Article Text

Abstract
Targeting chemokines and/or chemokine receptors appears to be an intriguing new approach to treating chronic inflammatory disorders like rheumatoid arthritis, inflammatory bowel diseases, multiple sclerosis, and transplant rejections. The involvement of chemokines and chemokine receptors in inflammatory joint diseases, the in vitro and in vivo characteristics of the chemokine family in inflammatory joint disease, and initial clinical data on chemokine blockade in patients with rheumatoid arthritis suggest that targeting the chemokine and chemokine receptor family might provide a new, promising antirheumatic strategy.
- IL, interleukin
- MCP, monocyte chemoattractant protein
- OA, osteoarthritis
- PsA, psoriatic arthritis
- RA, rheumatoid arthritis
- SF, synovial fluid
- ST, synovial tissue
- TNFα, tumour necrosis factor α
- rheumatoid arthritis
- chemokines
- synovium
Statistics from Altmetric.com
- IL, interleukin
- MCP, monocyte chemoattractant protein
- OA, osteoarthritis
- PsA, psoriatic arthritis
- RA, rheumatoid arthritis
- SF, synovial fluid
- ST, synovial tissue
- TNFα, tumour necrosis factor α
Rheumatoid arthritis (RA) is a chronic inflammatory disease that affects synovial tissue (ST) in multiple joints. Although its aetiology is still unknown, RA is thought of as an autoimmune disease. Severe morbidity and structural damage of joints caused by chronic inflammation requires early and effective treatment.1 The inflammatory process is characterised by infiltration of leucocytes in ST and the synovial fluid (SF) in the joints. A wealth of information now exists demonstrating that this process is mediated by the coordinated action of a variety of inflammatory mediators. Leucocytes adhere to the activated endothelium, leading to migration across the endothelial barrier into the ST.2 A complex network of adhesion molecules and chemokines coordinate cell migration, by working in concert to induce an inflammatory response.3,4 Investigators have recently extended initial observational studies to explore the role of chemokines and chemokine receptors in the pathogenesis of inflammatory joint disease.
As the clinical management of RA is difficult, there is a need for the development of a broader range of treatments targeting specific points of the complex pathogenetic network underlying RA. The chemokine and chemokine receptor families may provide intriguing targets for innovative treatment of RA and other chronic inflammatory disorders by the use of biological agents or targeted small molecules.
CHEMOKINES
Chemokines are a specialised family of small (8–10 kDa) structurally related proteins that function as potent mediators of inflammation by their ability to recruit and activate specific leucocyte subpopulations.5,6 The members of the chemokine family can be distinguished from classical chemoattractant molecules (for instance, complement fragment peptides C3a and C5a, and lipid molecules such as leucotriene B4 and platelet activating factor) by their shared structural similarities. Chemokines have four conserved cysteine residues that form disulphide bonds, which are critical for the tertiary structures of these proteins.7,8
The chemokine family is organised into four subfamilies according to the position of the first two cysteines. The two major subclasses include the CC chemokines where the cysteines are adjacent and the CXC chemokines where the cysteines are separated by one amino acid. The CXC chemokines mainly act on neutrophils and lymphocytes, whereas the CC chemokines mainly act on monocytes and lymphocytes without affecting neutrophils.6
At least two chemokines do not fit into this classification. Lymphotactin is a chemokine, which lacks cysteines one and three of the typical chemokine structure (C-) and is a potent attractant for T lymphocytes, but not for monocytes.9 Fractalkine is a membrane bound glycoprotein, in which the first two cysteine residues are separated by three amino acids (CXXXC).10 These two chemokines may represent additional families.
“Four chemokine subfamilies are mediators of inflammation”
The discovery that certain chemokine receptors (CCR5 and CXCR4) are important co-receptors for HIV entry in lymphocytes gave a strong impulse to both basic research and to the development of new therapeutic agents targeting chemokine receptors.
One of the most commonly suggested characteristics of chemokines is the redundancy of the system. Redundancy occurs by having multiple ligands for each chemokine receptor and multiple receptors for each chemokine ligand, leading to the generation of multiple pathways directing similar cellular responses. This is discussed in more detail below.
NOMENCLATURE OF CHEMOKINES
To date more than 50 different human chemokines have been identified. Initially, chemokines were named according to their distinctive functions, for instance monocyte chemoattractant protein (MCP-1) or regulated upon activation normal T cell expressed and secreted (RANTES). The fact that the same chemokine was given multiple names by different groups (for instance HCC-2 = Lkn-1 = MIP-1δ) made the already large and complex family even more complex. Moreover, additional research has shown that some chemokines have more functions than initially described and, therefore, their original names might suggest limited function. For instance MCP-1, a CC chemokine initially identified as a monocyte-specific chemoattractant, has also been shown to attract activated T cells, NK cells, and basophils.11 Besides attracting inflammatory cells, MCP-1 also plays a part in T cell differentiation and angiogenesis.12,13
Therefore, a new nomenclature has been proposed. The system is based on the structural homology of different subgroups. Thus, chemokines can be divided into CXC, CC, CXXXC, and XC chemokines. For the ligands an L and a number are added (for example, MCP-1 is CCL2) and for the receptors an R and a number (for example, CCR1, CCR2 etc.)(table 1⇓).14–,16
CXC, C, CX3C, and CC chemokine/receptor families. Adapted/modified from OpenUrlCrossRefPubMedWeb of Science
HOMOEOSTATIC CHEMOKINES IN RA
Chemokines promote the accumulation of leucocytes at the source of chemokine production in multiple ways (fig 1⇓). This can be part of a homoeostatic process or an inflammatory reaction. Chemokines can be grouped into either homoeostatic/constitutive chemokines or inflammatory/inducible chemokines, although some may have a dual role depending on how they are produced.17–,19
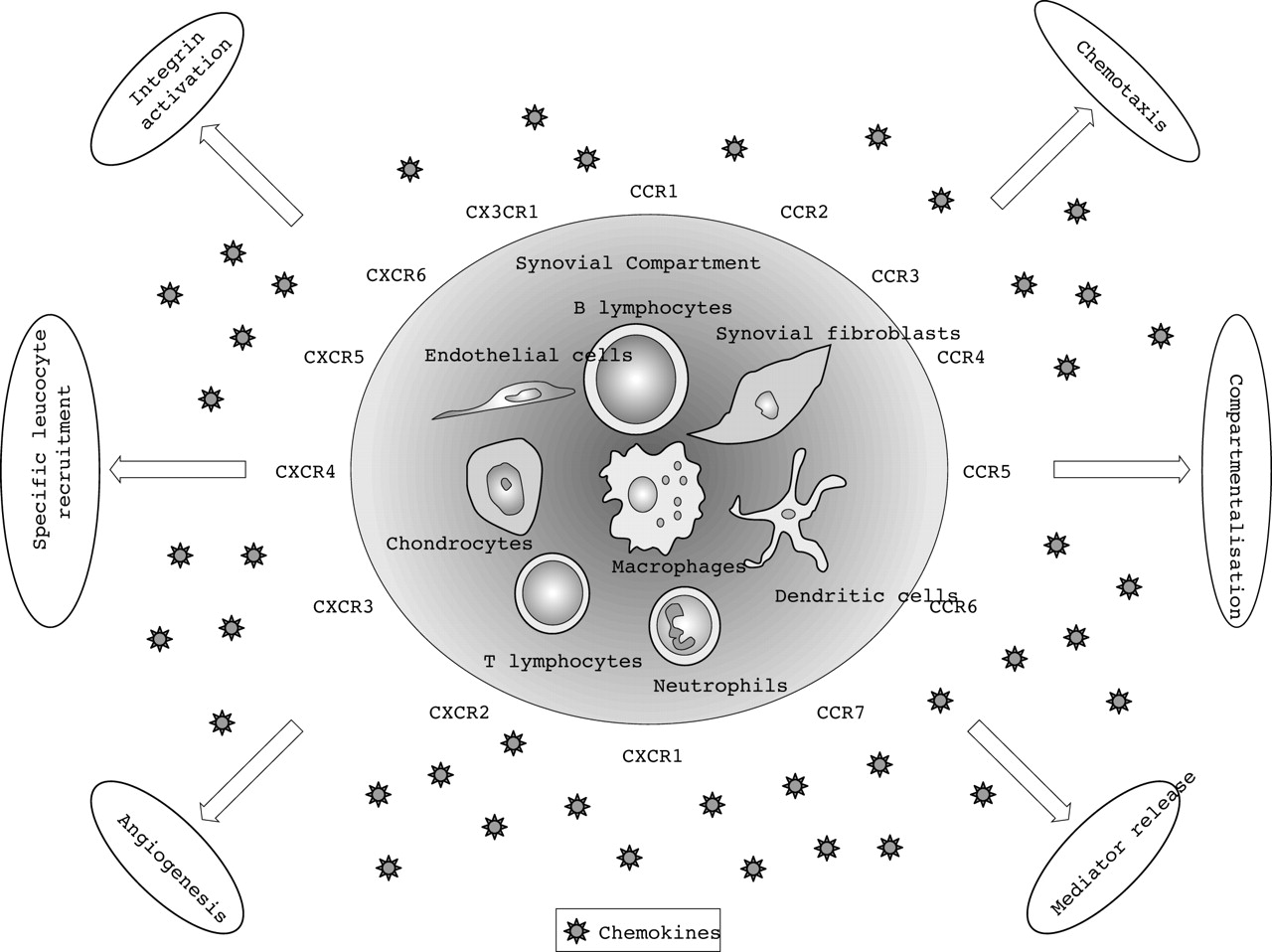
Overview of homoeostatic and inflammatory functional interactions of chemokines and their receptors in inflammatory joint disease.
Homoeostatic chemokines are produced within the lymphoid tissues and they are involved in maintaining homoeostatic leucocyte traffic and cell compartmentalisation in these tissues under non-inflammatory conditions.20,21 They also assist in controlling the specificity of memory cell subsets in specific tissues.22,23 In RA the expression of certain homing chemokines in the inflamed synovium is associated with the formation of lymphocyte aggregates resembling germinal centres in lymphoid tissue.24 These structures can be found in about 20–30% of patients with RA.25,26 Although research describing expression of homoeostatic chemokines and their receptors in inflamed ST is still limited, some homing chemokines like CXCL13/BCA-1, the ligand of CXCR5, have been implicated in the formation of these ectopic lymphoid structures in rheumatoid ST. CXCL13/BCA-1 expression was detected predominantly in follicular dendritic cells in germinal centres.27
“Chemokines promote leucocyte accumulation in many ways”
For homing of immature dendritic cells into RA synovium, the ligand–receptor pair CCL20/MIP-3α–CCR6 appears to be critical. These cells are especially found in the intimal lining layer and perivascular infiltrates.28 CCL20/MIP-3α can be detected at higher concentrations in SF of patients with RA than in controls. It is also a chemoattractant for T cells, B cells, and monocytes.29,30 A recent report showed that immunodepleting CCL20/MIP-3α resulted in an overall inhibition of monocyte chemotaxis of 40%.30 The homing of mature dendritic cells to the lymphocyte infiltrates is influenced by CCL19/MIP-3β and CCL21/6-chemokine, the ligands for CCR7. They are expressed in the perivascular lymphocyte infiltrates and the vascular endothelium and are thought play a part in the migration of CCR7+ mature dendritic cells into the synovium.28
Another homoeostatic receptor, CXCR4, may have a role in the active retention of T cells within the synovial compartment; its only ligand known so far, CXCL12/SDF-1, is expressed by synovial endothelial cells and synovial fibroblasts.31,32 Moreover, CXCL12/SDF-1 co-localises with the neoangiogenesis marker αvβ3 integrin and can modulate angiogenesis in in vivo assays in mouse models.33
The ligand/receptor pair CXCL16–CXCR6 seems to be especially involved in plasma cell recruitment to bone marrow and target tissues.34 Expression of CXCR6 has been described in RA and could also attract effector T cells, mediating Th1 responses.35
As homoeostatic chemokines and their receptors may play an important part in the continuation of inflammation by the formation of germinal centres and the sustained retention of cells in the synovial compartment, targeting these proteins might be an interesting new approach to the treatment of RA. Additional studies are needed to determine the therapeutic potential and safety of this approach. A concern might be that one does not want to interfere with the role of these molecules under non-inflammatory, physiological conditions. Clearly, most suitable potential targets still need to be defined.
INFLAMMATORY CHEMOKINES IN RA
In contrast with the homoeostatic chemokines, which are mainly involved in physiological processes, although some of them seem to play a dual role in inflammation, the inflammatory/inducible chemokines are preferentially produced in peripheral tissue at the sites of inflammation. At these sites they function to recruit neutrophils, monocytes, immature dendritic cells, B cells, and activated T cells.5,6 Several cell types, including endothelial, epithelial, and stromal cells as well as leucocytes, can produce inflammatory chemokines. Although many different cell types have the ability to produce inflammatory chemokines in the synovium, macrophages and fibroblast-like synoviocytes seem to be the most potent producers based upon in vitro experiments.36–,39 Chemokine production is induced at high levels in response to inflammatory stimuli, such as lipopolysaccharide, interleukin (IL)1, and tumour necrosis factor α(TNFα). The inflammatory chemokines promote inflammation by an array of biological mechanisms13(fig 1⇑).
Leucocyte extravasation involves the sequential steps of selectin dependent initial attachment and rolling, G protein regulated activation, firm arrest through integrin binding to endothelial ligands, and transendothelial diapedesis.3,4,40 Chemokines participate in this process by providing a chemokine gradient and by activation of integrins.13
CXCL8/IL8 was among the first chemokines shown to be involved in leucocyte chemotaxis.41 In patients with RA, CXCL8/IL8 is present in high quantities in both ST and SF.42,43 Of interest, CXCL8/IL8 protein and mRNA levels are specifically increased in clinically affected joints compared with clinically unaffected joints, suggesting a role in the development of clinical signs and symptoms.44 The notion that CXCL8/IL8 may be involved in the pathogenesis of RA is supported by the observation that a single intra-articular injection of CXCL8/IL8 can initiate arthritis in a time and dose dependent manner in rabbits. This is associated with an early increase in neutrophil numbers followed by an increase in mononuclear cells in the joints of these animals.43 In another study CXCL8/IL8 was also shown to have a major role in the trafficking of monocytes towards the ST.45
CCL2/MCP-1, a ligand of CCR2, has also been shown to have important chemotactic characteristics. CCL2/MCP-1 promotes transmigration of monocytes when present in a (soluble) gradient across an endothelial monolayer.46 CCL2/MCP-1 can also attract T cells, NK cells, and basophils.47,48 ST macrophages are the dominant source of CCL2/MCP-1 production in RA, and levels of CCL2/MCP-1 are significantly increased in RA ST, SF, and peripheral blood.49 The possible role of CCL2/MCP-1 in monocyte migration towards the joint is supported by the observation that injection of CCL2/MCP-1 into rabbit joints leads to a clear increase in, especially, macrophage numbers.50
CCL5/RANTES (a ligand for CCR1, CCR3, and CCR5) is another chemokine implicated in leucocyte chemotaxis in RA. In RA fibroblast-like synoviocytes express CCL5/RANTES, and CCL5/RANTES gene expression is increased in SF mononuclear cells and peripheral blood mononuclear cells.51–,53 CCL5/RANTES proved to be a potent inducer of migration of both monocytes and T lymphocytes in in vitro models.6 By blocking CCL5/RANTES using neutralising antibodies it was shown that in vitro migration towards SF and ST from patients with RA could be inhibited.54,55
CCR5, one of the receptors for CCL5/RANTES, is together with CXCR3 predominantly (but not exclusively) expressed by Th1 cells. Thus, this receptor could have a crucial role in a Th1-type disease like RA.56–,59 Examination of the expression of the ligands of CCR5 and CXCR3 in RA ST, SF, and peripheral blood showed that there is a chemotactic gradient between the serum and synovium for CCL3/MIP-1α, CCL4/MIP-1β, CXCL9/Mig, and CXCL10/IP-10, favouring migration into tissue.60,61
Other studies showed the importance of CCL3/MIP-1α in neutrophil and macrophage chemotaxis into the synovial compartment.62 Besides a ligand for CCR5, this is also a ligand for CCR1. In addition to attracting neutrophils and macrophages CCL3/MIP-1α is also involved in attracting lymphocytes to SF, although lymphocyte migration seems to be dependent on multiple chemokines.63
The ability to provide chemokine gradients and the ability to bind to synovial structures may vary between different chemokines. Only a subset of the chemokines appears to bind efficiently to structures in the extravascular space, which may be dependent on their glycosaminoglycan binding capacity, indicating that the formation of chemotactic gradients in tissues may be very complex.64 Although blockage of single chemokines may inhibit migration, the combination of several antibodies targeting different chemokines seems to be more effective in reducing the migratory activity towards the synovial compartment.63
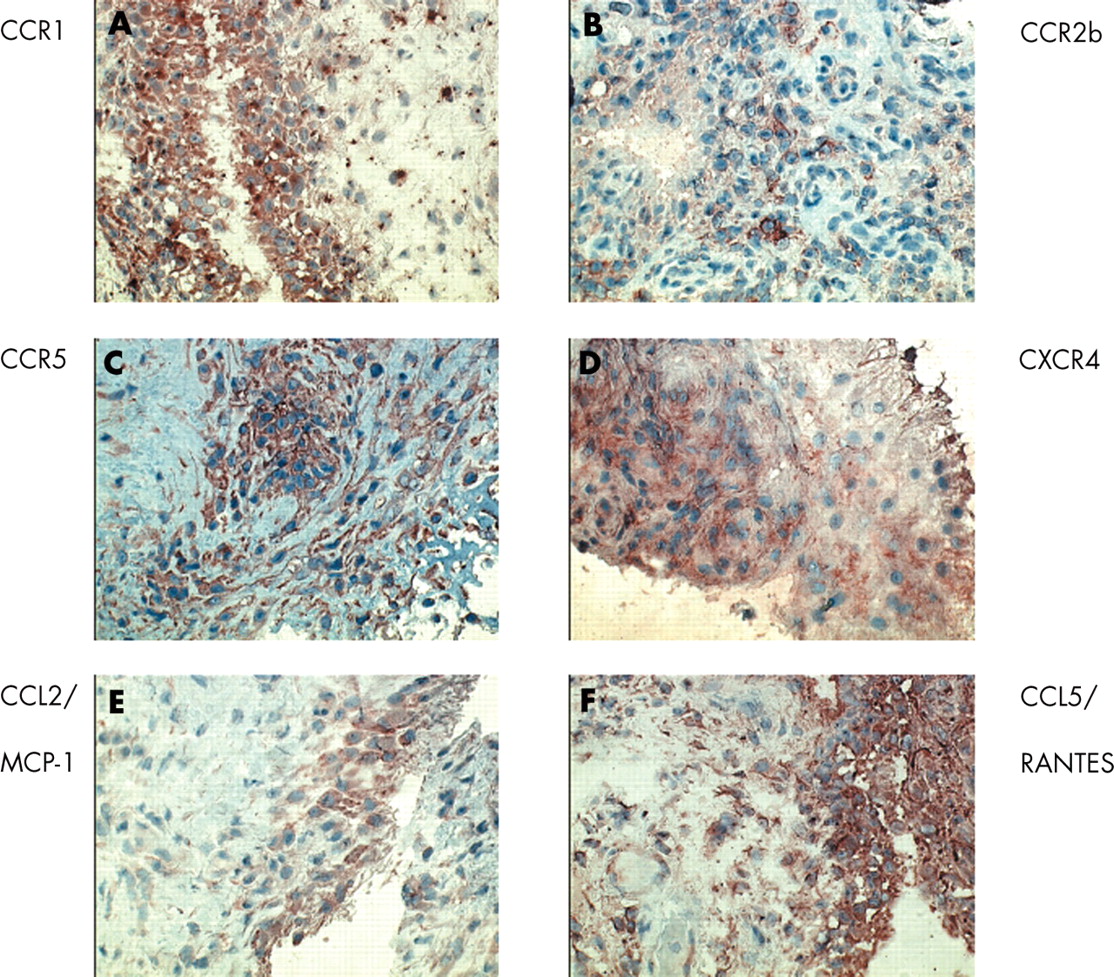
Besides providing a functional gradient, chemokines can also promote chemotaxis by inducing a rapid but transient increase in adhesion of leucocytes to immobilised integrin ligands.65,66 This can occur by two different mechanisms: up regulation of integrin affinity through conformational changes and changes in integrin avidity that occurs through integrin clustering.65,67 Using in vitro flow chamber assays, it was shown that some of these chemokines could up regulate integrin function and then induce arrest and firm adhesion of rolling cells. For example, CXCL8/IL8 can induce adhesion of CXCR1 and CXCR2 expressing neutrophils; CCL2/MCP-1 may trigger adhesion through its receptor CCR2, and different memory lymphocyte subsets can adhere in response to ligands for CXCR3 (CXCL9/Mig and CXCL10/IP-10).68–,70Figure 2⇓ shows the expression of CCR1, CCR2b, CCR5, CXCR4, CCL2/MCP-1, and CCL5/RANTES in the ST of a patient with RA.

{kind=link}
{kind=link}
Expression of chemokine and chemokine receptor staining in RA ST. CCR1 positive cells are scattered throughout the synovium, and are expressed predominantly by macrophages. There is marked CCR1 expression in the intimal lining layer. CCR2b is expressed by macrophages, especially in the synovial sublining. CCR5 is expressed by both T lymphocytes and macrophages. CXCR4 is mainly expressed by T lymphocytes in the synovial sublining. CCL2/MCP-1, a ligand for CCR2b is almost exclusively expressed in the intimal lining layer. CCL5/RANTES, a ligand for CCR1, CCR3, and CCR5 is expressed in both the intimal lining layer and the synovial sublining. (Single stain peroxidase technique, positive staining in red/brown, Mayer’s haematoxylin counterstained, original magnifications ×400).
CYTOKINE–CHEMOKINE INTERACTIONS
There is a complex relationship between inflammatory cytokines and chemokines.
Multiple studies have shown that a vast number of proinflammatory cytokines, like IL1, IL2, interferon γ, and TNFα can stimulate chemokine production by different cell types present in rheumatoid ST.36,71–,73 Macrophages may produce a broad variety of chemokines, including CC and CXC chemokines like CXCL8/IL8, CCL2/MCP-1, and CCL3/MIP-1α.42,49,62 In addition, lymphocytes and fibroblast-like synoviocytes can produce various chemokines upon appropriate stimulation.36,37,74 Conversely, chemokines may also stimulate cells to release inflammatory mediators including cytokines. Stimulation of fibroblast-like synoviocytes from patients with RA with CCL2/MCP-1, CCL5/RANTES, and CXCL12/SDF-1 results, for instance, in enhanced production of IL6 and IL8.75 Similarly, CCL3/MIP-1α has been shown to stimulate the synthesis of IL1, TNF, and IL6 by murine peritoneal macrophages.76
A relationship between chemokines and matrix metalloproteinases (MMPs) has also been described. The interaction between CXCL12/SDF-1 and its receptor CXCR4 on chondrocytes may result in increased secretion of MMP-3, a cartilage matrix degrading enzyme.77 CCL2/MCP-1, CCL5/RANTES, and CXCL1/GROα were also shown to stimulate MMP-3 release by chondrocytes isolated from osteoarthritis (OA)articular cartilage samples.78
Although there is clear evidence that chemokines can stimulate various cell types in the joint to produce cytokines and MMPs, promoting synovial inflammation and joint degradation, the data are still limited.
ANGIOGENESIS
Angiogenesis, the process by which new capillaries are formed from existing vessels, is fundamental to a variety of physiological activities, such as wound healing and the female reproductive system. In addition, angiogenesis plays a part in pathological processes, including tumour growth and chronic inflammation.79 In RA, neovascularisation is one of the early changes in the ST, and in this complex, but coordinated, process chemokines are intimately involved. Chemokines may have either angiogenic or angiostatic properties. Initially, for CXC chemokines it was shown that this was dependent on the so called ELR motif, which is the sequence of three amino acids (Glu-Leu-Arg) near the NH2 terminus before the first cysteine. Chemokines containing this motif were thought to be angiogenic, whereas chemokines without this motif appeared to be angiostatic.80 CXCL8/IL8 was the first chemokine with an ELR motif shown to have angiogenic properties. Other ELR-containing CXC chemokines with angiogenic features include CXCL5/ENA-78 and CXCL1/GROα, whereas non-ELR chemokines like CXCL4/PF4 and the CXCR3 ligands CXCL9/Mig, CXCL10/IP-10, and CXCL11/I-TAC are angiostatic.80–,83 Although studies have shown that the CXCR3 ligands function as angiostatic mediators, in high concentrations they could in vitro also induce endothelial cell chemotaxis.84 There are more exceptions to the “ELR” rule, like the ELR negative chemokines CXCL12/SDF-1 and CXCL2/GROβ, which do have angiogenic features.85,86 Other chemokines with angiogenic properties lacking the ELR motif include the CC and CX3C chemokines CCL2/MCP-1 and CX3CL1/fractalkine.87–,89
It is also possible that the angiogenic or angiostatic involvement does not depend on the chemokine ligand, but on its receptor. Endothelial cells may express functional chemokine receptors, which can induce angiogenic signals, like CXCR2 and CXCR4. Alternatively, they may express receptors like CXCR3, which induce angiostatic signals.83,90 Although the role of chemokines and their receptors in angiogenesis is as yet not completely understood, it has become clear that they are not only involved in stimulating inflammation, but may also have inhibiting features.
CHEMOKINES IN JOINT DISEASES OTHER THAN RA
Although the vast majority of research on chemokines in inflammatory joint disease is focused on RA, it seems clear that chemokines and their receptors also have a role in other inflammatory arthropathies like OA and psoriatic arthritis (PsA).
In OA, traditionally considered a non-inflammatory arthropathy, synovial inflammation has been well documented.91–,93 There is accumulating evidence suggesting the involvement of chemokines and their receptors in the disease process. Among others, CCL2/MCP-1 and CCL5/RANTES are expressed in both SF and ST of patients with OA.94–,96 Fibroblast-like synoviocytes from patients with OA have the capacity to produce CXCL8/IL8, CCL2/MCP-1, CCL3/MIP-1α, and CCL5/RANTES.37,54,97 Although most of the described chemokines and receptors appear to be expressed at higher levels in ST and SF in RA than in other inflammatory arthropathies, there might be at least one exception—namely, CCL4/MIP-1β. It is found at significantly greater levels in the SF of patients with OA than in other forms of inflammatory arthritis like RA.98 CCL4/MIP-1β, which is a ligand for CCR5, may be responsible for a substantial fraction of OA SF chemotactic activity for monocytes, particularly for activated monocytes.
In addition, chemokines and chemokine receptors are expressed by chondrocytes and, as mentioned earlier, interaction between chemokine receptors and their ligands may cause MMP-3 induction in chondrocytes.78,99,100 Compared with RA and PsA, OA cartilage appears to exhibit higher expression of the chemokine receptors CCR1, CCR3, CCR5, CXCR1, CXCR2, and CXCR3, but still lower than the cartilage of normal donors.101
In PsA CCL2/MCP-1, CCL5/RANTES, CXCL1/GROα, CXCL8/IL8, CXCL9/Mig, as well as CCR2 and CCR5, have been described in both ST and SF.95,102–,104 CCR2 and CCR5 were also detected in the SF of patients with ankylosing spondylitis, reactive arthritis, Lyme arthritis, and gout.103
Evidently, chemokines and their receptors are present in the peripheral blood, synovial compartment, and cartilage in patients with arthritides other than RA. Although the level of expression may vary between the different forms of arthritis, none of the inflammatory chemokines or receptors appears specific for any of the conditions; presumably their presence reflects the effector mechanisms involved in the inflammatory process. This suggests that inhibition of the chemokine repertoire might not only be a useful strategy in the treatment of RA but also in other forms of inflammatory joint disease.
ANIMAL MODELS
In addition to in vitro studies suggesting the potential of reducing cell migration by inhibition of chemokines and chemokine receptors, animal models have been used to assess the value of the chemokine inhibitors in vivo. As animal models can be used to assess the role of several pathogenic factors at various stages of the disease, they provide a sophisticated tool for studying the relevance of chemokines and chemokine receptors.
Various chemokines have been described in arthritis models. MIP-2, the murine equivalent of CXCL8/IL8 has been detected in the joints of mice with collagen induced arthritis in the early stage of disease, whereas other chemokines like CCL5/RANTES and CCL3/MIP-1α were present in early as well as late disease stages.105 In the rat adjuvant arthritis model it was shown that among different cytokines, the chemokines CCL3/MIP-1α and CXCL5/ENA-78-like protein are involved in the onset of arthritis and JE (CCL2/MCP-1) is involved in late phases of the disease.106
Chemokines and their receptors can be targeted in different ways, using neutralising antibodies, modified chemokines or small molecule receptor antagonists. Examples of targeted chemokines and receptors in animal models of arthritis include CCL2/MCP-1, CCL5/RANTES, CXCL5/ENA-78, CXCL10/IP-10, CXCR2, CXCR4, CCR2, and CCR5.107–,115 Targeting CCL2/MCP-1 in rats or its receptor CCR2 in mice showed that both approaches reduced arthritis activity. In both models there was a reduction in macrophage infiltration and protection against joint destruction.107,108 Treatment of rat adjuvant arthritis with a polyclonal antibody directed against human CCL5/RANTES resulted in clinical improvement and reduced cell infiltration and joint damage, while antibody treatment directed at CCL3/MIP-1α in the same model did not influence the disease.109 It has also been shown that targeting the CXC chemokine CXCL10/IP-10 may inhibit leucocyte migration and induce an alteration in the Th1/Th2 balance in rats.111 Another successful example of chemokine blockade using antibodies is antihuman ENA-78 antibody treatment in rat adjuvant arthritis, which targets the CXC ligand CXCL5/ENA-78 like protein. This approach was effective when given before disease onset, but it could not modify the disease when administered after the development of clinical signs of arthritis.112
Of interest, several synthetic, non-peptide, highly specific chemokine receptor antagonists have been reported to be effective in animal models. AMD3100, a specific antagonist of the CXCR4 receptor, could delay the onset of arthritis and reduce the severity of symptoms in mice. Treatment with AMD3100 also led to a decrease in infiltration by mononuclear cells and neutrophils and a reduction in synovial tissue hyperplasia and pannus formation.113 Similarly, treatment with a selective non-peptide antagonist of CXCR2 (the receptor of CXCL8/IL8) in a rabbit arthritis model resulted in reduced neutrophil, monocyte, and lymphocyte counts in SF.114 Finally, a non-peptide CCR5 antagonist resulted in reduction of both incidence and severity of collagen induced arthritis by modulating T cell migration.115
Many of these studies clearly suggest that targeting migration of leucocytes by chemokine blockade might be feasible and that this approach could be used to ameliorate clinical signs of arthritis and inhibit joint destruction. Although in most reported studies chemokine blockade prevented arthritis, it appears to be more difficult to reduce established disease.112 Taken together, the available evidence suggests that specific inhibition of one chemokine or one chemokine receptor may have relevant clinical and biological effects in animal models, despite the redundancy of the system. Obviously, however, promising results in animal models need to be confirmed in human disease.
HUMAN STUDIES
Chemokine receptors are G protein coupled, seven transmembrane spanning receptors. Several small molecule antagonists targeting these receptors have recently been developed.116,117 To date only a few publications have described the effects of chemokine or chemokine receptor antagonism in patients with chronic inflammatory disease. It has been suggested in a review article118 that there was lack of efficacy of treatment in a phase II study using a monoclonal antibody against CXCL8/IL8 in patients with RA, but the full dataset has as yet not been published.
Recent work suggested the potential of treatment with a CCR1 antagonist in patients with RA.116,119 In a relatively small phase Ib study, patients were treated with a potent and selective antagonist for CCR1 for 2 weeks, which was well tolerated.119 Analysis of ST samples showed a clear reduction in synovial inflammation in the active treated group compared with the placebo group. Cells capable of expressing the CCR1 receptor were affected, whereas cells unable to express this receptor were not influenced by the treatment, confirming the specificity of the compound.
Although this study was not designed to determine clinical efficacy, one third of the patients in the active group fulfilled the ACR20 criteria for improvement (including one patient with an ACR50 response) already after 2 weeks of treatment, whereas there were no responders in the (small) placebo group. This study is the first in humans showing that chemokine antagonism might be feasible and suggesting that it is indeed possible to influence migration of inflammatory cells in a chronic inflammatory disease using a specific orally available chemokine receptor antagonist. Obviously, the clinical relevance needs to be confirmed in larger, well controlled clinical trials. The study with the CCR1 antagonist adds to the rationale for such trials.
REDUNDANCY OF THE CHEMOKINE SYSTEM
Based upon in vitro data one might conclude that most of the chemokines have redundant or similar functions compared with other chemokines, although particular receptors are expressed by specific cells (for instance CXCRs mainly by neutrophils, and CCRs mainly by monocytes and lymphocytes). Examination of the in vivo data in animal models showed, as discussed above, that the production of chemokines is coordinated and that different chemokines function at distinct stages of disease evolution. Data from knockout models showed the effects of depletion of one particular chemokine or chemokine receptor—for instance, CCR1 knockout mice were partially protected against experimental allergic encephalomyelitis compared with CCR1+/+ mice.120
The differences in the dynamic interactions between a ligand and receptor also imply distinct functions for different receptors. For instance CXCR1 and CXCR2, both expressed on neutrophils and both serving as receptor for CXCL8/IL8, respond differently to CXCL8/IL8 binding. CXCR1 is slowly internalised and rapidly recycled after binding, whereas CXCR2 is rapidly internalised, targeted for degradation, and reappears only slowly at the membrane after ligand activation.121
Many chemokines and chemokine receptors are expressed within the synovial compartment of patients with RA, which might suggest redundancy in chronic synovial inflammation. It should be noted, however, that differences in expression may also suggest distinct pathogenetic functions for various ligands and receptors. CCR1, for example, appears to be involved in the initial recruitment of monocytes from the circulation to the sites of inflammation, whereas CCR5 may be more important in the retention of cells within the synovial compartment.122
Increasing evidence suggests that the chemokine system functions in a delicately controlled manner with distinct as well as overlapping functions for individual chemokines and receptors, depending on the stage of the disease. The data from blocking studies in animal models provide evidence that redundancy of the system does not exclude beneficial clinical and biological effects of specific chemokine antagonism. On the other hand, it might even provide a safe background for therapeutic application of highly specific antagonists. The first report of the effects of treatment with a specific CCR1 antagonist in RA confirms that inflammation might be decreased safely by specifically targeting one chemokine receptor.119
CONCLUSION
Clearly, a very sophisticated system exists, controlling the movement of leucocytes under physiological and pathological conditions. Chemokines and their receptors have an important role in this system not only by directing the migration of cells but also by controlling other important mechanisms like release of mediators of inflammation and angiogenesis. Despite the redundancy of the system, animal studies have shown the potential effect of specific antagonism of ligands and receptors in disease models, with apparently few side effects. The first human study with a small molecule serving as a chemokine receptor antagonist was recently performed in patients with RA and showed clear biological efficacy. Although biological treatments using neutralising antibodies or modified chemokines as receptor antagonists might provide effective treatments, the ideal therapeutic agent may be a small molecule antagonist which can be administered orally.
There are several caveats, however, that need to be recognised. Conceivably, it is necessary to use polychemokine antagonists117 or combine different chemokine antagonists63 to block specific chemokine pathways. Additionally, it is possible that some ligands may act as agonists at one receptor and as antagonists at others.123
In conclusion, although several questions remain to be answered, chemokine blockade may be a promising, innovative approach for treating chronic inflammation.