
Abstract
Objective. Regulatory T cells are characterized by expression of the transcription factor FoxP3 and are thought to be involved in the pathogenesis of autoimmune diseases. We determined the frequency and phenotypic characteristics of CD4+FoxP3+ T cells in the blood and synovial fluid (SF) of patients with inflammatory joint diseases.
Methods. SF from 10 patients with ankylosing spondylitis (AS), 20 patients with other spondyloarthritides or with peripheral arthritis (pSpA), and 12 patients with rheumatoid arthritis (RA), and peripheral blood (PB) from 22 patients with AS, 19 with pSpA, 15 with RA, and 12 healthy controls were stained for CD4, FoxP3, CD25, and CD127 and different effector cytokines and then analyzed by flow cytometry. Methylation pattern of the Treg-specific demethylated region (TSDR) was determined after bisulfite treatment by quantitative polymerase chain reaction.
Results. In all groups of patients we observed higher frequencies of Foxp3+ cells/CD4+ T cells within SF compared to PB. The frequency of synovial Foxp3+ cells/CD4+ T cells was significantly higher in patients with pSpA (18.79% ± 6.41%) compared to patients with AS (9.69% ± 4.11%) and patients with RA (5.95% ± 2.21%). CD4+FoxP3+ T cells were CD25+ and CD127− and lacked effector cytokine production in any of the different patient groups. The majority of the CD4+CD25+CD127− T cells showed demethylation of the TSDR within the Foxp3 locus, confirming its regulatory phenotype.
Conclusion. Our data show accumulation of Foxp3+ T cells within inflamed joints. These Foxp3+ T cells are mainly of stable T regulatory phenotype. The high frequency of Foxp3+ T cells in pSpA might contribute to the spontaneous resolution and remitting course of arthritis in pSpA as compared to the more persistent joint inflammation in RA.
- FOXP3
- TREG
- JOINT INFLAMMATION
- SPONDYLOARTHRITIS
The spondyloarthritides (SpA) comprise ankylosing spondylitis (AS), reactive arthritis (ReA), arthritis/spondylitis with inflammatory bowel disease, arthritis/spondylitis with psoriasis, and undifferentiated spondyloarthritis (uSpA). A high association with HLA-B27 is common among these diseases, which also share some clinical symptoms such as inflammatory back pain, asymmetrical involvement of large peripheral joints of the lower limbs, and the possible occurrence of sacroiliitis, spondylitis, enthesitis, and uveitis1. The SpA can be divided into SpA with predominant axial and SpA with predominant peripheral involvement. Both forms overlap in about 20%–30% of cases2. Axial inflammation in AS is often fluctuating with a change between appearance and disappearance of inflammation at specific sites. Also, peripheral joint inflammation in SpA is usually self-limiting, but can remit and migrate from joint to joint. This is different from rheumatoid arthritis (RA), where normally persistent inflammation in the same joint is observed3.
T regulatory cells (Treg) are essential for balancing the immune response against infection and for the establishment and maintenance of self-tolerance4. Initially, CD4+ Treg were characterized by high expression levels of CD25, a component of the high-affinity interleukin 2 (IL-2) receptor4. The identification of the forkhead box transcription factor FoxP3 as a master transcription factor of Treg provided a powerful tool for Treg analysis and identification5. The transcription factor Foxp3 is necessary for the development of Treg in the thymus, and is required for the maintenance of the suppressive function of mature peripheral Treg6. In mice, FoxP3 gene mutations and FoxP3 deletion results in lymphoproliferative autoimmune disorder due to the lack of Treg7. In humans, a disease called “immune dysregulation, polyendocrinopathy, enteropathy, X-linked” (IPEX) is caused by mutations in the FoxP3 gene and is associated with multiple inflammatory autoimmune organ involvement5. This suggests that immune and autoimmune responses are regulated by a sensitive balance between regulatory and effector T cells8.
To determine the potential effect of Treg in joint inflammation associated with SpA and RA, we analyzed the frequency and functional characteristics of FoxP3+ cells/CD4+ T cells in the peripheral blood (PB) and in the synovial fluid (SF) of patients with different forms of SpA and patients with RA.
MATERIALS AND METHODS
Patients
We obtained PB from 22 patients with AS, 19 patients with peripheral spondyloarthritis (pSpA; 8 patients with uSpA, 6 with reactive arthritis, 3 with psoriatic arthritis, and 2 with Crohn’s disease and arthritis), 15 patients with RA, and 12 healthy controls. We also analyzed SF mononuclear cells from 10 patients with AS, 20 with pSpA (8 patients with uSpA, 7 with reactive arthritis, 3 with psoriatic arthritis, and 2 with Crohn’s disease and arthritis), and 12 patients with RA. All patients with AS fulfilled the modified New York criteria, and all patients with pSpA fulfilled the European Spondylarthritis Study Group criteria. Patients’ characteristics are given in Table 1. More patients with RA than with AS or pSpA were on disease-modifying antirheumatic drug (DMARD) therapy. Matched samples of PB and SF could be analyzed from 8 patients with AS, 6 with RA, and 11 with pSpA.
Characteristics of patients giving samples of peripheral blood or synovial fluid.
All patients gave consent to the study, which was approved by the local ethics committee of the Charité Universitätsmedizin Berlin, Campus Benjamin Franklin.
Staining for T cell surface markers, FoxP3, intracellular cytokines, and analysis by flow cytometry
For staining of whole PB and SF, erythrocytes were lysed by incubation with Becton Dickinson Fluorescence Activated Cell Sorting (FACS) lysing solution (Becton Dickinson, Heidelberg, Germany). Then cells were washed with phosphate buffered saline/bovine serum albumin and stained for surface markers, i.e., CD4, CD25, or CD127. Foxp3 and cytokines were stained intracellulary after permeabilization using FACS™ Permeabilizing Solution (Becton Dickinson) as described9. To avoid nonspecific binding, staining was performed in the presence of Beriglobin (1 mg/ml; Centeon Pharma, Berlin, Germany). The following antibodies were used: anti-human CD4 (clone SK3; Becton Dickinson), anti-FoxP3 (clone eBio64DEC17; eBioscience, San Diego, CA, USA), anti-human CD127 antibody (clone hIL-7R-M21; Becton Dickinson), anti-CD25 antibody (2A3), anti-IL-2 (clone MQ1-17H12; Becton Dickinson), anti-IL-4 (clone Mp4-25D2; Becton Dickinson), anti-IL-10 (clone JES3.19F1), and anti-interferon-γ (IFN-γ; clone B27; Becton Dickinson). For flow cytometric analysis, a FACS Calibur (Becton Dickinson) and Cellquest software were used.
For detection of cytokines, PB or SF were stimulated for 6 h in the presence of anti-CD28 alone (1 μg/ml; clone B27.2; Becton Dickinson), or in the presence of anti-CD28 plus staphylococcus enterotoxin B (SEB; 1 μg/ml; Sigma-Aldrich, Deisenhofen, Germany). After 2 h of stimulation, brefeldin A (BFA, 10 μg/ml; Sigma Aldrich) was added to prevent cytokine release from cells.
FOXP3 methylation
To determine the methylation pattern within the Treg-specific demethylated region (TSDR) of the Foxp3 genomic locus, PB mononuclear cells (PBMC) or SF mononuclear cells were prepared by centrifugation over Ficoll-Paque™ Plus (GE Healthcare Bio-Science AB, Uppsala, Sweden). CD4+CD25+ T cells were enriched from SF or PBMC by magnetic absorbance cell separation (MACS−) sorting using the CD4+CD25+ Regulatory T cell isolation kit (Miltenyi, Bergisch Gladbach, Germany) according to the manufacturer’s instructions. After sorting, the percentage of CD4+CD25+Foxp3+ T cells was determined by flow cytometry. Some MACS-enriched samples were further stained with anti-CD4-PerCP, anti-CD127-PE, and CD25-APC, and Treg were further purified by sorting the CD4+CD25+CD127− T cell fraction by FACS. Genomic DNA of these enriched Treg populations was isolated using the QIAamp® DNA Blood Mini Kit (Qiagen, Hilden, Germany) and treated with bisulfite (EpiTect®, Qiagen). To quantify the fraction of methylated and nonmethylated FOXP3 TSDR, 60 ng bisulfite-treated DNA or the respective amount of plasmid standard was used for real-time polymerase chain reaction performed in a final reaction volume of 20 μl containing 10 μl FastStart Universal Probe Master (Roche Diagnostics, Mannheim, Germany), 50 ng/μl Lamda DNA (New England Biolabs, Frankfurt, Germany), 5 pmol/μl methylation or nonmethylation-specific probe, and 30 pmol/μl methylation or nonmethylation-specific primers as described10. The samples were analyzed in triplicates on an ABI 7500 Cycler (Applied Biosystems, Weiterstadt, Germany). The percentage of nonmethylated FOXP3 TSDR content was then calculated by dividing the nonmethylated copy number by the total genomic FOXP3 copy number. The percentage of TSDR-nonmethylated cells in the MACS-enriched population was adjusted to the percentage of CD4+CD25+CD127− cells determined by flow cytometry.
Statistics
For group analysis of data we used the Wilcoxon signed-rank test and, if paired samples were analyzed, the paired Wilcoxon signed-rank test and SPSS for Windows software.
RESULTS
Increased frequency of synovial FoxP3+ cells/CD4+ T cells in SpA vs RA
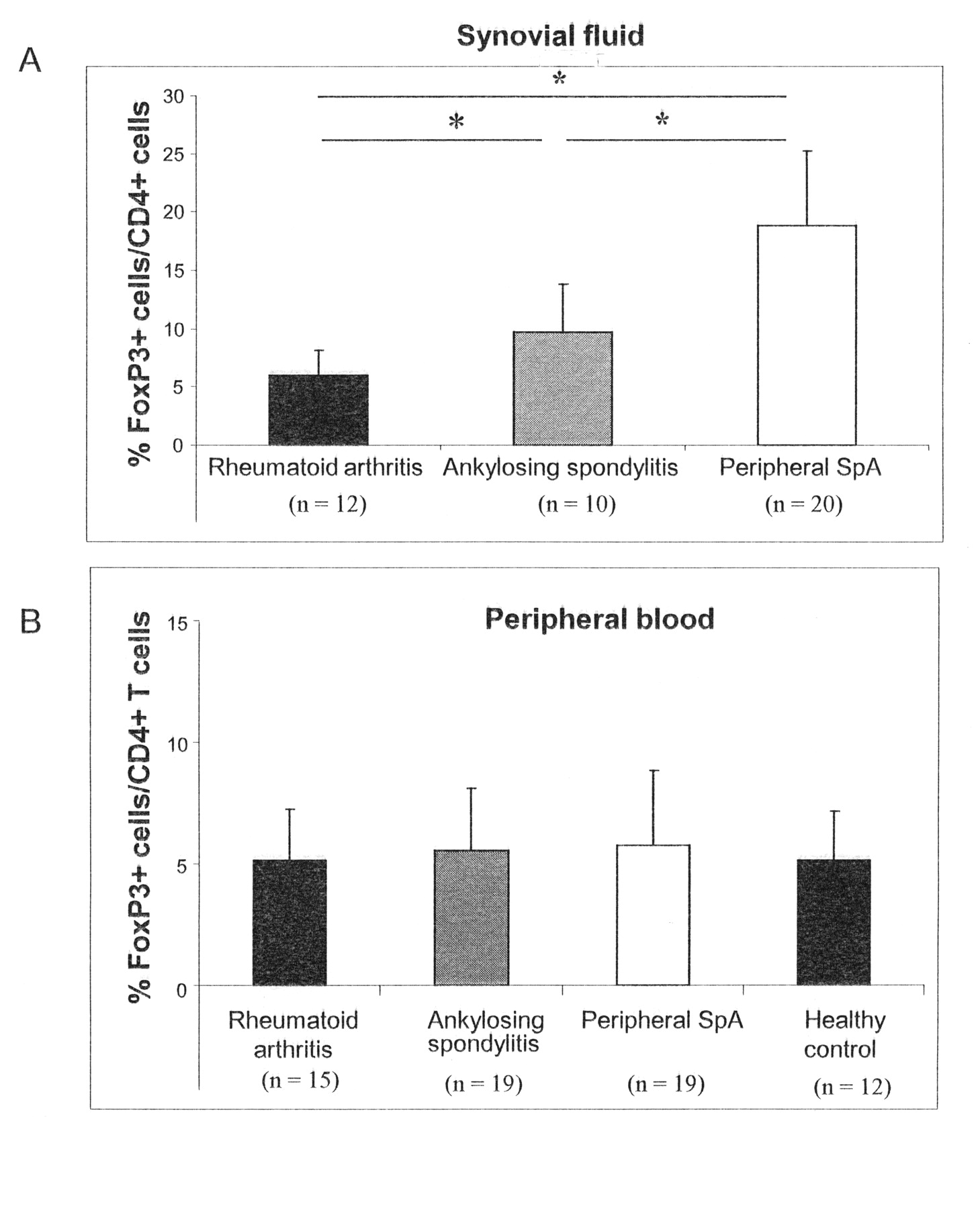
The percentage of Foxp3+ cells among CD4+ T cells was determined in the PB and SF of patients with RA, AS, and pSpA. There was a clear difference among the diseases in the frequency of synovial CD4+FoxP3+ T cells. The highest frequency of FoxP3+ cells/CD4+ cells was seen in patients with pSpA. In this disease the frequency of synovial FoxP3+ cells/CD4+ T cells (18.79% ± 6.41%) was significantly higher than in AS (9.69% ± 4.11%; p = 0.005) and in RA (5.95% ± 2.21%; p = 0.002; Figure 1A). Among patients with pSpA, no difference in the frequency of FoxP3+ cells/CD4+ T cells was found between patients with axial involvement and patients without axial involvement (data not shown). Comparing AS and RA, significantly higher frequencies of synovial FoxP3+ cells/CD4+ T cells were observed in patients with AS (p = 0.007).

Increased frequencies of Foxp3+cells/CD4+ T cells in peripheral arthritis and ankylosing spondylitis (AS) compared to rheumatoid arthritis (RA) in synovial fluid. The frequency of Foxp3+cells/CD4+ T cells (mean ± SD) was determined cytofluorometrically in synovial fluid (A) and peripheral blood (B) from patients with RA, AS, peripheral spondyloarthritis, and healthy individuals. *p < 0.05; Wilcoxon signed-rank test.
In contrast to synovial Foxp3+ T cells, no significant differences were found in the frequency of PB FoxP3+ cells/CD4+ T cells in patients with RA, AS, and pSpA (all p > 0.05). Moreover, none of the different groups of patients showed a significant difference of blood FoxP3+ cells/CD4+ T cells compared to healthy controls (all p > 0.05; Figure 1B).
Increased frequencies of FoxP3+ cells/CD4+ T cells in synovial fluid compared to peripheral blood
The previous data suggested that Foxp3+ T cells are enriched within the SF compared to blood. To study this in more detail we analyzed matched PB and SF samples from 6 patients with RA, 8 with AS, and 11 with pSpA. As shown in Figure 2, the frequency of Foxp3+ T cells/CD4+ T cells was higher in SF compared to PB in most of the patients, no matter whether they had RA, AS, or pSpA. In line with the previous data, we observed the most prominent accumulation of Foxp3+ T cells/CD4+ T cells in the SF of patients with pSpA.

Increased frequency of FoxP3+cells/CD4+ T cells in synovial fluid (SF) compared to peripheral blood (PB) in patients with rheumatoid arthritis (RA), ankylosing spondylitis (AS), and peripheral spondyloarthritis (pSpA). The frequency of Foxp3+cells/CD4+ T cells was determined within blood and synovial fluid (matched samples) of patients with RA, AS, and pSpA. *p < 0.05, **p < 0.01; paired Wilcoxon signed-rank test.
By analyzing data of all patients with simultaneous measurement of C-reactive protein (CRP) as a marker for disease activity, a correlation of the percentage of PB FoxP3+ cells/CD4+ T cells and CRP was found in patients with pSpA (Spearman r = 0.7608; p = 0.0065), but not in patients with AS (Spearman r = 0.3176; p = 0.2306), nor in patients with RA (Spearman r = 0.2308; p = 0.4273). Synovial fluid FoxP3+ cells/CD4+ T cells did not correlate to CRP in all groups of patients, although there was a trend in patients with pSpA (pSpA: Spearman r = 0.4750, p = 0.0736; AS: Spearman r = 0.2857, p = 0.4927; RA: Spearman r = 0.1538, p = 0.6331).
CD4+FoxP3+ T cells do not secrete effector cytokines
In addition to its presence on regulatory T cells, Foxp3 is temporarily expressed on activated T cells. To demonstrate that the T cells detected by Foxp3 staining within the blood and in particular in the inflamed tissue are indeed regulatory T cells, we analyzed whether synovial CD4+FoxP3+ T cells secrete effector cytokines spontaneously or after stimulation with SEB.
As shown in Figure 3, CD4+FoxP3+ T cells did not secrete effector cytokines such as IL-2, IFN-γ, tumor necrosis factor-α (TNF-α), or IL-10 either spontaneously or after SEB stimulation in any of the different patient groups (Figure 3). In contrast, cytokine production was readily detectable among CD4+Foxp3− T cells.
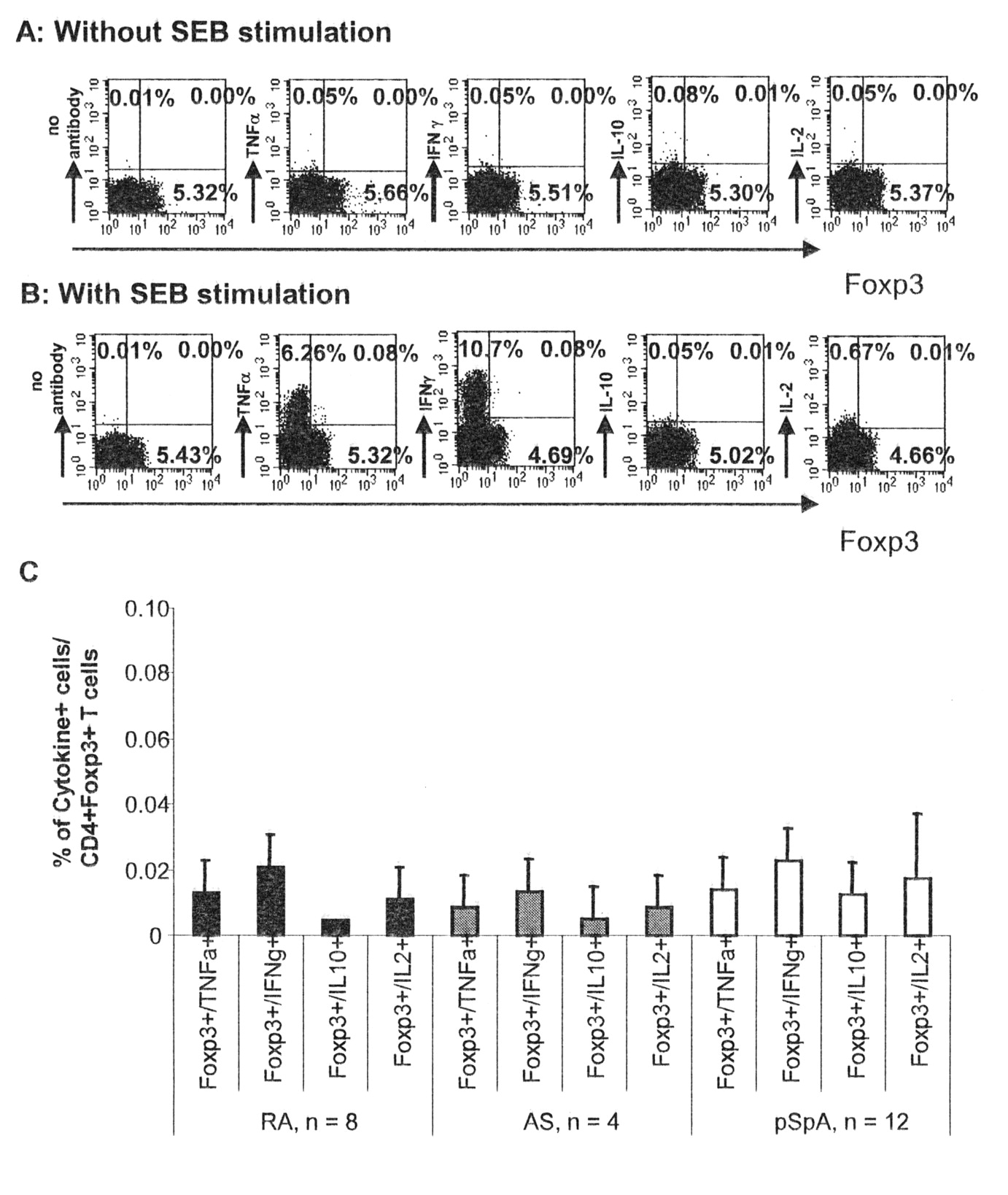
Synovial fluid CD4+FoxP3+ T cells do not secrete tumor necrosis factor-α, interferon-γ, interleukin 2 (IL-2), or IL-10. Synovial fluid cells were stimulated in the absence or presence of staphylococcus enterotoxin B (SEB), and cytokine production and Foxp3 expression among gated CD4+ T cells was determined by intracellular staining. An example of cytokine secretion in relation to Foxp3 expression of CD4+ T cells without (A) and with (B) stimulation is shown. Numbers indicate the frequency of cells/quadrant. A summary of all analysis in patients with rheumatoid arthritis (RA), ankylosing spondylitis (AS), and peripheral spondyloarthritis (pSpA) is shown in C.
The majority of peripheral CD4+FoxP3+ T cells coexpress CD25 and lack CD127 expression
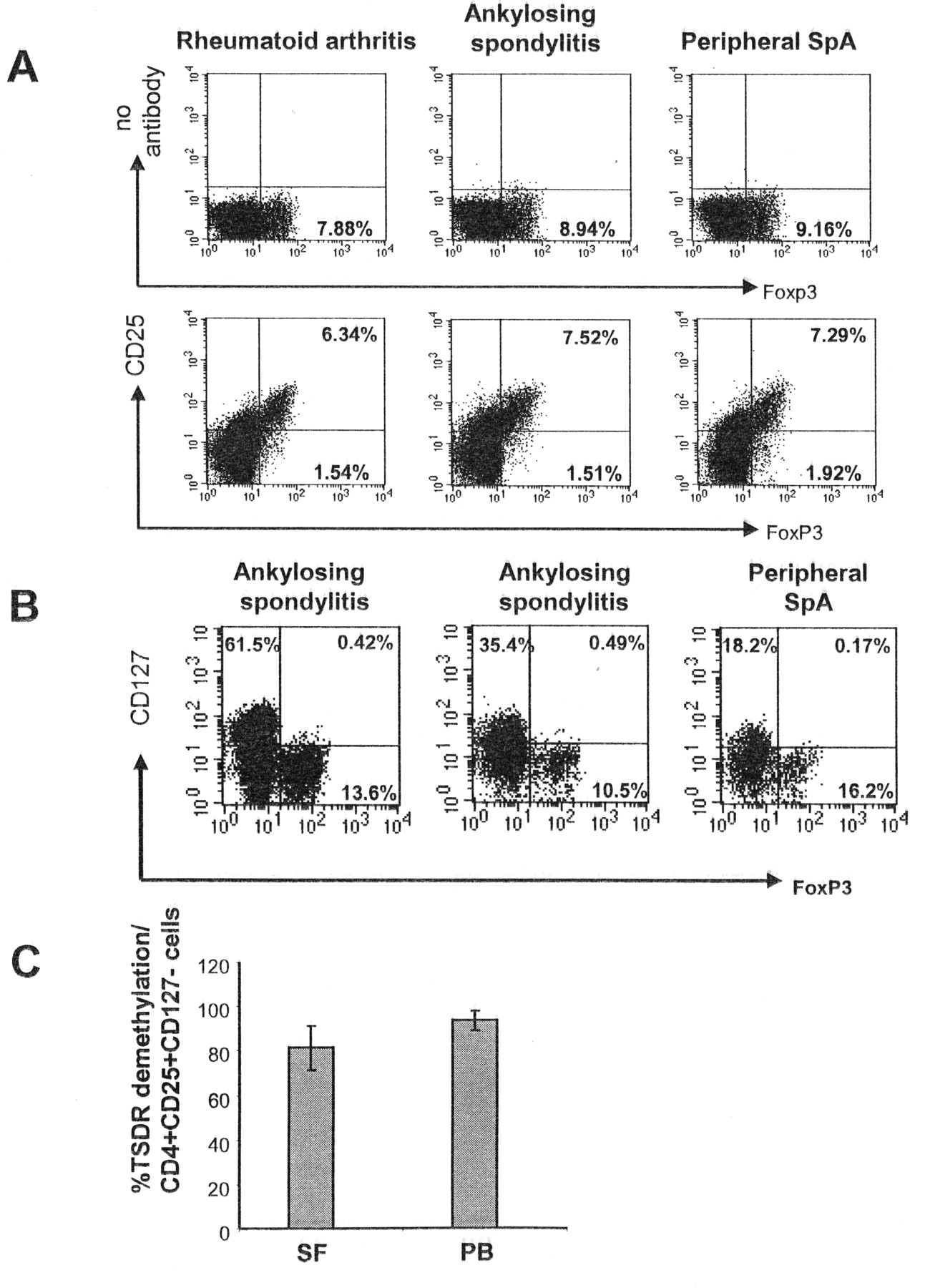
To further characterize the CD4+Foxp3+ T cell subset, we looked for CD25 coexpression on Foxp3+ T cells. In 10 patients, > 75% of the FoxP3+CD4+ T cells within the peripheral blood also expressed CD25 (Figure 4A). In 2 patients with AS and 1 patient with pSpA we further analyzed expression of CD127. CD127, representing the IL-7 receptor, is expressed on the majority of naive T cells and on effector T cells but not on Treg, and can be used to discriminate activated T cells from Treg11,12. In all 3 patients, > 95% of Foxp3+ T cells within the inflamed joint were CD127-negative (Figure 4B). To get further evidence for the Treg origin of the Foxp3+ T cells found in the SF or PB of patients with SpA, we determined the DNA methylation pattern within the TSDR of the Foxp3 locus. Demethylation within this region corresponds to a stable Treg phenotype10,13. Therefore, we determined the percentage of TSRD demethylation in CD4+CD25+ MACS-enriched or CD4+CD25+CD127− FACS-sorted T cells in the blood of 2 patients with AS and in SF of 4 patients with AS and adjusted the content of nonmethylated TSDR to the percentage of CD4+CD25+CD127− T cells as putative Treg in the sample. As shown in Figure 4C, more than 80% of the CD4+CD25+CD127− T cells of SF samples and more than 90% of the CD4+CD25+CD127− T cells in the blood of patients with AS showed demethylation within the TSDR, indicating that the majority of CD4+CD25+CD127− T cells, from both blood and inflamed tissue of patients with SpA, indeed represent Treg.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
The majority of CD4+Foxp3+ T cells exhibits characteristics of regulatory T cells. The frequency of CD25+ cells among CD4+Foxp3+ T cells was determined in peripheral blood T cells of patients with rheumatoid arthritis, ankylosing spondylitis (AS), and peripheral spondyloarthritis (pSpA; A). The control staining is shown in the upper panel; the positive staining is shown in the lower panel. Numbers indicate the percentage of cells/quadrant on gated CD4+ T cells. Synovial T cells of 2 patients with AS and 1 patient with pSpA were stained for CD127 in relation to Foxp3 expression (B). Numbers indicate the percentage of cells/quadrant in gated CD4+ T cells. C. Demethylation of the Treg-specific demethylated region (TSDR) within the Foxp3 locus was determined for CD4+CD25+CD127− T cells isolated from peripheral blood (PB; n = 2) or synovial fluid (SF; n = 4) of patients with AS.
DISCUSSION
There is increasing evidence that Treg compartmentalization and trafficking may be specific for distinct sites of inflammation, resulting in a specific retention of Treg at inflamed sites that require immune regulation14. Therefore, we determined the distribution of Foxp3+ T cells in PB and in the inflamed joints of patients with pSpA, AS, or RA. In PB we did not observe enhanced Treg numbers in any of the diseases compared to healthy controls. In contrast, we found higher frequencies of Foxp3+ cells/CD4+ T cells in the inflamed joint fluid compared to PB in patients with RA, AS, and pSpA, suggesting that regulatory T cells accumulate within the inflamed joints.
Contradictory results about the frequency of peripheral and SF Treg are reported in different inflammatory joint diseases, but especially in RA. In RA the frequency of CD4+CD25high+ Treg was reported to be higher in the SF than in the PB using matched samples15,16, similar to our findings, while others reported no difference in the frequency of Treg in both compartments17 in unmatched patients with RA. Statistical analysis of our unmatched samples from patients with RA also did not show significant differences (data not shown), suggesting that the comparison of matched samples is superior in identifying small differences.
As Foxp3 is also transiently expressed on activated T cells, we gained further evidence for the “true” Treg origin of the cells identified by Foxp3 staining in patients with inflammatory joint disease by further characterizing these cells. We showed that these cells coexpressed the high-affinity IL-2 receptor CD25 but lacked expression of CD127, the IL-7 receptor that is considered a specific feature of Treg11,12. Further, it is well documented that Treg cells fail to proliferate or produce effector cytokines such as IFN-γ or TNF-α, nor do they produce IL-2 in response to T cell receptor stimulation18. We proved that the CD4+Foxp3+ T cells, neither those from blood nor from SF, produce cytokines such as IL-2, IFN-γ, or TNF-α. Also at the genomic level, i.e., by analyzing the methylation pattern of the TSDR within the Foxp3 locus, we were able to show that most of the CD4+CD25+CD127− T cells exhibit demethylated TSDR, which was found to correlate with stable Foxp3 expression and Treg function10,13. However, the slightly lower percentage of TSDR demethylation among T cells within the SF compared to the blood might indeed reflect a higher fraction of activated T cells transiently expressing Foxp3 and CD25 in the inflamed synovial compartment.
Several experimental mouse models suggest that Foxp3+ T cells actively suppress inflammation in the periphery19,20. Considering that joint inflammation in pSpA remits and migrates from joint to joint, whereas in patients with RA it is rather persistent in individual joints, Foxp3+ T cells within the SF might indeed contribute to the control and resolution of local joint inflammation. This is also supported by the correlation between systemic (and to some extent synovial) frequencies of FoxP3+ T cells/CD4+ T cells and CRP, as a marker of disease activity, in patients with pSpA. In line with our data, in patients with juvenile idiopathic arthritis a correlation between the percentage of synovial Treg and the clinical course has also been suggested. Patients with a milder disease showed a higher frequency of CD4+CD25high+ T cells than patients with a more severe disease21. In reverse, the lower numbers of Foxp3+ T cells in RA and the lack of correlation to CRP might reflect an insufficiency to control inflammation that, according to elevated CRP and ESR levels, persists even though the patients are treated with DMARD.
In RA, inflammation within the peripheral joints can lead to local erosions, i.e., bone destruction, and it is suggested that Treg are involved in the control of this bone destruction22. Peripheral arthritis in AS and pSpA is usually nonerosive. It is therefore tempting to speculate that the higher frequency of Treg in pSpA and AS might contribute to the nonerosive expression of arthritis. However, the gradual difference in the percentage of synovial FoxP3+ cells/CD4+ T cells between pSpA and AS and the rather small difference in synovial Treg numbers between RA and AS suggest that other disease-specific mechanisms are superior to Treg in the control of local bone destruction.
We have shown that Treg accumulate within the inflamed joints of patients with RA, AS, and pSpA, with the highest frequencies seen in pSpA. Because of the high percentage of TSDR demethylation and phenotypic characteristics, as well as the lack of cytokine secretion of Foxp3+ T cells within the joints, our data suggest that most of the Foxp3+ T cells are Treg. The numeric differences in synovial Foxp3+ T cells among various forms of joint inflammation point to differences in the immunological mechanisms regulating joint inflammation.
Footnotes
-
Supported by grants from the Deutsche Forschungsgemeinschaft (DFG): Ap82/3-1, SFB 633, Project A4.
- Accepted for publication June 28, 2011.
REFERENCES
- 1.
- 2.
- 3.
- 4.
- 5.
- 6.
- 7.
- 8.
- 9.
- 10.
- 11.
- 12.
- 13.
- 14.
- 15.
- 16.
- 17.
- 18.
- 19.
- 20.
- 21.
- 22.