

Abstract
Objective. Osteoarthritis (OA) is a condition that features inflammation and immune responses of innate and adaptive immunity. The role of T cells in knee OA pathogenesis is still unclear. Our aim was to characterize T cell functions and their clonality in patients with knee OA in peripheral blood (PB) and infrapatellar fat pads (IPFP).
Methods. We isolated T cells from PB and IPFP of patients with knee OA and PB of healthy individuals and determined soluble mediators produced from these cells. In addition, we performed a clonal analysis of activated CD8+ T cells and compared the T cell receptor β-variable gene chain (TRBV) usages between T cells in PB and IPFP of patients with knee OA.
Results. Our results suggest that in patients with knee OA, circulating T cells possess a more “cytotoxic” profile or rather impaired cytokine production, but the knee microenvironment allows for these T cells to produce proinflammatory cytokines [interleukin (IL)-1β, IL-6, tumor necrosis factor], IL-17, and interferon-γ within IPFP. Activated CD8+ IPFP T cells carry different repertoire distribution from those present in PB of patients with knee OA. Shared TRBV usage of activated CD8+ IPFP T cells among the 3 patients with knee OA was also observed.
Conclusion. Our study describes the nature of T cells in knee OA that may be due to “unhealthy” aging or other factors that drive healthy aging T cells into a state of imbalance, thus contributing to the pathogenesis of knee OA.
Osteoarthritis (OA) is a debilitating disease featured by periarticular tissue dysfunction and inflammation of surrounding tissues, e.g., fat pads1,2. It was originally described as a degenerative condition1,3 from biomechanical loading of the joints, but current evidence suggests immune responses may also be involved in OA development4,5,6.
Immune responses related to knee OA include both innate and adaptive immune compartments7,8,9,10. Low-grade inflammation is well described in knee OA, with presence of soluble proteins and inflammatory mediators in synovial fluids5,11,12. Infiltration of inflammatory immune cells within the infrapatellar fat pad (IPFP) and synovium are also key findings in knee OA pathology13,14,15. The IPFP (or Hoffa’s pad) is an intraarticular and intracapsular adipose tissue that serves as a local producer of adipocytokines under the influence of local stimuli13,16,17. It has been proposed that IPFP may serve for both biomechanical (pressure absorption) and biochemical (inflammatory mediator secretion) purposes in maintaining the integrity of the knee joint18. Tissue composition of IPFP includes adipocytes, immune cells, nerve tissues, and connective tissue13 and resembles that of visceral adipose tissue13. IPFP volume may affect knee OA development, but studies are still being debated19,20,21,22,23. Regardless, the IPFP is a site where inflammatory cells can accumulate, including macrophages, T cells, B cells, mast cells, and others24,25. In our study we mainly focus on IPFP T cells.
T cells are the second most abundant immune cells within IPFP25. There are many studies investigating the role of T cells in the pathogenesis of osteoarthritis10, but fewer studies of IPFP-infiltrating T cells24,25. Mice lacking CD4+ or CD8+ T cells show a decrease in severity of knee OA, suggesting their role in driving knee OA pathology26,27. However, the mechanisms are still undefined. Thus, it was in our interest to investigate T cells from IPFP and peripheral blood of patients with knee OA. In this study, we characterized T cell function by determining the levels of cytokine and cytotoxic molecule production from IPFP T cells and peripheral blood T cells of patients with knee OA. In addition, we analyzed the T cell receptor (TCR) repertoire of IPFP and peripheral blood T cells of these patients.
MATERIALS AND METHODS
Patient recruitment and tissue sample collection
Patients with knee OA and healthy individuals were recruited from King Chulalongkorn Memorial Hospital. Peripheral blood and IPFP were obtained from patients with knee OA undergoing total knee arthroplasty, while peripheral blood came from healthy individuals. For ethical reasons, we were not able to obtain healthy IPFP from living donors, and access to postmortem IPFP was limited. Procedures were performed in accordance with the ethical standards and with approval from the committee on human experimentation (Institutional Review Board) at the Faculty of Medicine, Chulalongkorn University, Bangkok, Thailand (IRB no. 574/57) and with the Helsinki Declaration of 1975, as revised in 2000. All samples were obtained with the patient’s/individual’s signed consent. Tissue samples obtained were processed immediately.
Peripheral blood mononuclear cell isolation
Whole blood was layered on to Ficoll-Paque (GE Healthcare) and centrifuged at 2000 rpm for 20 min at room temperature (no deceleration force). Cells were washed twice with RPMI 1640 (Life Technologies) supplemented with 10% fetal calf serum (FCS; Life Technologies) and cryopreserved in 90%FCS/10% dimethyl sulfoxide (DMSO; Amresco) until experiments were performed.
IPFP T cell isolation
Fat pads were washed with phosphate buffer saline (PBS), cut into 2–3 mm pieces, and digested with 3 µg/ml collagenase type IV (Worthington), 0.1 µg/ml DNase I (Worthington), and 5% FCS in PBS with a shaking force of 200 rpm, 37°C for 90 min. The supernatant was filtered through a 40-µm filter and washed with RPMI 1640 (Life Technologies) supplemented with 10% FCS and cryopreserved in 90%FCS/10% DMSO (Amresco).
Ex vivo T cell analysis. For cytokine detection, 105 mononuclear cells were cultured in the presence of anti-CD3/anti-CD28 beads (Thermofisher Scientific) and brefeldin A (BFA) added (final 20 µg/ml) and incubated at 37°C for 16–18 h. Cells were labeled with anti-CD3-PE-Cy7 (clone UCHT1), anti-CD4-PE-Cy5 (clone RPA-T4), and anti-CD8-PerCp/Cy5.5 (clone SK1) antibodies at 4°C for 30 min, then fixed with 1% formaldehyde in PBS at 4°C for 30 min and further labeled with anti-interleukin (IL)-17-PE (clone SCPL1362), anti-IL1-β-FITC (clone H1b-98), anti-IL-6-APC (clone MQ2-13A5), anti-interferon (IFN)-γ-APC-Cy7 (clone 4S.B3), and anti-tumor necrosis factor (TNF)-AlexaFluor 700 (clone MAb11) antibodies in 0.3% saponin (Sigma-Aldrich).
For cytotoxic molecule detection, 105 cells were initially labeled with anti-CD107a-PE-Cy7 (clone H4A3) antibody and stimulated with anti-CD3/anti-CD28 beads (Thermofisher Scientific) for 1 h, then BFA (final 20 µg/ml) and monensin (final 20 µg/ml) were added and incubated at 37°C for 16–18 h. Cells were washed and labeled with anti-CD3-APC (clone UCHT1), anti-CD4-APC-Cy7 (clone RPA-T4), and anti-CD8-AF700 (clone SK1) antibodies at 4°C for 30 min, then fixed with 1% formaldehyde in PBS and labeled with antigranzyme B-FITC (clone GB11), anti-perforin-PerCp/Cy5.5 (clone SG9), and anti-granulysin-PE (clone DH2) antibodies in 0.3% saponin (Sigma-Aldrich). All antibodies were purchased from Biolegend. Cells were then acquired on the BD LSRII flow cytometer. Analysis was performed using the Flowjo software (Treestar; Supplementary Figures 1–3, available with the online version of this article).
TCR repertoire determination
Cells were labeled with anti-CD3-APC (clone UCHT1), anti-CD8-AF700 (clone SK1), and anti-CD69-PE (clone FN50; Biolegend) antibodies. CD3+CD8+CD69+ cells were single cell-sorted into 96-V well PCR plates using the BD FACS Aria II-cell sorter. cDNA was synthesized using SuperScript VILO cDNA Synthesis Kit (Invitrogen) in 2.5 µl reaction mixes as the manufacturer’s description. Products were amplified by multiplex nested PCR for TCR transcripts. Primers were used as previously described28. In the first round, 2.5 µl of cDNA was added to 2.5 µl of 10× PCR buffer, 1.5 mM MgCl2, 0.2 mM deoxynucleotide triphosphate, 0.75 U of Taq DNA polymerase, and 2.5 pmol each of the external sense (TRBV and TRAV) and external antisense (TRBC and TRAC) primers28. Then, 5 µl of PCR products were used as templates for subsequent rounds replacing the primers with internal sense TRAV primers and internal antisense TRAC primers; or internal sense TRBV primers and internal antisense TRBC primers28. PCR reaction mixes were incubated at 95°C for 2 min, following 35 cycles of 95°C for 20 s, 52°C for 20 s, 72°C for 45 s, and 1 cycle of 72°C for 7 min. DNA products were detected by 2% agarose gel electrophoresis and amplified using internal antisense TRAC or TRBC primer after treatment with Illustra ExoProStar 1-Step Kit (GE Healthcare). Data were analyzed using the ImMunoGeneTics database.
Statistical analysis
All data were analyzed using the Statistical Package for Social Sciences (SPSS 15.0, SPSS Inc.) or Graph Pad InStat version 5.0 software. The results were presented as mean ± SD. Independent sample t tests was used to compare the value between 2 groups (between healthy control and patients with knee OA; and peripheral blood and IPFP of patients with knee OA). Statistical significance was defined as p ≤ 0.05, p ≤ 0.01, or p ≤ 0.001.
RESULTS
Demographic characteristics of studied populations
Patients with knee OA (n = 31) and healthy individuals (n = 11) were included in the study (Table 1). Demographic characteristics showed mean values (± SD values) of age, sex distribution, weight, height, and body mass index (BMI; Table 1). The mean age, weight, and BMI of patients with knee OA was higher than those of healthy individuals, with both populations having comparable mean height (Table 1).
Demographic characteristics between healthy individuals and osteoarthritic patients.
Peripheral blood T cell cytokine and cytotoxic mediator profiling of patients with knee OA
Inflammation in knee OA is characterized by an imbalance between catabolic and anabolic cytokines29. Among the catabolic cytokines, IL-1β, IL-6, and TNF are well-described cytokines related to knee OA30,31 and can be secreted from T cells32,33. In addition, pathogenic IL-17-producing Th cells have been shown to be pivotal in the pathogenesis of rheumatoid arthritis34. IFN-γ is a T cell signature cytokine secreted in response to antigenic stimulation, in which autoantigens have been proposed in knee OA35. For these reasons, we evaluated for T cell production of IL-1β, IL-6, TNF, IL-17, and IFN-γ from patients with knee OA and healthy individuals. First, peripheral blood T cells were isolated and evaluated for their cytokine production ex vivo (unstimulated). We observed a significant increase in TNF production from CD4+ T cells and significant decreases in IL-6, TNF, and IFN-γ production from CD8+ T cells (Figure 1A). When these cells were stimulated with anti-CD3/anti-CD28 beads, cytokine level production between patients with knee OA and healthy individuals was mostly comparable, with only IL-6-producing and IFN-γ-producing CD8+ T cells in patients with knee OA showing significantly lower levels of cytokine production than those in healthy individuals (Figure 1A). These findings may suggest an impaired function in IL-6 and IFN-γ production from peripheral blood T cells in patients with knee OA compared to healthy individuals.

Peripheral blood T cell cytokine and cytotoxic molecule production of patients with knee OA and healthy individuals. Peripheral blood T cells from knee OA patients and healthy individuals were isolated and stimulated with anti-CD3/anti-CD28 beads. Intracellular cytokine staining and degranulation assay was performed to evaluate the levels of T cells producing the following: IL-1β, IL-17, IL-6, TNF, IFN-γ, perforin, granzyme B, and granulysin. T cells were grouped based on their coreceptor expression of CD4 or CD8. In all settings, T cells were evaluated in 2 conditions: (1) ex vivo (unstimulated) and (2) stimulated with anti-CD3/anti-CD28 beads. Shown are mean values. Error bars represent SD values. Healthy individuals (white bars), n = 11; patients with knee OA (black bars), n = 31. * p ≤ 0.05, ** p ≤ 0.01, *** p ≤ 0.001. A. Percentage of cytokine+ T cells from total CD4+ (or CD8+) T cells in unstimulated (top panel) and stimulated (bottom panel) conditions. B. Percentage of CD107a+CD4+ and CD107a+CD8+ T cells from total CD4+ and total CD8+ T cells, respectively, in unstimulated (top panel) and stimulated (bottom panel) conditions. C. Percentage of cytotoxic molecule production from total CD107a+CD4+ or CD107a+CD8+ T cells in unstimulated (top panel) and stimulated (bottom panel) conditions. Prf: perforin; GrB: granzyme B; GNLY: granulysin; IL: interleukin; TNF: tumor necrosis factor; OA: osteoarthritis; IFN: interferon.
A feature of knee OA is cartilage destruction9. We hypothesized that chondrocytes may serve as target cells for T cell–mediated cytotoxicity. Therefore, we evaluated production of cytotoxic molecules secreted from peripheral blood T cells of patients with knee OA. Degranulation levels of T cells were also determined, as represented by cell surface expression of CD107a. Degranulation levels of ex vivo CD4+ and CD8+ T cells in patients with knee OA and healthy individuals were comparable (Figure 1B). However, after anti-CD3/anti-CD28 bead stimulation, cell surface expression of CD107a of knee OA peripheral blood CD4+ T cells were not upregulated to levels comparable to those in healthy individuals (Figure 1B). Thus, we further selected only degranulating cells (CD107a+ cells) to determine cytotoxic molecule production. Our results showed that ex vivo CD8+CD107a+ T cells expressed significantly higher levels of perforin production. After T cell stimulation with anti-CD3/anti-CD28 beads, perforin and granulysin production of peripheral blood CD4+ and CD8+ T cells from patients with knee OA were increased when compared to healthy individuals, with significant increases seen in CD107a+CD4+perforin+, CD107a+CD4+ granulysin+, and CD107a+CD8+granulysin+ T cells (Figure 1C). The inability of CD4+ T cells to express levels of cell surface CD107a comparable to healthy individuals along with increased abundance of perforin and granulysin production after stimulation suggests that peripheral blood CD4+ T cells in patients with knee OA may be constantly polarized to cytotoxic CD4+ T cells. However, CD8+ T cells from patients with knee OA still exhibited their ability to be activated by anti-CD3/anti-CD28 bead stimulation and also showed a cytotoxicity profile. Altogether, our results here suggest that peripheral blood T cells in patients with knee OA are biased toward a cytotoxicity profile.
A comparison of IPFP T cells and peripheral blood T cells in patients with knee OA
The IPFP is the most adjacent site of adipose tissue to the knee joint and is described as a site of inflammatory cell infiltration25. Thus, we further evaluated T cells isolated from IPFP for their cytokine and cytotoxic molecule production. Owing to the limited number of isolated cells, we determined only the levels of ex vivo (unstimulated) conditions. Our study also lacks healthy IPFP for direct comparison.
Our results show that both CD4+ and CD8+ IPFP T cells had significantly higher levels of IL-1β, IL-6, and TNF production than did peripheral blood T cells (Figure 2A). In addition, we observed significantly higher levels of IFN-γ production in CD8+ T cells (p = 0.003) and significantly higher levels of IL-17 production in CD4+ T cells (Figure 2A). Cell surface expression of CD107a revealed higher levels of degranulation of IPFP CD4+ T cells (p < 0.0001) than peripheral blood of patients with knee OA, but levels of CD107a for CD8+ T cells between the 2 tissues were comparable (Figure 2B). We then determined cytotoxic molecule production of IPFP T cells regardless of their cell surface CD107a. IPFP T cells exhibited increased levels of cytotoxicity as shown with significant increases of perforin-secreting and granulysin-secreting CD4+ T cells and perforin producing and granzyme B-producing CD8+ T cells (Figure 2C). These findings reflect T cells as potential sources of cytokines (TNF, IL-6, IL-1β, IL-17, and IFN-γ) and cytotoxic molecules (perforin, granzyme B, and granulysin) within IPFP. However, the extent of their contribution to local inflammation within the knee joint requires further evaluation. We further assessed the ability of cytokine-producing T cells to produce multiple cytokines. Our results show that the majority of IPFP T cells secreted only 1 cytokine; IL-1β and IL-6 were the most dominant (Figure 2D). A very few dual-cytokine secreting T cells were observed (Figure 2D).

Ex vivo T cell cytokine and cytotoxic molecule production in IPFP of patients with knee OA compared with patient-matched peripheral blood. T cells were isolated from peripheral blood and IPFP of patients with knee OA. Intracellular cytokine staining and degranulation assay was performed to evaluate the levels of T cells producing IL-1β, IL-17, IL-6, TNF, IFN-γ, perforin, granzyme B, and granulysin. T cells were grouped based on their coreceptor expression of CD4 or CD8. Because of the limited number of cells isolated from IPFP, only ex vivo conditions were used to determine the levels of cytokine-producing and cytotoxic molecule–producing T cells. Not all patients were matched between both tissues. Shown are mean values. Error bars represent SD values. Peripheral blood (white bars), n = 26; IPFP (black bars), n = 17. *p ≤ 0.05, ** p ≤ 0.01, *** p ≤ 0.001. Prf: perforin; GrB: granzyme B; GNLY: granulysin; IL: interleukin; TNF: tumor necrosis factor; OA: osteoarthritis; IFN: interferon; PBMC: peripheral blood mononuclear cells; IPFP: infrapatellar fat pads.
Clonal analysis of activated CD8+ T cell usage in patients with knee OA
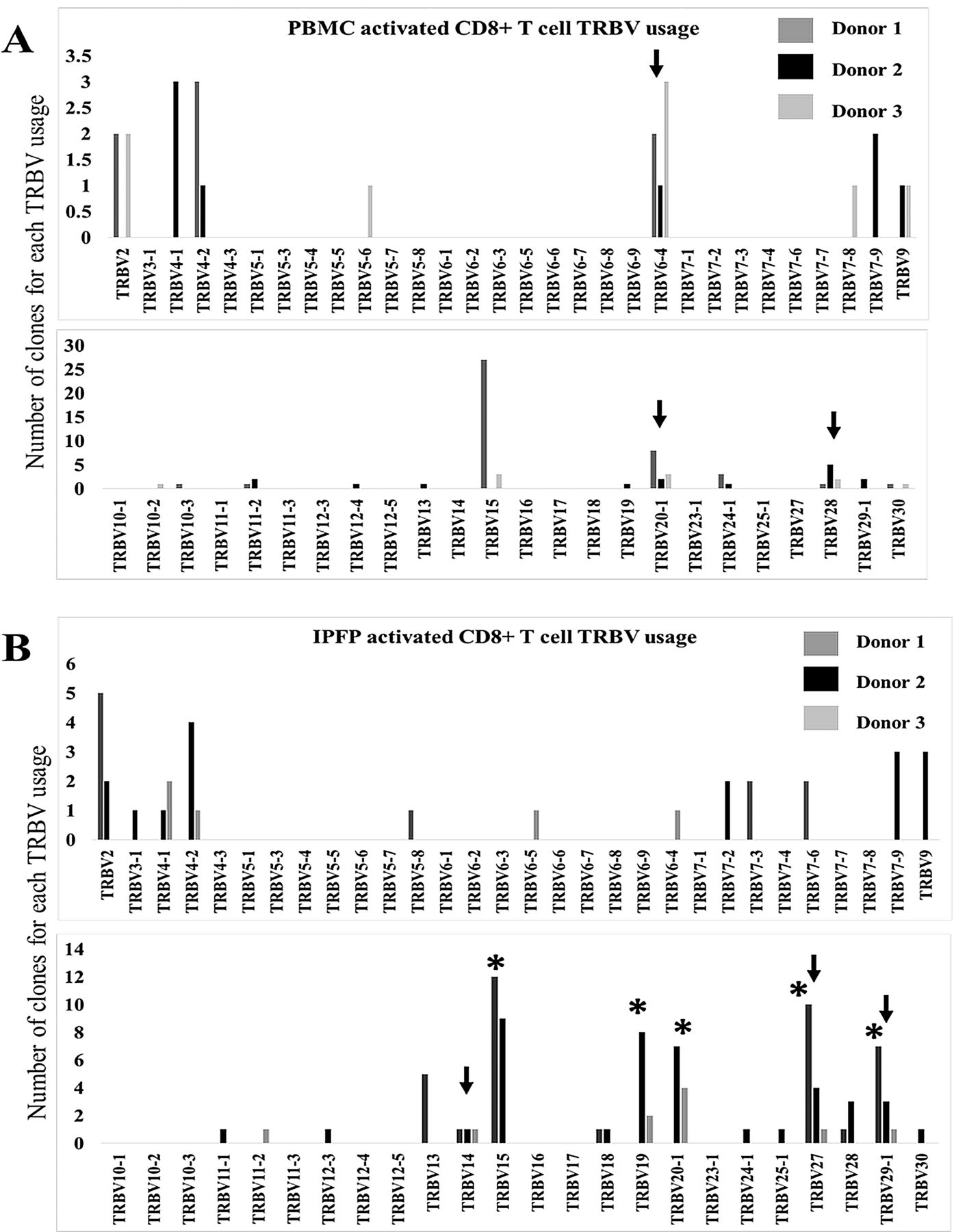
Knee OA is correlated with increased BMI values36, but not directly with weight37,38. Increased body weight is linked to alterations of adipose tissue function and metabolic states described to be associated with OA prevalence39,40. Mice fed a high-fat diet, which leads to increased weight, had initial migration of CD8+ T cells to adipose tissues, followed by other immune cells41. These adipose tissue T cells also display a restricted TCR repertoire42. Previously, we have shown that in patients with knee OA, circulating CD8+ T cells were more activated than CD4+ T cells, and increased frequency of IPFP CD8+ T cells was observed with higher radiographic grading of knee OA15. In addition, Ponchel, et al detected a correlation between increasing numbers of peripheral blood CD8+ T cells, but not CD4+ T cells, with age in osteoarthritic patients43. Thus, we investigated the TCR repertoire of activated CD8+ T cells in the IPFP of patients with OA and compared them to peripheral blood T cell clones by performing a clonal analysis of β-chain usages of CD8+CD69+ T cells in both tissues from 3 patients. Clonal analysis revealed that activated CD8+ T cells in IPFP have different repertoire patterns of TRBV usage from the peripheral blood in each individual (Figure 3A). The repertoire profiling showed that each donor also contained different predominant TRBV usage between IPFP and peripheral blood (Figure 3A and 3B). Peripheral blood activated CD8+ T cells were dominated by TRBV 15 and 20-1; TRBV 28 and 4-1; and TRBV 6-4, 20-1, and 15; for donors 1, 2, and 3, respectively (Figure 3A; Table 2). IPFP activated CD8+ T cells were dominated by TRBV 15, 27, 29-1, 2, and 13; TRBV 15, 19, and 20-1; and TRBV 20-1, 4-1, 19, and 29-1 for donors 1, 2, and 3, respectively (Figure 3A, Table 2). Comparison of activated CD8+ T cell clones in peripheral blood and IPFP among the 3 patients (interdonor comparison) demonstrates that in peripheral blood, shared TRBV usage was seen in TRBV 6-4, 20-1, and 28 (Figure 4A), but these did not represent the shared TRBV usage present in IPFP. We observed that IPFP-activated CD8+ T cells between each patient with knee OA possessed overlapping TRBV usages that were present as predominant clones and those that were not predominant (present in 2 or more patients; Figure 4B, Table 2). In IPFP, shared TRBV usage included TRBV 14, 15, 27, and 29-1 (Figure 4B). With the number of clones that we obtained, the data show that activated CD8+ T cells in IPFP differ from those in peripheral blood of the same individual. Also, the shared TRBV usages observed in activated CD8+ IPFP T cells supports the notion of a common driver among patients with knee OA in specific CD8+ T cell recruitment or proliferation. These drivers would more likely be stimuli local to the joint because peripheral blood–activated CD8+ T cells do not reflect the repertoire hierarchy in IPFP.
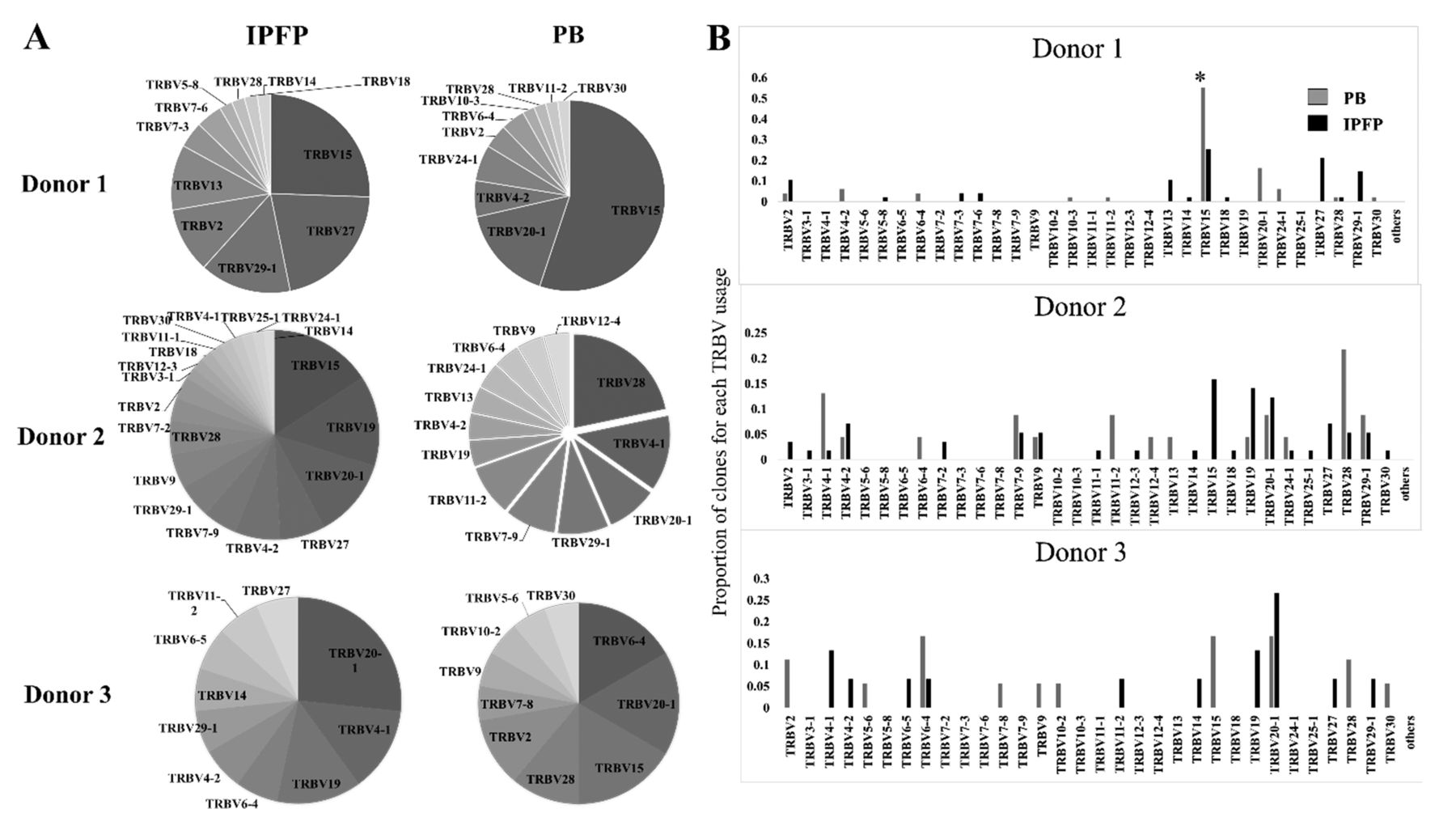
TRBV usage of activated (CD69+) CD8+ T cells in peripheral blood and IPFP of 3 patients with knee OA. Number of clones: Donor 1, PB n = 23, IPFP n = 59; Donor 2, PB n = 49, IPFP n = 47; Donor 3, PB n = 21, IPFP n = 16. A. Proportions of each TRBV usage in IPFP and peripheral blood of donors 1, 2, and 3. B. Shared TRBV usage by activated CD8+ T cells in peripheral blood and IPFP in each donor. * This TRBV15 usage by Donor 1 contained clones that were identical (same CDR3-β) in 25/27 clones. X axis represents each TRBV. Y axis represent the proportion of clones for each TRBV usage. OA: osteoarthritis; PB: peripheral blood; IPFP: infrapatellar fat pads; TRBV: T cell receptor β-variable gene chain.

Common shared TRBV usage among 3 patients with knee OA in activated CD8+ T cells of peripheral blood and IPFP. A. TRBV usage of peripheral blood activated CD8+ T cells from 3 patients with knee OA. Black arrows represent TRBV chains that are shared among all 3 donors. B. TRBV usage of IPFP-activated CD8+ T cells from 3 patients. Black arrows represent TRBV chains that are shared among all 3 donors. Asterisks (*) represent TRBV that were selected for their dominant presence in IPFP. TRBV: T cell receptor β-variable gene chain; OA: osteoarthritis; IPFP: infrapatellar fat pads; PBMC: peripheral blood mononuclear cells.
Predominant TRBV usage of activated (CD69+) CD8+ T cells from patients with knee OA, from peripheral blood and IPFP.
DISCUSSION
T cells are effector cells of the adaptive immune response that function by proliferation, cytokine production, cytotoxicity, and differentiation44 and have been described in knee OA because of their presence in synovial fluids, synovium linings, and IPFP10,13,25. Our study focused on understanding the role of T cells in IPFP and peripheral blood of patients with knee OA by comparing their phenotypes to healthy individuals. In addition, we also compared the TCR repertoire of IPFP and peripheral blood T cells in 3 patients with knee OA.
Here, we show that in peripheral blood of patients with knee OA, there was a tendency of CD4+ and CD8+ T cells to have lower cytokine production and an increase in cytotoxic molecule production. Despite previous studies showing that increased frequency of CD8+ T cells is related with knee OA with increasing age, our data suggest that T cells driving inflammation in knee OA may also have altered function in addition to disproportionate CD4+/CD8+ imbalances. Aging causes a decline in immune function referred to as immunosenescence45 and is accompanied by low-grade inflammation of the host46. Because of differences between healthy individuals and patients with knee OA in our study, interpretation of results should take this factor into account. T cells are most severely affected and have increased CD8+ T cell clonality because of the decline in CD4+ T cell survival, presence of chronic simulation by latent viral infection, thymic involution, and accumulation of immune exposure history45. In other words, aging, along with additional driving forces, may polarize such CD4+ T cells to a declining state with dysfunctional phenotypes in knee OA.
Immune function profiling of IPFP T cells suggests their capability in producing proinflammatory cytokines (IL-1β, IL-6, TNF), IL-17, and IFN-γ. Whether this cytokine production from T cells is increased in IPFP of knee OA is still inconclusive owing to our lack of healthy IPFP from living individuals and postmortem samples. However, it does suggest that the local microenvironment plays a role in driving CD4+ and CD8+ T cells to secrete these cytokines, especially IL-1β and IL-6, whose secretions are tightly regulated32,33,47,48. A scenario in which T cells do secrete IL-6 is present in patients with human T cell lymphotrophic virus infection48. However, it is unknown whether certain viral infections are related directly to knee OA, although there are some studies relating viral infections of osteoblasts found in patients with OA49. Regardless, the extent to which IPFP T cells are contributing to knee inflammation, and whether there are antigen-specific T cells within the joints, need further investigation. If T cells are indeed more stringent with their response to inflammatory microenvironment, then T cells may serve as potential indicators of inflammation in knee OA.
CD8+ T cells are the first immune cells to migrate to adipose tissue in response to increased weight and also function as recruiters of macrophages41. Thus, it was important to investigate the TCR repertoire of IPFP CD8+ T cells. In support of oligoclonal T cell analysis previously reported in the synovium50, our clonal analysis from 3 patients with knee OA revealed that activated CD8+ T cells within IPFP carry a much narrower repertoire than activated CD8+ T cells in peripheral blood. In the peripheral blood of 1 patient, there was a very predominant usage of TRBV15. It was also interesting to observe that TRBV15 usage was present in high proportions among IPFP activated CD8+ T cells in at least 2 of the 3 donors tested. In addition, we also observed common TRBV chains that were shared among the 3 IPFP activated CD8+ T cell repertoire. Age, being a risk factor for knee OA, may also indirectly influence the TCR repertoire in patients with knee OA because chronic infection in elderly patients is a key factor driving the T cell landscape to bias toward selected clones45. An increase in weight may also contribute to this phenomenon; in adipose tissues of obese mice, limited TCR diversity is observed42. However, the predominant β-chain usage in peripheral blood differed from that of IPFP T cells, suggesting a selective pressure for CD8+ T cells that are accumulated in the IPFP. Regardless, more studies are required to distinguish whether the selected clones present in IPFP are a result of migration of selected clones or local clonal selection and expansion that occurs within the IPFP.
We show here that T cells are capable of producing soluble mediators locally and are biased toward cytotoxicity. Also, activated CD8+ T cells within the IPFP contained a limited repertoire diversity when compared to peripheral blood T cells (Supplementary Figure 4, available with the online version of this article). Unknown forces driving the selection, recruitment, and altered function of T cells within the knee joint need to be further studied.
ONLINE SUPPLEMENT
Supplementary material accompanies the online version of this article.
Acknowledgment
We thank the participating patients and clinical staff members at the orthopedics ward for providing and collecting samples that were used in this study.
Footnotes
This project was supported by Faculty of Medicine Ratchadapiseksompot Grant, Grant no. RA58/054 and the Thai Research Fund, Grant no. MRG6080122. TS is an MS student supported by the Chulalongkorn University Graduate Scholarship to Commemorate the 72nd Anniversary of His Majesty King Bhumibol Adulyadej.
- Accepted for publication July 19, 2018.








{kind=link}
{kind=link}
{kind=link}
{kind=link}