

Abstract
Objective. To investigate whether the expression of cytokine, nociception-associated ion channel, and axon guidance genes in patients with skin cell fibromyalgia syndrome (FMS) differs from healthy controls, potentially contributing to pain and small-fiber degeneration in FMS.
Methods. We prospectively recruited 128 patients and 26 healthy controls. All study participants underwent neurological examination, and a skin punch biopsy was obtained from the lateral calf and thigh. Skin samples were processed to histologically determine intraepidermal nerve fiber density (IENFD) and for primary fibroblast and keratinocyte cell cultures. Gene expression of selected pro- and antiinflammatory cytokines, nociception-associated ion channels, and axon guidance cues was assessed with quantitative real-time PCR.
Results. In fibroblasts, transforming growth factor–ß1 (TGF-ß1) gene expression was higher in patients with FMS compared to controls (calf and thigh: p < 0.001). Also, expression was higher in patients than in controls for these variables: hyperpolarization-activated cyclic nucleotide-gated ion channel 2 (calf: p < 0.01), ephrin-A4 (EFNA4; calf: p < 0.05, thigh: p < 0.001), and ephrin receptor-A4 (EPHA4; thigh: p < 0.05). In keratinocytes, interleukin 10 gene expression was higher in patients with FMS than in controls (thigh: p < 0.05). While no intergroup difference was found for nociception-associated ion channels, EFNA4 and EPHA4 (calf: p < 0.01 each) expression was higher in patients with FMS than in controls. Axon guide expression did not correlate with IENFD.
Conclusion. In FMS, skin cells may contribute to cutaneous nociception by differentially expressing membrane-bound and soluble pain mediators and axon pathfinders.
Small nerve fiber pathology is present in a subgroup of patients with fibromyalgia syndrome (FMS)1; however, the underlying mechanism and the effect on FMS pain remains unclear. Given the close vicinity of keratinocytes, fibroblasts, and axon terminals, a vital effect of skin cells on nociceptor structure and function is assumed. There is also increasing evidence for a pathophysiological role of keratinocytes and fibroblasts in sensory transduction and nociceptor sensitization2,3. Mechanisms may involve pro- and antiinflammatory cytokines4, neurotrophic factors5, and axon guidance cues6,7. Keratinocytes and fibroblasts are highly active in secreting algesic mediators4,8 and express many nociception-associated ion channels9,10. Further, skin cells are involved in axon pathfinding5,11.
A better understanding of cutaneous cytokine, ion channel, and axon guide expression may unravel the potential interplay of skin cells and nociceptors in FMS pain. While no data are available on the expression of nociception-associated ion channels and axon guidance cues in FMS skin, some studies showed higher proinflammatory cytokine expression in FMS skin samples12, while others did not13. Higher skin expression of the microRNA let-7d in patients with FMS potentially contributes to reduced intraepidermal nerve fiber density through downregulation of the proregenerative insulin-like growth factor-1 receptor14.
The major drawback of these studies is that whole skin preparations were used, which does not allow determination of the cellular source of the detected mediators and may disguise subtle differences in target expression. We therefore set out to investigate relative gene expression of selected pro- and antiinflammatory cytokines, nociception-associated ion channels, and axon guidance cues in individual fibroblast and keratinocyte primary cell cultures obtained from patients with FMS and healthy controls to further elucidate the role of skin cells in nociception and nerve fiber growth.
MATERIALS AND METHODS
Patients and controls
From 2014 to 2017 we prospectively recruited 128 patients with FMS (118 women, 10 men; median age: 51 yrs; range: 22–75 yrs) from all over Germany. FMS was diagnosed according to current criteria [American College of Rheumatology (ACR) criteria 1990 and 2010; German S3 guidelines for FMS]15,16,17. Extensive clinical data of our patients have been summarized elsewhere17a. All patients were first interviewed using a structured survey by telephone and were then personally seen at our department (by DE and NÜ). Patients underwent a detailed medical interview, complete neurological examination, and nerve conduction studies (tibial and sural nerve) to exclude polyneuropathy. Additionally, they filled in the Fibromyalgia Impact questionnaire (FIQ)18. Diagnostic criteria were individually checked. We additionally recruited a group of 26 healthy controls (22 women, 4 men; median age: 50 yrs; range: 23–66 yrs) from among friends and acquaintances of the patients. Our study was approved by the Würzburg Medical School Ethics Committee (#121/14) and written informed consent was obtained from all study participants.
Skin punch biopsies
Six-millimeter skin punch biopsies (Stiefel) were obtained from the lateral calf and thigh. One half of each specimen was used to determine intraepidermal nerve fiber density (IENFD) as described earlier19. The other half was processed for primary fibroblast and keratinocyte cell cultures.
Primary skin cell culture
Epidermis and dermis were mechanically separated using a scalpel and then cut into 12 pieces. Fibroblasts were cultured in Dulbecco modified Eagle’s medium F-12 (DMEM/F-12, Life Technologies), supplemented with penicillin/streptomycin (1%) and fetal calf serum (10%). Keratinocytes grew in EpiLife medium (Thermo Fisher Scientific), which was supplemented with 10,000 U/ml penicillin/streptomycin and 1× EpiLife Defined Growth Supplement (Life Technologies). Fibroblasts and keratinocytes were cultivated at 5% CO2 and 37°C. TrypLE Express (Life Technologies) was used for trypsinization of fibroblasts (1 min at 37°C, CO2 5%). Keratinocyes were enzymatically digested using StemPro Accutase (Life Technologies) for up to 20 min at 37°C, 5% CO2. Cells were stored in liquid nitrogen before further processing.
RNA extraction
After 2 passages, RNA was extracted from keratinocytes and fibroblasts using the miRNeasy Mini Kit (Quiagen). Following suspension in 700 µl QIAzol lysis reagent, samples were incubated for 5 min at room temperature. Upon adding 140 µl chloroform (Carl Roth GmbH & Co.), samples were mixed by vigorous shaking and were incubated for up to 3 min at room temperature. After centrifugation, 525 µl ethanol 100% was added to the upper phase (12,000 g, 15 min, +4°C) and samples were recentrifuged at room temperature (8000 g, 20 s). Total RNA was eluted in 30 µl RNAse-free water. RNA concentration was determined using a Nanodrop spectrophotometer (Peqlab) and samples were stored at −80°C before further processing.
Reverse transcription PCR
Reagents and cyclers were purchased from Applied Biosystems. Using TaqMan Reverse Transcription Reagents, 500 ng mRNA was reverse transcribed. The reactions contained 10 µl 10× PCR buffer, 6.25 µl multiscribe reverse transcriptase, 2 µl RNase inhibitor, 22 µl MgCl2, and 20 µl deoxyribonucleoside triphosphate. PCR analyses were run in an ABI PRISM 7700 Cycler under the following conditions: annealing (25°C, 10 min), reverse transcription (48°C, 60 min), and enzyme inactivation (95°C, 5 min).
Gene expression analyses
Five µl cDNA were used for quantitative real-time PCR (qRT-PCR) analysis with TaqMan Universal Master Mix and a StepOne Plus cycler. We investigated proinflammatory cytokines [interleukin (IL-)1ß (Hs00174097_m1)], IL-6 (Hs00174131_m1), IL-8 (Hs00174103_m1), tumor necrosis factor-α [(TNF-α), Hs00174128_m1], antiinflammatory cytokines (IL-4, Hs00174122_m1), IL-10 (Hs00174086_m1), transforming growth factor-ß1 [(TGF-ß1), Hs99999918_m1], nociception-associated ion channels [transient receptor potential vanilloid-1 (TRPV1), Hs00218912_m1], voltage-gated sodium channel 1.7 (Nav1.7, Hs00161567_m1), hyperpolarization-activated cyclic nucleotide-gated ion channel-2 (HCN2, Hs00606903_m1), and 2 pairs of axon guidance molecules with their main receptor [ephrin-A4 (EFNA4), Hs00193299_m1; ephrin-receptor-A4 (EPHA4), Hs00953178_m1; and netrin1 (NTN1), Hs00924151_m1], uncoordinated-5 homolog B (Unc5b; Hs00900710_m1). Serving as an endogenous control was 18sRNA (Hs99999901_s1). Each reaction contained 12.5 µl TaqMan Master Mix and 1.25 µl primer. The cycler conditions were 50°C, 2 min; 95°C, 10 min; 45 cycles with 95°C, 15 s; 60°C, 1 min. PCR plates contained a negative control and a calibrator sample, which was the sample of the control subject whose threshold cycles (Ct) values were next to the calculated mean of all control samples individually measured for each primer. We applied the 2-∆∆Ct method for relative gene expression analysis20.
Statistical analysis
We used SPSS 25 (IBM Corp.) for statistical analysis and calculated the Mann-Whitney U test for intergroup comparisons of the non-normally distributed qRT-PCR data. IENFD data were used as continuous variables. Results are presented as scatter plots with integrated box plots created with GraphPad Prism 7. Median values are marked with a black horizontal line, the 25th and 75th percentile are determined by whiskers. P < 0.05 was considered significant.
RESULTS
Basic clinical characteristics
Neurological examination and nerve conduction studies were normal in all cases. There were 117/128 patients (91%) who fulfilled the ACR 1990 criteria, and all patients fulfilled the ACR 2010 criteria and the German S3 guidelines for FMS. Patients reported a median current pain intensity of 6/10 (0–9) on a numerical rating scale (NRS) and a maximum pain intensity of 8/10 (5–10). The NRS ranged from zero to 10 (no pain–worst pain). Median pain duration was 11 years (0.75–56 yrs). Median FIQ sum score reached 48 points (18–71). A total of 102/128 patients (80%) were taking analgesic medication: 61 took one, 34 took two, and 7 took three drugs. Regarding comorbidities, 92/128 patients (72%) reported none, while the top 3 comorbidities requiring medical treatment were hypothyroidism (31), arterial hypertension (22), and depressive symptoms (15).
Higher expression of TGF-ß1 in skin fibroblasts of patients with FMS compared to controls
P values of gene expression comparisons between skin cells of patients with FMS and healthy controls are summarized in Appendix 1. Gene expression of the investigated proinflammatory cytokines did not differ between groups in skin fibroblasts obtained from the calf and the thigh (Figure 1A, 1B). Among antiinflammatory cytokines, TGF-ß1 gene expression was higher in fibroblasts from both the calf and the thigh in patients with FMS compared to healthy controls (p < 0.001 each, Figure 1C, 1D).
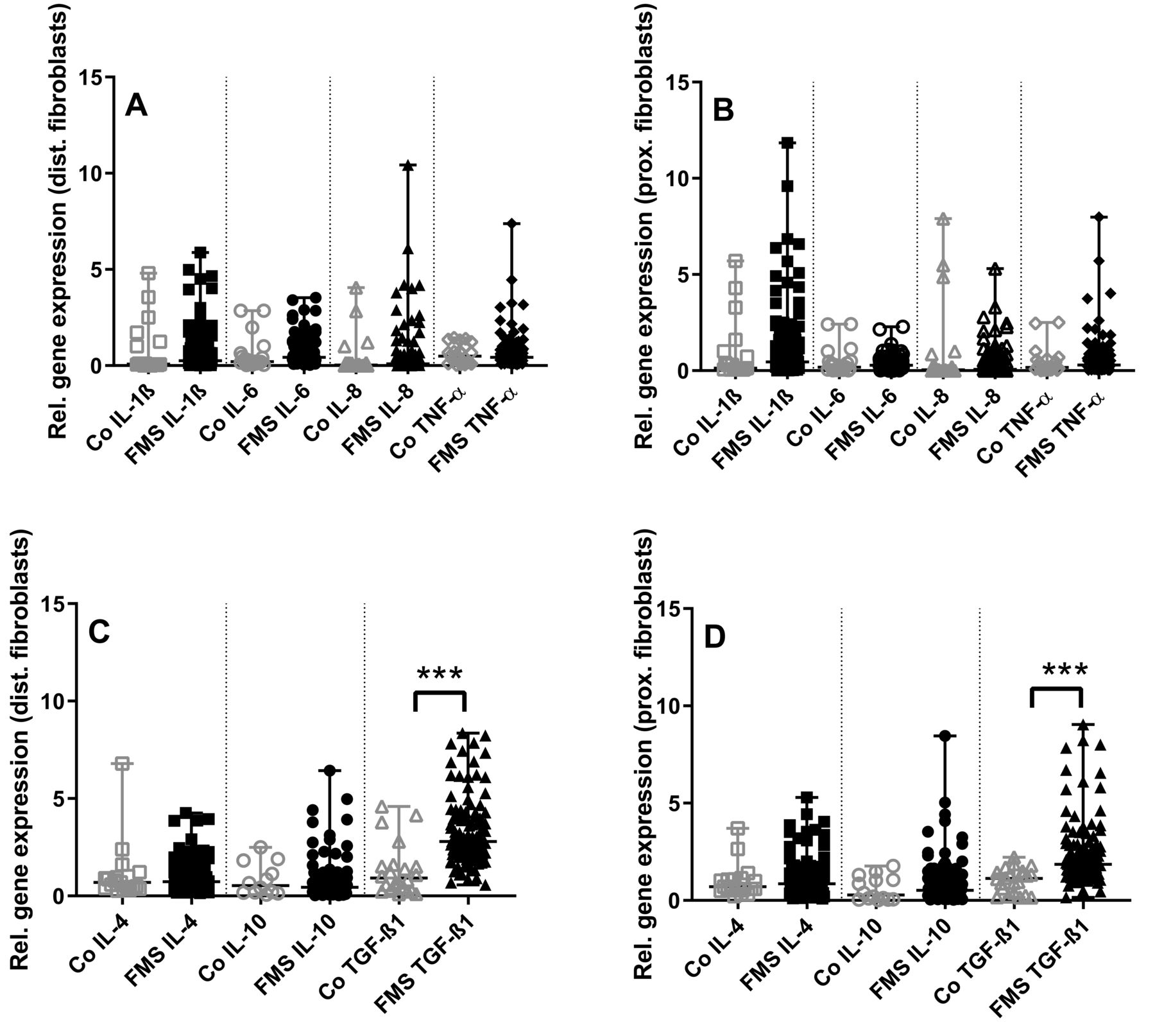
Gene expression panel of pro- and antiinflammatory cytokines in skin fibroblasts of patients with fibromyalgia syndrome (FMS) compared to healthy controls (Co). The scatter plots illustrate gene expression of the proinflammatory cytokines interleukin (IL-) 1β, IL-6, IL-8, tumor necrosis factor-α (TNF-α) (panels A, B), and the antiinflammatory cytokines IL-4, IL-10, transforming growth factor-β1 (TGF-ß1; panels C, D). Data were obtained from fibroblasts derived from skin punch biopsies of the calf (A, C) and the thigh (B, D). Proinflammatory cytokine expression did not differ between groups (A, B). Among the antiinflammatory cytokines, TGF-ß1 expression was higher in fibroblasts from the calf and the thigh compared to controls (C, D). *** p < 0.001.
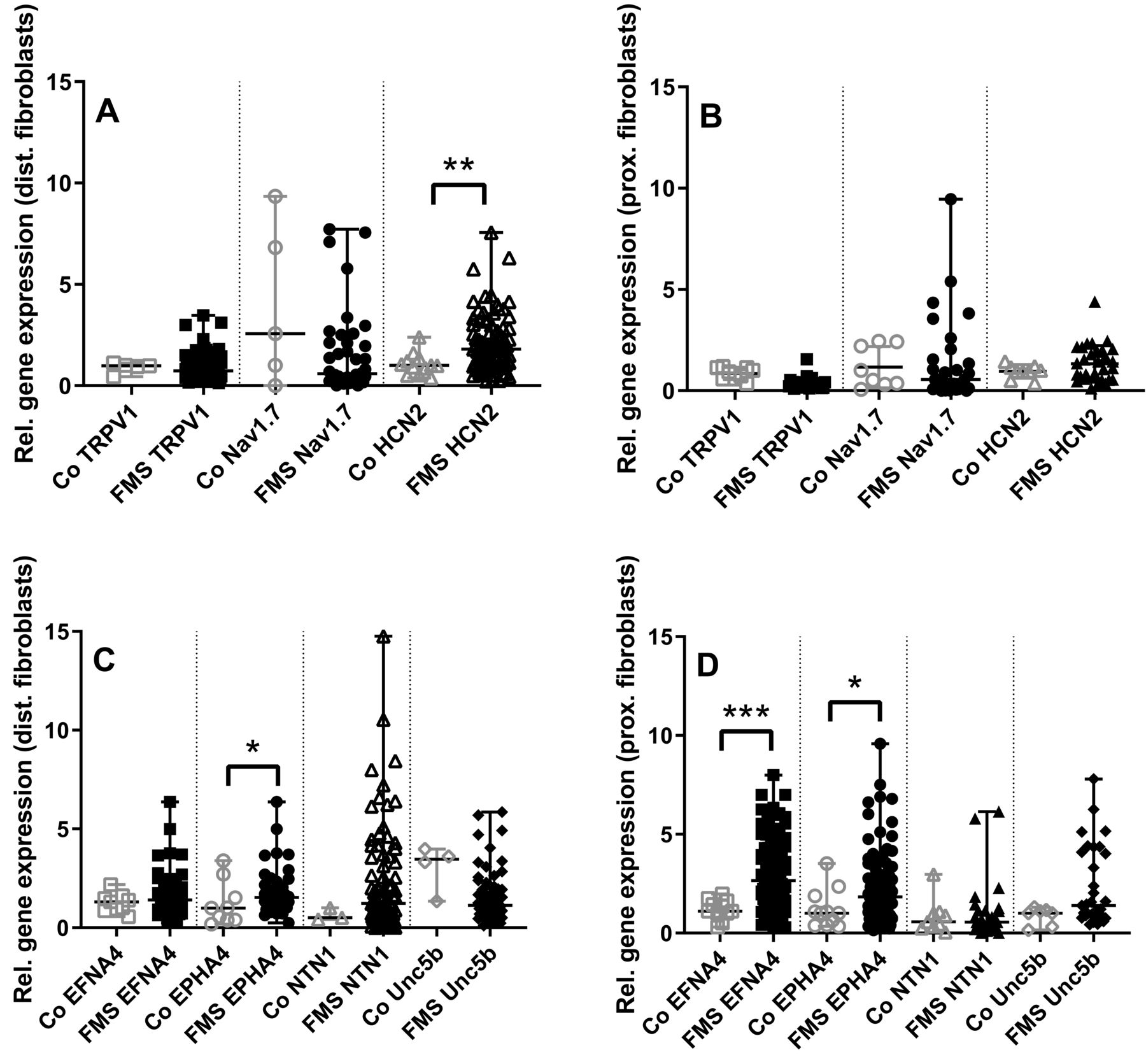
Higher expression of HCN2, EFNA4, and EPHA4 in fibroblasts from calf skin of patients with FMS compared to controls
Gene expression of nociception-associated ion channels was similar in fibroblasts of patients with FMS and healthy controls except for a higher expression of HCN2 in fibroblasts obtained from the calf of patients with FMS (p < 0.01, Figure 2A, 2B).
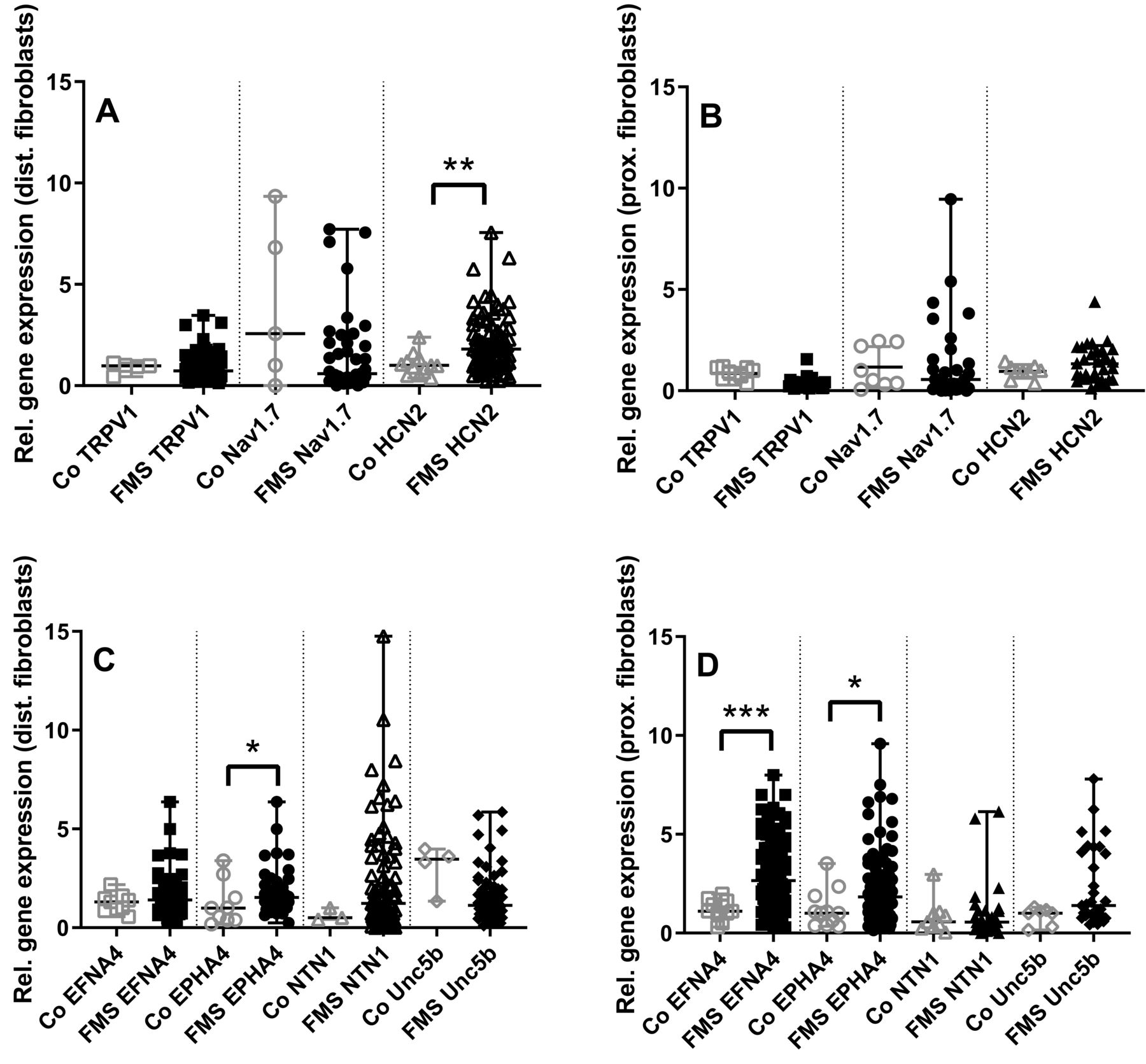
Gene expression panel of ion channels and axon guides in skin fibroblasts of patients with fibromyalgia syndrome (FMS) compared to healthy controls (Co). The scatter plots illustrate gene expression of the pain-associated ion channels transient receptor potential vanilloid-1 (TRPV1), voltage-gated sodium channel (Nav) 1.7, hyperpolarization-activated cyclic nucleotide-gated ion channel-2 (HCN2; panels A, B), and the axon guidance cues ephrin-A4 (EFNA4), ephrin receptor-A4 (EPHA4), netrin1 (NTN1), and uncoordinated-5 homolog B (Unc5b; panels C, D). Data were obtained from fibroblasts derived from skin punch biopsies of the calf (A, C) and the thigh (B, D). HCN2 expression was higher in fibroblasts obtained from the calf of patients with FMS compared to controls (A, B), while no intergroup difference was found for the other pain-associated ion channels investigated. EFNA4 and EPHA4 expression was higher in fibroblasts obtained from calf and thigh of patients with FMS compared to controls (C, D). RNA expression was not detectable or gave identical results for some targets in the group of healthy controls, hence plotted data points do not add up to n = 26. * p < 0.05, ** p < 0.01, *** p < 0.001.
When investigating the expression of axon guidance cues in skin fibroblasts, we found higher gene expression of EFNA4 in skin fibroblasts from the calf of patients with FMS (p < 0.05) and also higher expression of EFNA4 (p < 0.001) and EPHA4 (p < 0.05) in skin fibroblasts from the thigh of patients with FMS compared to controls (Figure 2C, 2D).
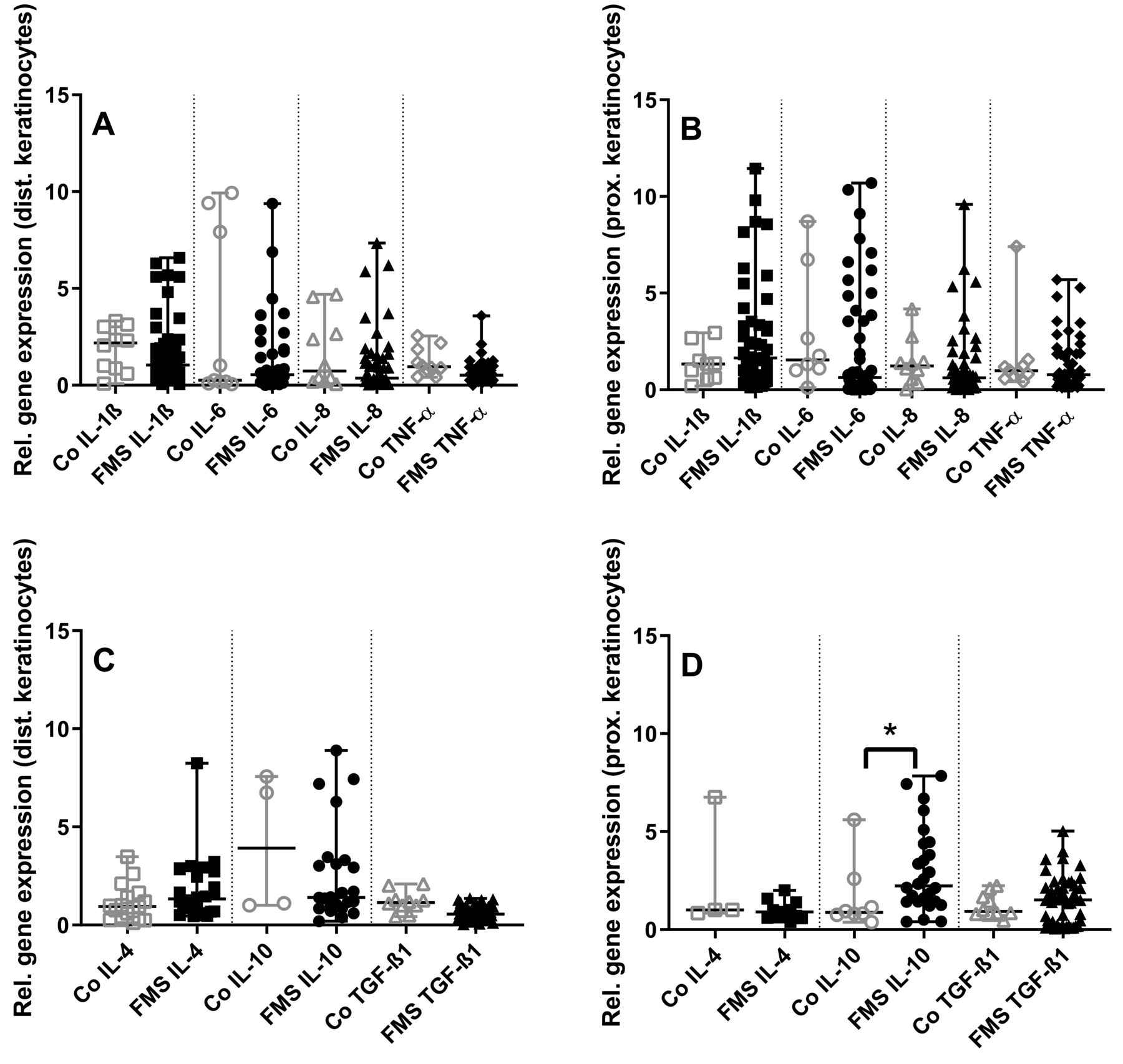
Higher expression of IL-10 in skin keratinocytes from the thigh of patients with FMS compared to controls
Assessment of keratinocytes revealed no intergroup differences for proinflammatory cytokines (Figure 3A, 3B). Among the investigated antiinflammatory cytokines, IL-10 gene expression was higher in keratinocytes from the thigh of patients with FMS than in controls (p < 0.05, Figure 3C, 3D).
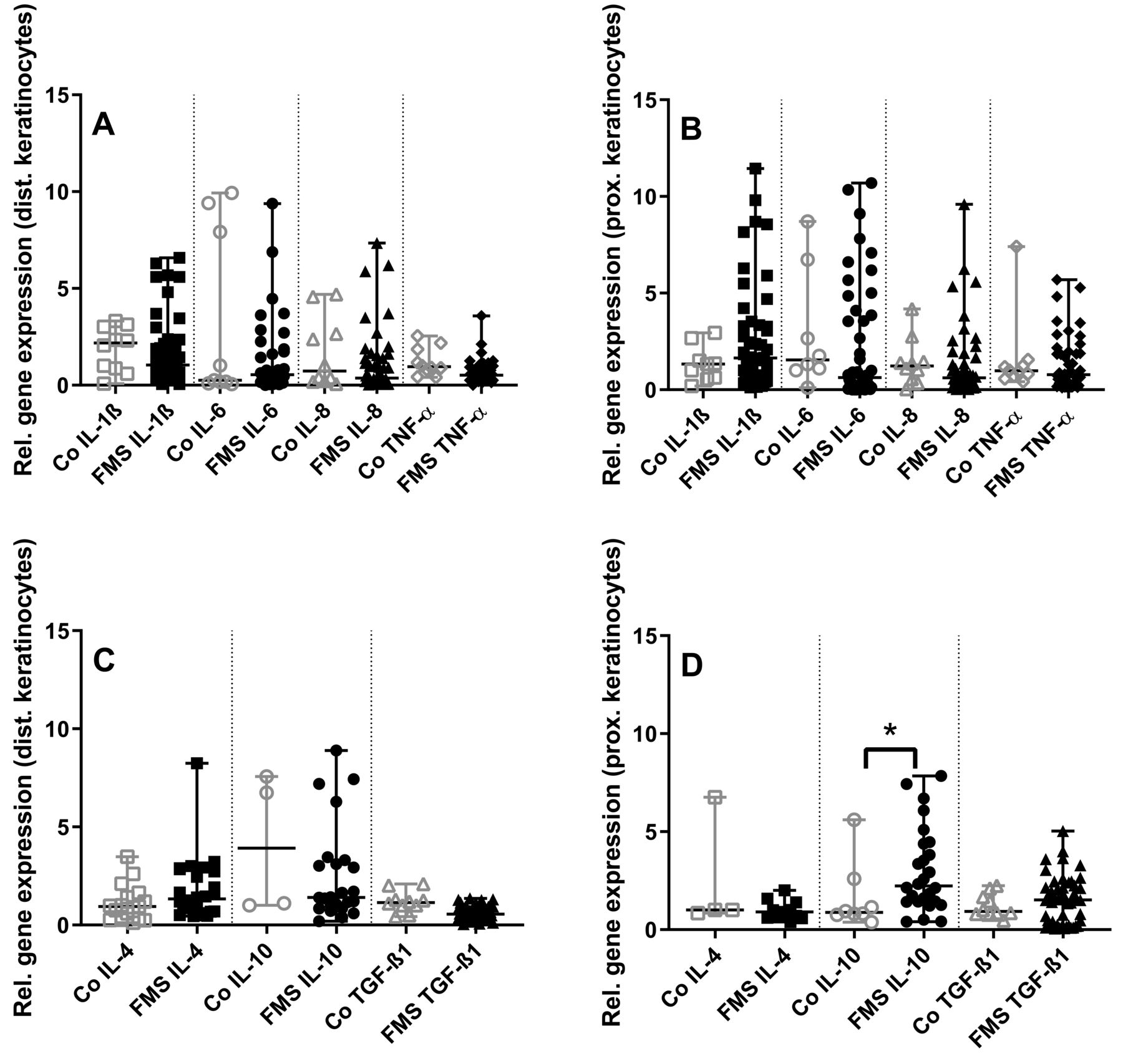
Gene expression panel of pro- and antiinflammatory cytokines in skin keratinocytes of patients with fibromyalgia syndrome (FMS) compared to healthy controls (Co). The scatter plots illustrate gene expression of the proinflammatory cytokines interleukin (IL-) 1ß, IL-6, IL-8, tumor necrosis factor-α (TNF-α; panels A, B), and the antiinflammatory cytokines IL-4, IL-10, transforming growth factor-β1 (TGF-ß1; panels C, D). Data were obtained from keratinocytes derived from skin punch biopsies of the calf (A, C) and the thigh (B, D). Proinflammatory cytokine expression did not differ between groups (A, B). Among the antiinflammatory cytokines, IL-10 expression was higher in keratinocytes from the thigh compared to controls (C, D). * p < 0.05.
Intergroup difference for keratinocyte nociception-associated ion channel expression, and expression of EFNA4 and EPHA4 in keratinocytes of patients with FMS compared to healthy controls
Gene expression in keratinocytes cultured from the calf and the thigh did not differ between groups for nociception-associated ion channels (Figure 4A, 4B).
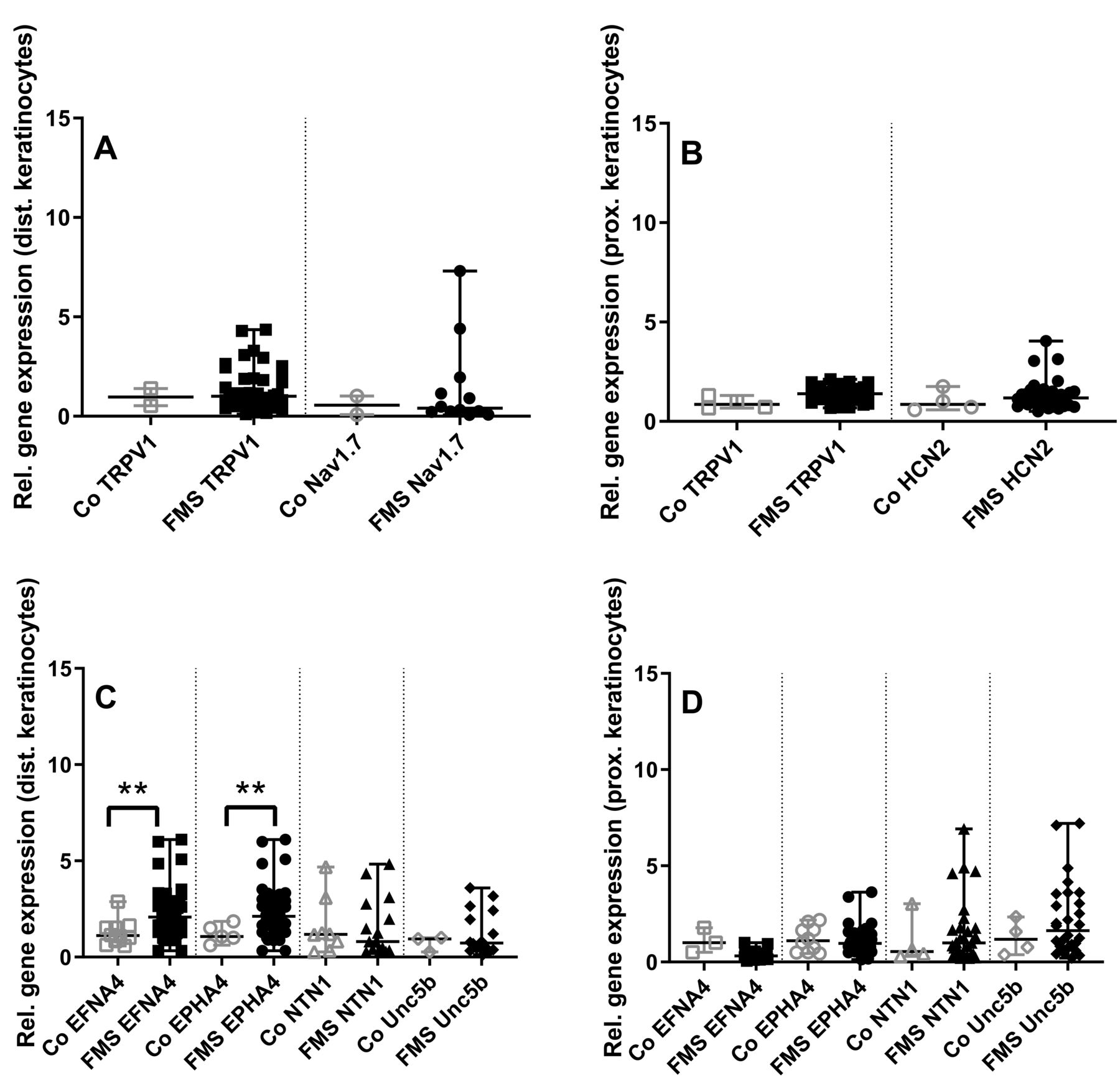
Gene expression panel of ion channels and axon guides in skin keratinocytes of patients with fibromyalgia syndrome (FMS) compared to healthy controls (Co). The scatter plots illustrate gene expression of the pain-associated ion channels transient receptor potential vanilloid-1 (TRPV1), voltage-gated sodium channel (Nav) 1.7, hyperpolarization-activated cyclic nucleotide-gated ion channel-2 (HCN2; panels A, B), and the axon guidance cues and their receptors ephrin-A4 (EFNA4), ephrin receptor-A4 (EPHA4), netrin1 (NTN1), and uncoordinated-5 homolog B (Unc5b; panels C, D). Data were obtained from keratinocytes derived from skin punch biopsies of the calf (A, C) and the thigh (B, D). Gene expression of keratinocytes did not differ between groups for the investigated pain-associated ion channels (A, B). Among axon guidance cues, EFNA4 and EPHA4 expression was higher in keratinocytes obtained from the calf of patients with FMS compared to controls (C, D). ** p < 0.01.
Among the investigated axon guidance cues, there was higher gene expression of EFNA4 and EPHA4 (p < 0.01 each) in keratinocytes obtained from the calf compared to controls (Figure 4C, 4D). No intergroup difference was found in keratinocytes cultured from the thigh.
IENFD is reduced in subgroups of patients with FMS, but does not correlate with axon guidance gene expression
Comparing IENFD at the calf and the thigh of patients with FMS with our laboratory reference values based on 180 healthy controls, we found reduced IENFD only at the calf in 27/127 (21%), only at the thigh in 29/127 (23%), and at the thigh and calf in 29/127 (23%) patients; 1 patient had a normal IENFD at the calf but refused biopsy at the thigh. There were 124 women (median age 50 yrs, 20–84) and 56 men (median age 53 yrs, 22–76). Normal values were calf 9 ± 3 fibers/mm, and thigh 12 ± 4 fibers/mm. Hence, the total number of patients with pathologically reduced distal IENFD was 56/127 (44%). No intergroup difference was found for EFNA4 and EPHA4 gene expression of keratinocytes and the innervation pattern of patients with FMS. Also, axon guide expression did not correlate with IENFD (data not shown).
The obtained qRT-PCR data did not show correlations with the clinical characteristics detailed above.
DISCUSSION
We investigated gene expression of selected pro- and antiinflammatory cytokines, nociception-associated ion channels, and axon guidance cues in primary skin fibroblasts and keratinocytes from patients with FMS compared to healthy controls to allow analysis of the cellular source of each target. We found higher gene expression of TGF-ß1, HCN2, EFNA4, and EPHA4 in fibroblasts, and higher gene expression of IL-10, EFNA4, and EPHA4 in keratinocytes of patients with FMS compared to healthy controls, while gene expression of proinflammatory cytokines did not differ between the groups.
In several studies, systemic gene expression of pro- and antiinflammatory cytokines was investigated in the blood21,22 and cerebrospinal fluid23,24 of patients with FMS. The studies reported elevated IL-8 as one relatively consistent result, but otherwise data were mostly conflicting. Also, the pathophysiological relevance of alterations in expression patterns of single cytokines remains elusive. In contrast, few studies have focused on local cytokine patterns in patient skin samples. TNF-α, IL-1ß, and IL-6 gene expression were detected in skin samples of patients with FMS, but not in healthy controls12. No intergroup difference was found for TNF-α and monocyte chemoattractant protein-1 immunoreactivity between FMS and control skin25. In our own previous study using whole skin punch biopsy samples of patients with FMS and healthy volunteers, we did not find intergroup differences in gene expression for selected pro- and antiinflammatory cytokines13. While subtle differences may remain undetected when using whole skin, the present approach using primary cell cultures of fibroblasts and keratinocytes also failed to show intergroup differences for proinflammatory cytokines. Among antiinflammatory cytokines, we previously showed a reduction in systemic expression of the analgesic and antiinflammatory cytokine IL-4 in patients with FMS26 and also provided evidence for a pathophysiological role of IL-4 deficiency in pain27,28. Here we show a higher expression of TGF-ß1 in fibroblasts and of IL-10 in keratinocytes of patients with FMS compared to controls, while IL-4 expression levels did not differ between cell types and study groups (Figure 1C, 1D; Figure 3C, 3D).
Keratinocytes express nociception-associated ion channels such as TRPV9, voltage-gated sodium channels10, and purinergic receptors29. Fibroblasts also carry respective ion channels30,31,32. These cellular properties together with their close vicinity to cutaneous nerve fibers make keratinocytes and fibroblasts interesting potential contributors to cutaneous nociception33. Hence, we hypothesized that skin cells of patients with FMS may show differential expression of nociception-associated ion channels. We did not, however, find intergroup differences except for a higher HCN2 expression in fibroblasts from the calf of patients compared to controls (Figure 2A).
Axonal pathfinding is mainly regulated by axon guidance cues, which act on the growth cone and may repel or attract nerve fibers6,7. Depending on the expression pattern of respective axon pathfinding proteins and their receptors, tissue innervation may be enhanced or reduced, and nerve fiber composition may be directed. It is also possible that changes in nerve fiber morphology34 may alter the composition of axonal guidance receptors. NTN1 and one of its receptors, Unc5b, are frequently investigated axon guidance cues that lead to axon repulsion35. The ephrin family of receptor tyrosine kinases and their ligands are axon guidance molecules, with EFNA4 attracting and EPHA4 repulsing the axon growth cone36,37. Interestingly, many cell types co-express axon guidance ligands and receptors38, which enables flexible regulation of neurite growth. While expression patterns of NTN1 and Unc5b did not differ between groups, we found a higher expression of EFNA4 and its receptor EPHA4 in fibroblasts and keratinocytes of patients with FMS compared to controls. No correlation was found with patients’ IENFD and skin cell EFNA4 and EPHA4 expression levels, but the imbalance found in gene expression of these axon pathfinding cues may still indicate a relevant role in the pathophysiology of skin innervation, as schematically illustrated in Figure 5. An increase in epidermal expression of repulsive axon guides combined with the expression of attractant axon guidance cues in the dermis may direct cutaneous nerve fiber growth. This may lead to normal or reduced nerve fiber density, as was shown for semaphorin-3A, another axon guide produced by keratinocytes6 in diabetes mellitus11. By proinflammatory properties, axon guidance cues may also be involved in nerve fiber sensitization7. Further studies focusing on protein levels and skin cell–neurite interactions are warranted to answer these questions.
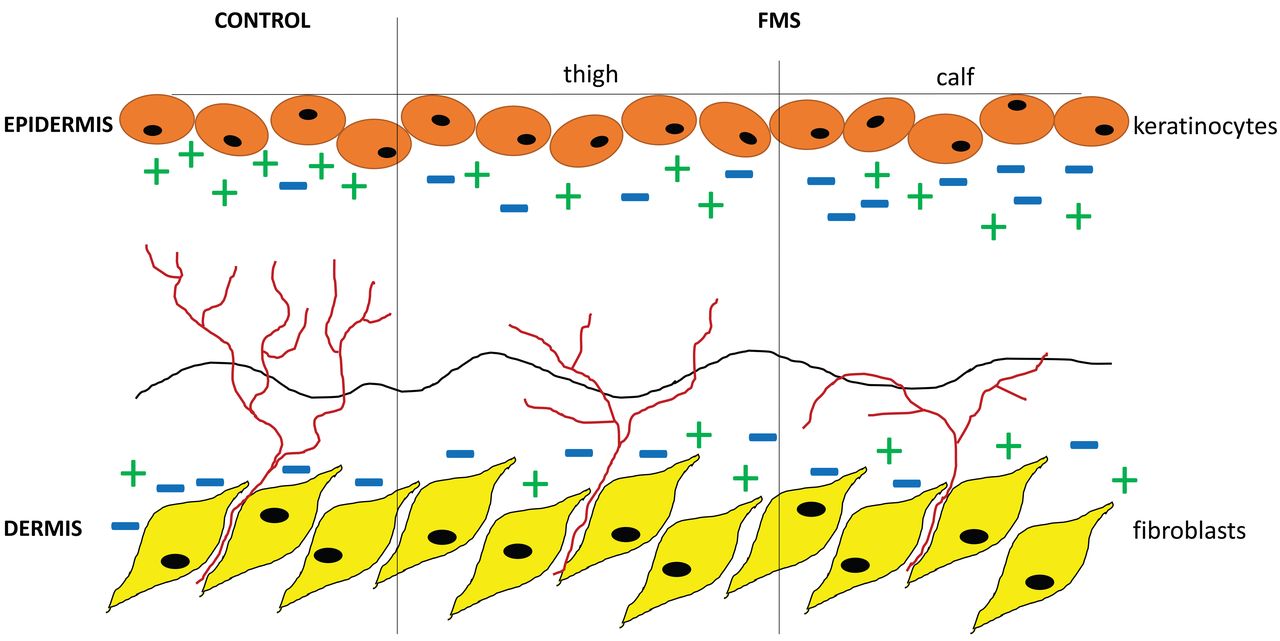
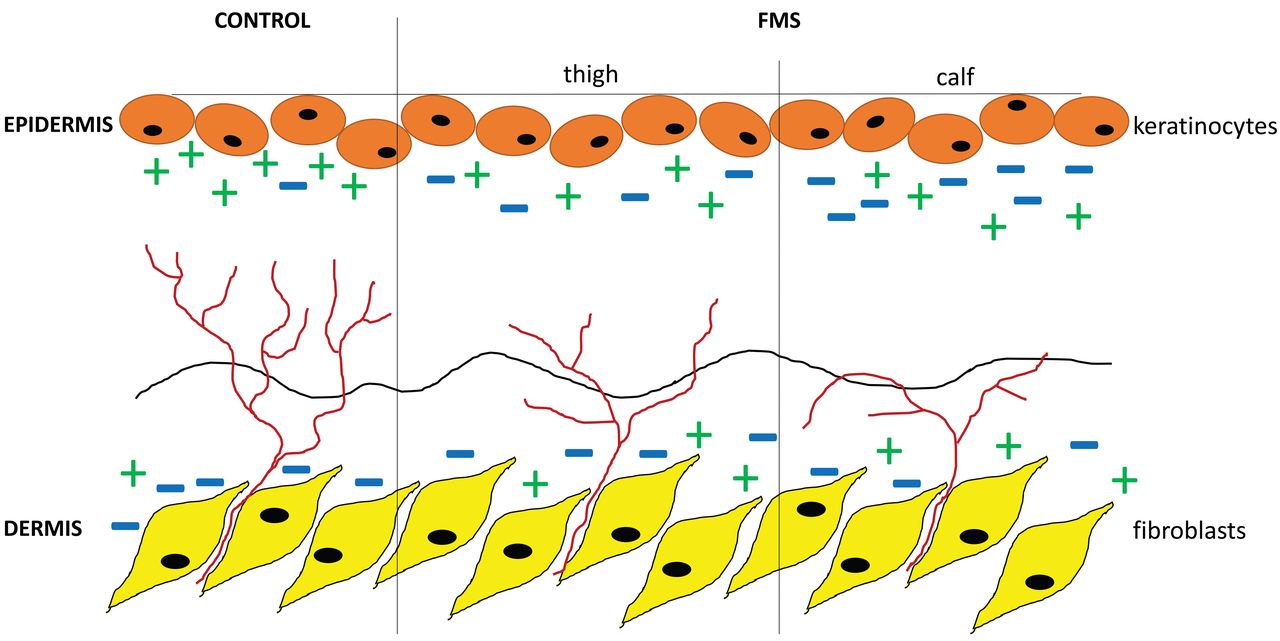
Schematic illustration of a potential mechanism for intraepidermal nerve fiber growth directed by axon guides. In healthy control skin (left), there is a balanced expression level of repellent (−) and attractant (+) axon guidance cues, hence intraepidermal nerve fiber density (IENFD) is normal. In contrast, the expression of axon guidance cues is unbalanced in the skin of patients with fibromyalgia syndrome (FMS), which leads to reduction of IENFD.
One major limitation of our study is that cultured cells do not reflect the actual status of the skin at the time of biopsy and that the culturing process itself may alter mediator production. Hence, confirmation of cell culture results in patient-derived tissue is necessary. We focused on fibroblasts and keratinocytes but did not investigate other potentially involved skin cells such as mast cells because of the challenges in establishing cell culture protocols for 3-mm human skin biopsy samples. We also have used only gene expression as an outcome variable but cannot provide protein data. When interpreting our results, the lack of a disease control group for comparison of data obtained in patients with FMS not only with healthy subjects but also with patients having other pain syndromes needs to be taken into account. Further, we have not controlled our data for patients’ comorbidities and medication, which may influence gene expression. However, these influences would be of relevant effect in systemic biomaterial such as blood or cerebrospinal fluid rather than in skin punch biopsies and even less so in skin cells that undergo an intensive cultivation procedure for several weeks in vitro before being used for RNA extraction.
The strength of our study is that we have investigated a large cohort of patients using individual primary fibroblast and keratinocyte cell cultures directly generated from our patients for differential mediator expression. We also provide a panel of relevant markers that may be used for future studies to investigate the underlying mechanisms of nociception and cutaneous nerve fiber degeneration in patients with FMS. Our results reveal that skin cytokine expression is differentially altered when investigating individual skin cell cultures, that fibroblasts have a broader array of secreted mediators in FMS skin than keratinocytes, and that fibroblasts and keratinocytes may be involved in nerve fiber degeneration. An imbalance in the expression of attractant and repulsive axon guides in the skin may reduce neurite outgrowth from the subepidermal nerve plexus and may thus affect epidermal innervation.
Acknowledgment
Excellent technical help by Daniela Urlaub, Korbinian Niedermüller, and Sonja Mildner is gratefully acknowledged. We also thank Korbinian Niedermüller for his help in designing Figure 5.
APPENDIX 1.
P values of gene expression comparisons between skin cells of patients with FMS and healthy controls.
Footnotes
The study was funded by the Else Kröner-Fresenius-Stiftung (EKFS, NÜ: 2014_A129).
- Accepted for publication May 30, 2019.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}