differences between the various cell populations in expression of surface markers or secretion of cytokines were assessed by Friedman's test and the variables were pairwise compared by Wilcoxon's test. Correlations were assessed by computing Spearman's rank correlation coefficients. In all statistical analyses, the null hypothesis was rejected when the p value was smaller than a threshold computed by applying Holm's sequential Bonferroni adjustment to a 5% level of significance.
Results
Surface markers of differentially polarised macrophages
The expression of CD14, CD16, CD32, CD64, CD80, CD163, CD200R and CD206 was analysed by flow cytometry after 4-day culture of CD14-positive monocytes from healthy donors either in the absence of a polarising cytokine (control cells) or in the presence of IFN-γ, IL-4, IL-10 or M-CSF (figure 1A). In comparison with monocytes, a significant downregulation of CD14, CD32 and CD64 and a significant upregulation of CD206 occurred in control cells. In comparison with control cells or with cells cultured in the presence of another cytokine, IFN-γ-polarised cells were characterised by very low expression of CD32 (p=0.0004 for IFN-γ vs control, IL-10 or M-CSF; p=0.0113 vs IL-4) and high levels of CD64 (all p=0.0004) and of CD80 (all p≤0.0008). In addition, CD200R expression was particularly low when compared with that of cells exposed to the other cytokines (p=0.0004 for all comparisons). IL-4-polarised cells differed from all other cells by their characteristically low expression of CD14 (all p≤0.0007) and of CD64 (all p=0.0004) and their remarkably high expression of CD206 (all p≤0.0010). In comparison with culture in medium alone or in the presence of IFN-γ or IL-4, exposure to IL-10 resulted in significantly higher levels of CD14 (all p≤0.0007), of CD32 (p=0.0004 vs medium or IFN-γ, p=0.0084 vs IL-4) and of CD163 (p=0.0022 vs medium, p≤0.0008 vs IFN-γ or IL-4). A similar change occurred after culture in the presence of M-CSF, as the derived cells also differed from the control and IFN-γ-polarised or IL-4-polarised cells by their higher expression of CD14 (all p values ≤0.0007), of CD32 (p=0.0004 vs control or IFN-γ, p=0.0077 vs IL-4) and of CD163 (all p≤0.0005). Moreover, IL-10-polarised or M-CSF-polarised cells both exhibited enhanced expression of CD16 in comparison with control cells (p=0.0007 or 0.0004, respectively), even though for this marker, M-CSF-polarised cells exhibited higher expression (p=0.0006). Therefore, despite significant differences in the expression of CD14, CD16 and CD206, the M-CSF-polarised and the IL-10-polarised cells exhibited similar phenotypes, characterised by an absence of CD80, high expression of CD14, CD32, CD163 and CD200R and low expression of CD206. This phenotypic resemblance was confirmed by hierarchical clustering of the median ratios of expression of each marker in the cytokine-exposed cells to its expression in the control cells. Indeed, the heat map and dendrogram shown in figure 1B illustrate that three distinct surface marker expression profiles distinguish polarisation induced by the three macrophage polarising cytokines IFN-γ, IL-4 or IL-10, and that the profiles adopted by cells exposed to M-CSF and to IL-10 were very similar. Finally, the culture medium also had an impact on this profile since on M-CSF-influenced cells significantly different levels of CD64, CD163 and CD200R were observed depending on the culture medium (cIMDM or cSFM; p=0.0077, 0.0061 and 0.0004, respectively; figure 1A). However, the two cell types expressed the same sets of markers, showing that cells of this model exhibit an ‘IL-10-like’ surface marker profile.

Expression of phenotypic markers on monocytes and on differentially polarised monocyte-derived macrophages. (A) Expression of CD14, CD32, CD80, CD64, CD16, CD163, CD206 and CD200R on blood monocytes and monocyte-derived macrophages obtained by culture in cIMDM alone (control), in cIMDM supplemented with IFN-γ, interleukin (IL)-4, IL-10 or M-CSF or in cSFM supplemented with M-CSF. Plots represent geometric mean fluorescence intensity (gMFI) ratios of the marker to that of the isotype control, boxes span 25th–75th centiles, central lines denote median values and whiskers indicate the non-outlier range. Horizontal lines connect conditions with significantly different marker expressions. Significant differences with control macrophages or between control macrophages and monocytes are indicated by asterisks or squares, respectively. (B) Heat map representation and hierarchical clustering dendrogram for the median values of expression of each marker in the indicated cytokine-polarised macrophages relative to its expression in control macrophages. Data represent 16 independent experiments with the cells of different blood donors.
Cytokine responses of differentially polarised macrophages
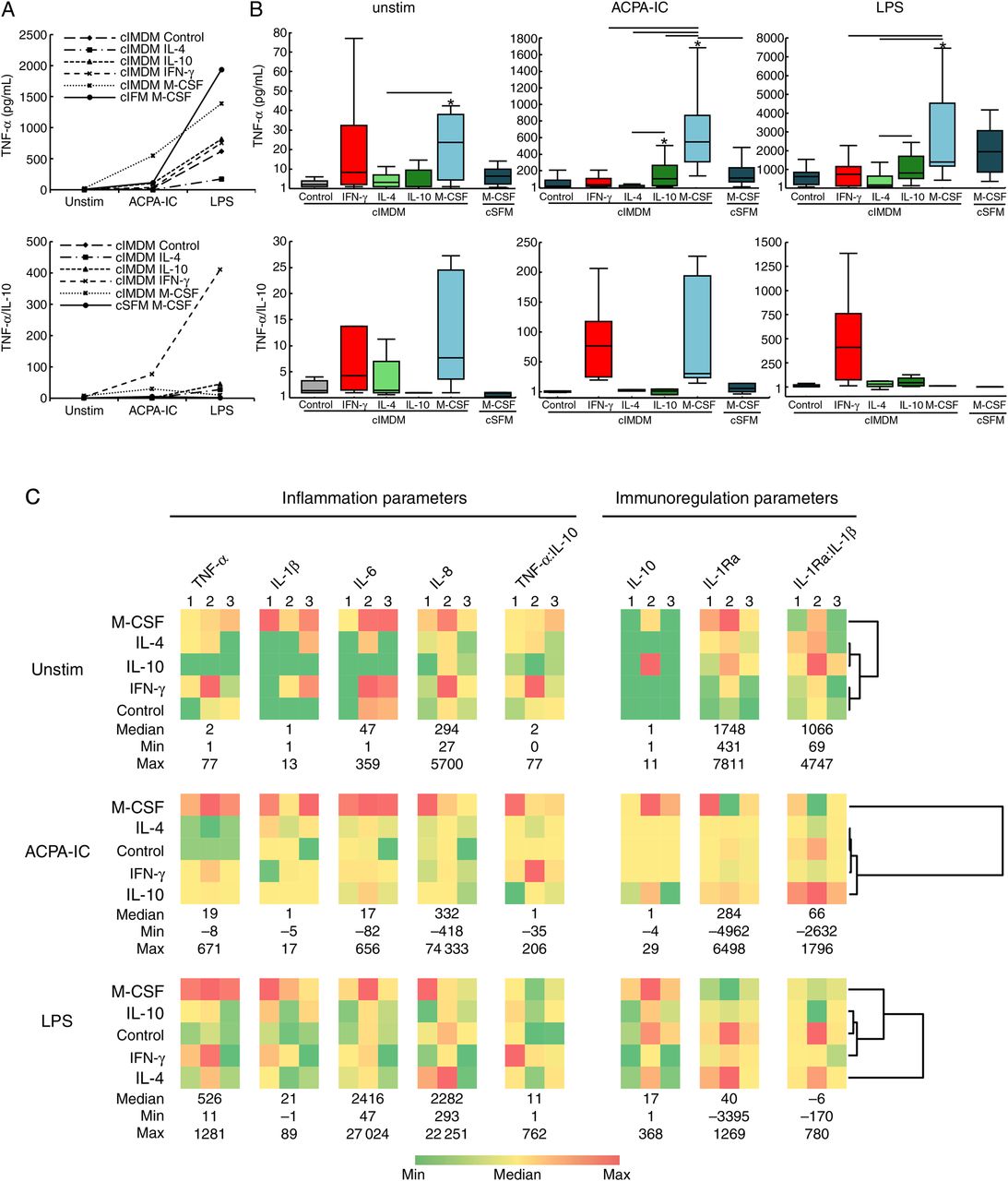
The inflammatory potential of the responses of the differentially polarised macrophages after stimulation by ACPA-IC or LPS was appreciated following assay of pro-inflammatory and anti-inflammatory cytokines in culture supernatants. In particular, as indicators of the pro-inflammatory potential of the cytokine secretion pattern, the TNF-α secretion of cells from 14 donors and the ratios of the TNF-α to the IL-10 secretion for cells from a subset of six of these donors were evaluated (figure 2A, B). Spontaneous secretion of TNF-α in the absence of any stimulation was low (median generally below 10 pg/mL) although it was somewhat higher in cells cultured in cIMDM and exposed to M-CSF (median 24 pg/mL, range 1–126). Similarly, IL-10 secretion was generally very low (ranging from undetectable to 16.5 pg/mL; data not shown) and although the TNF-α:IL-10 ratios tended to be somewhat higher for IFN-γ-polarised or M-CSF-polarised cells (median 4.3, range 1–76.8 and median 7.7, range 1–27.3, respectively), they were generally low (median below 1.5 for all other culture conditions). After stimulation by LPS, the TNF-α concentrations reached similarly high levels among control cells (median 621 pg/mL, range 73–1543), IFN-γ-polarised cells (median 754 pg/mL, range 11–2270) and IL-10-polarised cells (816 pg/mL, range 139–4722). The latter produced significantly more TNF-α than IL-4-polarised cells did (median 173 pg/mL, range 41–1385; p=0.0029). In addition, cells cultured in cIMDM and exposed to M-CSF produced significantly higher levels than IFN-γ-polarised cells (p=0.0019) or IL-4-polarised cells (p=0.0029). Moreover, even though, after correcting for multiple comparisons, the difference was not statistically significant, TNF-α secretion by IL-10-polarised cells also tended to be comparatively lower (p=0.0092). Finally, IFN-γ-polarised cells produced a TNF-α:IL-10 ratio (median 410, range 11–1387) that clearly appeared higher than that of all other cells, although, notably due to the multiple comparisons, the differences were not statistically significant. After stimulation by ACPA-IC, the TNF-α response did not differ among control cells, IFN-γ-polarised or IL-4-polarised cells (median 16 pg/mL, range −5–206; median 34 pg/mL, range −4–206; median 1.5 pg/mL, range 7–41, respectively), while for IL-10-polarised cells (median 104, range 3–1594) TNF-α secretion was higher comparatively with control cells and IL-4-polarised cells (p=0.0035 and p=0.0019, respectively). However, similarly to what was observed after LPS stimulation, the TNF-α:IL-10 ratio reached higher levels for IFN-γ-polarised cells (median 77, range 19–206) than for control cells (median 1, range 1–1; p=0.0277) or IL-4-polarised cells (median 1, range −8–25; p=0.0277) or IL-10-polarised cells (median 1, range −35–35; p=0.0277). Furthermore, very interestingly, cells exposed to M-CSF in cIMDM secreted the highest TNF-α concentrations (median 550, range 139–2257; p<0.0015 in comparison with all other cells), and their TNF-α:IL-10 ratio (median 30, range 14–227) was similar to that obtained for IFN-γ-polarised cells (p=0.9125). Of note, the culture medium influenced the cytokine response to ACPA-IC since TNF-α secretion for cells cultured in cSFM+MCSF (median 113.5 pg/mL, range 5.5–481) was lower than that of cells cultured in cIMDM+M-CSF (p=0.0015), and the TNF-α:IL-10 ratio (median 5, range −4–49) tended to be lower after culture in cSFM (p=0.0277). Finally, no correlations were found between TNF-α secretion or the TNF-α:IL-10 ratio and the expression of surface markers in any of the differentially polarised cells whether left unstimulated or stimulated by ACPA-IC or LPS.

{kind=link}
{kind=link}
Cytokine secretion by polarised macrophages in response to autoantibodies to citrullinated proteins immune complexes (ACPA-IC) and LPS. (A) Median spontaneous (unstim) or ACPA-IC-induced or LPS-induced secretion of tumour necrosis factor (TNF)-α or ratio of the TNF-α to the interleukin (IL)-10 (TNF-α:IL-10) secretions in macrophages obtained by culture in cIMDM alone (control), in cIMDM supplemented with IFN-γ, IL-4, IL-10 or M-CSF or in cSFM supplemented with M-CSF. TNF-α data were obtained in 14 independent experiments and TNF-α:IL-10 data in a subset of six experiments. (B) Box-and-whisker plot representations (as in figure 1) of the data are shown in (A), separately for the three conditions of stimulation. Significant differences are pointed as in figure 1. (C) Heat map representations and hierarchical clustering dendrograms for the spontaneous (unstim), ACPA-IC-induced or LPS-induced secretions of the pro-inflammatory cytokines TNF-α, IL-1β, IL-6 and IL-8, of the immunoregulatory cytokines IL-10 and IL-1Ra and of the corresponding pro-inflammatory TNF-α:IL-10 and immunoregulatory IL-1Ra:IL-1β ratios, all measured in an identical subset of three of the 14 experiments. For each cytokine (pg/mL) or cytokine ratio, the colour scale spans from the indicated minimum (dark green) to maximum (dark red) via median (yellow) values obtained across all macrophage subsets for the three experiments. For hierarchical classifications, the median values from the three experiments were taken into account and cytokine ratio values were excluded.
To further apprehend how polarisation shaped the cytokine response to stimulation by ACPA-IC or LPS, the pro-inflammatory IL-1β, IL-6, IL-8 and the anti-inflammatory IL-1Ra cytokines were assayed in the culture supernatants from cells of a subset of three of the 14 donors, using a multiplexed assay. The results were exclusively obtained in cells cultured in cIMDM. They are illustrated in figure 2C by heat maps and hierarchical clustering dendrograms of the median spontaneous, ACPA-IC-induced or LPS-induced secretions of all the investigated cytokines. The corresponding TNF-α:IL-10 and IL-1Ra:IL-1β ratios were also plotted with the pro-inflammatory and anti-inflammatory cytokines, respectively. Among unstimulated cells, the IFN-γ-polarised or M-CSF-polarised cells exhibited the highest spontaneous secretion of pro-inflammatory cytokines, whereas IL-4-polarised and IL-10-polarised cells provided the highest, that is, the least inflammatory IL-1Ra:IL-1β ratios. In response to ACPA-IC, the secretion of IL-1β, IL-6 and IL-8 remained roughly similar among control and IFN-γ-polarised, IL-4-polarised or IL-10-polarised cells. However, the IL-10-polarised cells provided the most anti-inflammatory IL-1Ra:IL-1β ratio and the least pro-inflammatory TNF-α:IL-10 ratio, and therefore were those which cytokine response exhibited the lowest inflammatory potential. Conversely, the M-CSF-polarised cells secreted the highest levels of IL-1β, IL-6 and IL-8 and provided the least anti-inflammatory IL-1Ra:IL-1β ratio. Moreover, as previously noted (figure 2B), they secreted the highest TNF-α concentrations and exhibited a high TNF-α:IL-10 ratio. Taken together, these data indicate that despite their phenotypic resemblance, M-CSF-polarised and IL-10-polarised cells clearly differ in their response to ACPA-IC and, in comparison with all other polarised cells, the cytokine secretion profile of the M-CSF-polarised cells exhibits the highest pro-inflammatory potential. Accordingly, hierarchical clustering clearly highlighted the M-CSF-polarised macrophages as secreting a distinctive cytokine profile. Remarkably, after stimulation by LPS, similarly to what was observed concerning TNF-α and IL-10, for IL-1β, IL-6, IL-8 and IL-1Ra, the hierarchy of responses among the differentially generated cells did not match with that observed after stimulation by ACPA-IC.
Discussion
The design of our study was largely built on the work previously published by Ambarus et al.29 Notwithstanding differences in the protocol of monocyte isolation (two successive gradient centrifugations vs gradient centrifugation followed by CD14-positive selection in our work) and in the culture conditions (adherence vs no adherence), when comparing marker expression in the IFN-γ-polarised, IL-4-polarised, IL-10-polarised cells and control cells, our study essentially confirmed previous results. Indeed, both studies identified CD80 and CD64 as markers of the IFN-γ-polarised cells, CD206 as a marker of the IL-4-polarised cells and CD32 and CD163 as markers of the IL-10-polarised cells. In addition, although we did not observe a significantly higher expression of CD16 by the IL-10-polarised cells in comparison with the IFN-γ-polarised or IL-4-polarised cells in our data set, there was a trend in that direction. The largest difference was that, despite the use of an identical antibody (Ab), we did not find that CD200R was specifically upregulated on the IL-4-polarised cells, but that high expression of this marker rather characterised all the M2 macrophages, that is, those generated in the presence of IL-4 or IL-10. This fits with the concept that CD200:CD200R interaction contributes to the anti-inflammatory properties of macrophages.30 Hierarchical clustering of the marker modifications specifically prompted by cytokines allowed three main profiles to be distinguished related to the IFN-γ-polarised cells, the IL-4-polarised cells and the cells cultured in the presence of IL-10 or M-CSF. Fitting with the report of Ambarus et al,29 high expression of CD16, CD32, CD163 and CD206 associated with the absence of CD80 showed that M-CSF-polarised macrophages (including those cultured in cSFM) were phenotypically close to the IL-10-polarised cells, which have been classified among the immunoregulatory (M2) macrophages.9
Our study assessed for the first time the influence of the macrophage polarisation on their cytokine response to ACPA-IC. Our data confirmed that the secretion of cytokines by macrophages is conditioned by their polarisation status and related to their stimulation conditions. Indeed, after LPS stimulation, the hierarchy of responses among the different macrophages was not identical with that found after ACPA-IC stimulation. Moreover, in M-CSF-cultured cells, the medium had a clear influence on the spontaneous and ACPA-IC-induced secretions of TNF-α and IL-10. The composition of the cSFM medium is not available to buyers, so we are unable to explain this phenomenon. More generally, this observation, made in a very simplified model, confirms the major influence of macrophage microenvironment on their functional properties. Finally, for IFN-γ-polarised, IL-4-polarised and IL-10-polarised cells, concerning the ACPA-IC-induced secretion of IL-6, TNF-α and IL-10 and the corresponding TNF-α:IL-10 ratio, the hierarchies between macrophages subsets matched with those reported for macrophages stimulated by immobilised IgG.31 However and interestingly, given their phenotypic classification as IL-10-like immunoregulatory macrophages, in comparison with other macrophage subsets, including the phenotypically alike IL-10-polarised cells, the M-CSF-treated cells produced the highest levels of pro-inflammatory cytokines and the ratio between their TNF-α and IL-10 secretions was similar to that produced by the IFN-γ-polarised cells. Thus, the limited phenotypic characterisation proposed by Ambarus et al29 does not allow to completely predict the macrophage function. New markers, notably allowing distinction between cells influenced by IL-10 or by M-CSF, need to be identified. Moreover, importantly, our results are in very good accordance with a recent study showing that contrary to their widely accepted classification as anti-inflammatory M2 macrophages, human M-CSF-polarised macrophages promote inflammation, instead of having immunoregulatory properties upon co-stimulation by immobilised IgG and a toll-like receptor (TLR) ligand.32 These and the above-mentioned results,31 also obtained with immobilised IgG, suggest that the results we obtained with the immobilised ACPA-IC, that we chose to study because of their strong relevance to the RA pathophysiology, might be generalisable to all immobilised IgG containing IC. However, this remains to be demonstrated as the hierarchies of the inflammatory character of the cytokine response of the different macrophage subtypes obtained with other types of IC, formed with different or additional antibody isotypes, exhibiting a different solubility, or also including ligands for receptors of pathogen-associated molecular patterns, also remain to be investigated.
The TNF-α secretion or the TNF-α:IL-10 ratio in response to ACPA-IC and the expression of FcγR were not correlated in any of the differentially polarised cells. However, we observed that in response to ACPA-IC, the IL-10-polarised and M-CSF-polarised macrophages, which are expressing the highest levels of CD32 and CD16 in comparison with IFN-γ-polarised and IL-4-polarised cells, secreted the highest levels of TNF-α. Moreover, we previously established the major contribution of the activating CD32a in the induction of TNF-α secretion by the ACPA-IC.19 The absence of correlation between expression of CD32 and ACPA-IC responses could be related to the fact that we analysed the global expression of CD32 without discriminating the stimulatory CD32a and inhibitory CD32b.
The phenotypic and functional properties of monocytes and macrophages have been largely explored in many animal models of tissue injury, including cancer,33 atherosclerosis34 and, quite recently, IC-induced arthritis.35 In this study, Ly6C-negative monocytes were found to be crucial for arthritis initiation, giving rise first to recruited inflammatory macrophages, then changing their phenotype from M1 to M2 in situ to permit resolution of the disease. Phenotypic and functional analyses of human tissue macrophages in diseases such as RA are still scarce. The lack of firmly established markers for the classification of polarised human macrophages and the extreme plasticity of each polarisation following changes in the environmental conditions make this investigation quite difficult. However, a recent transcriptome network analysis, based on 299 human polarised macrophages, will help first to understand how macrophages integrate signals from their microenvironment under inflammatory conditions, and second to define polarisation states of macrophages in diseased tissues.36 However, the idea that the M-CSF-induced macrophage phenotype is common in RA joints is sustained by strong circumstantial evidence. Indeed, despite the current difficulties, the use of the specific markers of human macrophages described by Ambarus et al29 has allowed a preliminary appraisal of the polarisation status of synovial macrophages in RA and spondyloarthritis.37 Histological analyses of biopsies showed that sublining macrophages had a heterogeneous phenotype because they co-expressed markers found on in vitro IFN-γ-polarised, IL-4-polarised or IL-10-polarised cells. In contrast, intimal lining layer macrophages revealed a strong co-expression of CD163 and CD32, leading the authors to propose that intimal macrophages displayed an IL-10-polarised-like phenotype.37 Since our data have highlighted the phenotypic overlap between the IL-10-exposed and the M-CSF-exposed cells, it could alternatively be proposed that the expression of these two markers results from an exposure to M-CSF. Fitting with this hypothesis, another immunohistochemical analysis has shown that in the lining and sublining areas of the synovium of patients with RA, a huge majority (90–85%) of the macrophages that express CD163 (upregulated by M-CSF or IL-10) are TNF-α-expressing cells, while a smaller proportion (70–35%) express IL-10.38 In addition, M-CSF is highly expressed in the RA ST and suspected to play a major role in the disease.39 ,40 The pro-inflammatory cytokines IL-1 and TNF stimulate the production of M-CSF by human fibroblast-like synoviocytes and by chondrocytes41 ,42 and synovial endothelial cells from patients with RA also produce M-CSF.43 Both in experimental arthritis and in RA, in association with the receptor activator of nuclear factor-kappa B (NF-kB) ligand (RANKL), M-CSF contributes to differentiation of synovial macrophages into osteoclasts.44 ,45 Furthermore, M-CSF exacerbates collagen-induced arthritis (CIA). Conversely, its neutralisation reduces the severity of established CIA.46 Finally, recent data support the evidence that blockade of the M-CSF receptor reduces inflammation in CIA and abrogates cartilage damage and bone erosion.47 Moreover, inhibitors of M-CSF and its receptor are in preclinical development for the treatment of RA and other autoimmune diseases.47 ,48
We previously demonstrated the pro-inflammatory potential of ACPA-IC using M-CSF-generated macrophages from healthy individuals and patients with RA.19 ,49 To better appraise the pathophysiological impact of these observations, it would have been ideal to carry out ACPA-IC stimulation on macrophages isolated from rheumatoid ST samples since their response to such stimulation, most probably variable depending on their localisation (intimal vs subintimal synovium) or of their origin (tissue-resident vs recruited macrophages), remains undescribed. Our first approach was to evaluate the influence of macrophage polarisation on the response to ACPA-IC using different polarised cells. It could be used as a reference when the RA-associated phenotype(s) of ST macrophages will have been better described. Nonetheless, we show for the first time that the cytokine response to ACPA-IC of M-CSF-polarised macrophages is the most pro-inflammatory in comparison with macrophages generated in the presence of IFN-γ, IL-4 or IL-10. Given the abundance of this cytokine, M-CSF-influenced macrophages are probably numerous in the RA synovium. Fitting with this hypothesis, the expression of surface markers known to be increased by M-CSF such as CD16, CD32 or CD163 has been described on RA synovial macrophages.50–54 Upon chronic exposure to ACPA-IC, the M-CSF-influenced macrophages likely mount a pro-inflammatory cytokine response which probably contributes to the synovitis.
Acknowledgments
The technical assistance of M.-F. Isaïa, M.-P. Henry, G. Offer and E. Parra is gratefully acknowledged. We also thank A. Desquesnes and L. Monbrun (Phenotype Analysis service of the Anexplo Platform in Life Sciences of Toulouse Genopole) for performing multiplex cytokine assays, and F.-E. L'Faqihi-Olive and V. Duplan-Eche (Cytometry and Cell Sorting service of the Toulouse RIO Imaging platform) for assistance in flow cytometry analyses.
References
Footnotes
Handling editor Tore K Kvien
GS and MS contributed equally.
Contributors CC, MS, FA and GS contributed to conception or design of the study. CC, LC and FA contributed to acquisition of data. CC, LC and MS contributed to their analysis and interpretation. CC and MS wrote the article and revised the final version. GS revised the article. All authors approved the submitted version.
Funding This study was supported by grants from the Toulouse III University, the ‘CNRS,’ the ‘INSERM’ and the ‘Société Française de Rhumatologie’.
Competing interests None declared.
Patient consent Obtained.
Provenance and peer review Not commissioned; externally peer reviewed.
Author note: Current addresses for FA : La Jolla Institute for Allergy and Immunology, La Jolla, California, USA; MS: Institut de Recherche en Santé Digestive (IRSD), U1220 INSERM-INRA-ENVT-Université Toulouse, France