

Abstract
Objective. To determine whether levels of plasma n-3 polyunsaturated fatty acids are associated with response to antitumor necrosis factor (anti-TNF) agents in rheumatoid arthritis (RA), and whether this putative effect may have its basis in altering anti-TNF–driven Th17 cell differentiation.
Methods. Plasma was collected at baseline and after 3 months of anti-TNF treatment in 22 patients with established RA, and fatty acid composition of the phosphatidylcholine (PC) component was measured. CD4+CD25− T cells and monocytes were purified from the blood of healthy donors and cocultured in the presence of anti-CD3, with or without etanercept (ETN), eicosapentaenoic acid (EPA), or the control fatty acid, linoleic acid (LA). Expression of interleukin 17 and interferon-γ was measured by intracellular staining and flow cytometry.
Results. Plasma PC EPA levels and the EPA/arachidonic acid ratio correlated inversely with change in the Disease Activity Score at 28 joints (DAS28) at 3 months (−0.51, p = 0.007 and −0.48, p = 0.01, respectively), indicating that higher plasma EPA was associated with a greater reduction in DAS28. Plasma PC EPA was positively associated with European League Against Rheumatism response (p = 0.02). An increase in Th17 cells post-therapy has been associated with nonresponse to anti-TNF. ETN increased Th17 frequencies in vitro. Physiological concentrations of EPA, but not LA, prevented this.
Conclusion. EPA status was associated with clinical improvements to anti-TNF therapy in vivo and prevented the effect of ETN on Th17 cells in vitro. EPA supplementation might be a simple way to improve anti-TNF outcomes in patients with RA by suppressing Th17 frequencies.
Antitumor necrosis factor (anti-TNF) agents have revolutionized the management of severe rheumatoid arthritis (RA), but a substantial proportion of patients either fail to respond or do so only partially1. The reasons for this remain to be fully elucidated, but are likely to include genetic, genomic, and environmental factors2. In some patients, inadequate response may be mediated by an anti-TNF–driven rise in proinflammatory Th17 cell differentiation3. Understanding the reasons for inadequate response may guide choice of an alternative biological agent for some patients or optimization of anti-TNF therapy for others through alteration of environmental factors or selection of synergistic agents.
Long-chain n-3 polyunsaturated fatty acids (PUFA), especially eicosapentaenoic acid (EPA; 20:5n-3) and docosahexaenoic acid (DHA; 22:6n-3), have long been considered to have antiinflammatory and immunomodulatory actions, and several explanatory mechanisms have been proposed. These include modulation of eicosanoid metabolites by competition with arachidonic acid (AA) and inhibition of cyclooxygenase, alteration of lipid rafts, signaling through receptors such as GPR120 and PPAR-γ, and generation of proresolution mediators4,5. N-3 (also known as omega-3) PUFA derive their name from having their first double-bond at the third carbon atom from the methyl end of their fatty acyl chain6. They cannot be synthesized de novo by animals and most human dietary intake is in the form of plant-derived α-linolenic acid (ALA; 18:3n-3)7,8. ALA is metabolized by a series of desaturation and elongation steps to the longer chain EPA and DHA. This process of conversion into longer-chain PUFA is poor in humans9, and is in direct competition with the desaturation and elongation of the considerably more abundant n-6 (omega-6) PUFA. Therefore, in humans, most EPA and DHA is of dietary origin, with marine foods being especially good sources, but having a highly variable intake.
Fish oils contain EPA and DHA. Fish oils ameliorate collagen-induced arthritis10,11,12, and demonstrate modest effects on RA, with a consistent reduction in tender joint count (TJC), morning stiffness, and nonsteroidal antiinflammatory drug (NSAID) usage being observed in multiple small clinical trials when added to conventional disease-modifying antirheumatic drug (DMARD) therapy13–22,23,24,25. Studies suggest that higher doses of fish oil (e.g., 3–6 g/day) are more effective than lower doses. Beneficial effects continue to be seen when fish oil is added to triple DMARD therapy with a treat-to-target approach in early RA26.
In this study, we sought to determine whether plasma levels of n-3 fatty acids correlated with response to anti-TNF in RA, and propose and test a mechanistic hypothesis for the heightened responsiveness we observed in association with higher EPA levels in the absence of high-dose fish oil supplementation.
MATERIALS AND METHODS
Patients
Plasma was available for 22 out of 23 patients with established RA who participated in a comparative study of etanercept (ETN) and infliximab (IFX)27. ETN is a p75 TNF receptor-Fc fusion protein and IFX is a chimeric monoclonal antibody to TNF. Eligible patients were aged ≥ 18 years, fulfilled the 1987 American College of Rheumatology classification criteria for RA, were positive for rheumatoid factor or anticyclic citrullinated peptide antibodies, had disease duration > 6 months and 28-joint Disease Activity Score (DAS28) > 4.0, had previously failed at least 1 DMARD, and were receiving a stable dose of at least 7.5 mg weekly of methotrexate (MTX). No other DMARD were allowed within the 4 weeks prior to commencing treatment. Participants were randomized to standard dosing of either IFX 3 mg/kg intravenously at weeks 0, 2, 6, and 10, or subcutaneous ETN 25 mg twice weekly. Therapy was kept stable for the first 3 months of treatment. The demographics of these patients are presented in Table 1. The study was conducted in compliance with the Helsinki Declaration and ethical approval was obtained from the West Glasgow Ethics Committee (06/S0703/64). All subjects gave written informed consent. The clinical trial registration number is ISRCTN44880063.
Patient demographics at study entry. P values compare the IFX and ETN arms. Values are median (interquartile range) unless otherwise specified.
Plasma phosphatidylcholine fatty acids
Plasma samples from baseline and 3 months were stored at −80°C prior to analysis. After addition of dipentadeconoyl phosphatidylcholine as an internal standard, total lipids were extracted with chloroform and methanol (2:1 vol/vol) and butylated hydroxytoluene added as an antioxidant. Phosphatidylcholine (the major phospholipid in human plasma) was isolated by solid-phase extraction on aminopropyl silica cartridges using chloroform to elute triacylglycerol and cholesteryl ester fractions prior to elution of phosphatidylcholine with chloroform/methanol (60:40 vol:vol). Purified phosphatidylcholine was dissolved in toluene and fatty acid methyl esters were produced by reaction with methanol containing 2% (vol/vol) sulfuric acid at 50°C for 2 h. After cooling and neutralization, fatty acid methyl esters were extracted into hexane and separated on a BPX-70 column fitted to a HP6980 gas chromatograph (Hewlett-Packard). Helium was used as the running gas and fatty acid methyl esters were detected by flame ionization, identified by comparison with previously run standards, and quantified using ChemStation software (Agilent Technologies). Data were expressed as absolute concentration (µg/ml plasma), which is related to fatty acid consumption28. Long-chain n-3 PUFA were considered 20:5n-3 (EPA), docosapentaenoic acid (DPA; 22:5n-3), and 22:6n-3 (DHA).
Activities of desaturase enzymes, and in particular Δ6 desaturase (D6D), are considered the rate-limiting steps in the conversion of the precursor n-3 PUFA ALA to the longer chain EPA and DHA, as well as in the parallel metabolism of n-6 PUFA. Activity of desaturase enzymes can be inferred from product to precursor ratios29. These ratios were used to estimate the activity of the Δ5 (20:4n-6/20:3n-6) and Δ6 (18:3n-6/18:2n-6) desaturases.
Th17 cell differentiation
Human CD4+CD25− T cells and autologous monocytes were isolated from peripheral blood mononuclear cells obtained from fresh leukocyte reduction system cones (National Blood Service) by ficoll gradient centrifugation. Negative selection reagents (StemCell Technologies) were used for the enrichment of monocytes and CD4+CD25− T cells, and greater than 95% cell purity was attained as confirmed by flow cytometry using antibodies against CD3, CD4, and CD14 (BD). CD25 expression on T cells could not be assessed because of the interference of the anti-CD25-TAC reagent used for the cell selection. Efficient depletion of regulatory T cells was therefore determined by staining for FoxP3 and CD127.
To assess Th17 differentiation, T cells and monocytes were cultured at a ratio of 1 monocyte to 5 T cells in the presence of anti-CD3 (OKT3 clone; 0.5 μg/ml). The cultures were prepared in RPMI 1640 medium supplemented with 50 U/ml penicillin and streptomycin and 2 mM L-Glutamine (Gibco, Life Technologies) and incubated at 37°C, 95% humidity, and 5% CO2. After 7 days, T cell expression of interleukin 17 (IL-17) and interferon-γ (IFN-γ) was assessed by intracellular cytokine staining and flow cytometry. In brief, T cells were restimulated for 5 h with phorbol myristate acetate (50 ng/ml) and ionomycin (1 μM). Brefeldin A (10 μg/ml) was added during the final 4 h to promote cytokine accumulation. Cells were washed with phosphate buffered saline (PBS) and dead cells labeled with a fixable live/dead discrimination dye (Life Technologies) before fixation for 12 min in 3% paraformaldehyde. Following 1 wash with PBS, cells were permeabilized with 0.1% saponin-PBS and labeled with antibodies against CD3 (BD), IL-17 (eBiosciences), and IFN-γ (BD). To assess the effects of ETN, EPA, and the control n-6 fatty acid [linoleic acid (LA)] on Th17 differentiation, the cultures were also supplemented with EPA or LA at 5 μg/ml, with or without ETN.
Statistical analysis
Comparison of baseline data between subjects randomized to IFX versus ETN used the Mann-Whitney U test or the independent samples Student t test depending on distribution of data. Subsequent analyses pooled both groups for a population size of 22. Comparison of fatty acid levels and product/precursor ratios between baseline and Week 12 were undertaken using the Wilcoxon signed-rank test. Correlations of plasma fatty acid levels at baseline, Week 12, and the mean of these timepoints (mean value considered the primary dependent variable of interest) with change in DAS28 were analyzed using Spearman ρ. The relationship of European League Against Rheumatism (EULAR) outcomes to EPA levels, and of EPA tertiles with change in DAS28 and its constituents, was tested by 1-way ANOVA. Analyses of the clinical and in vitro data were undertaken using SPSS 20 and Graphpad Prism 5, respectively.
RESULTS
Baseline and 12 weeks
Baseline demographics are shown in Table 1 and the 2 treatment groups were not statistically different. To determine the effect of anti-TNF on desaturase activity, we compared fatty acid profiles from baseline and Week 12 of treatment, with a view to using mean values across timepoints to give a more robust biomarker of PUFA status over the time course of the study. Interestingly, there was a trend toward a reduction in the 18:3n-6/18:2n-6 ratio following treatment with anti-TNF (p = 0.05), which might indicate a reduction in D6D activity after 12 weeks, being accompanied by a lower median 18:3n-6 product (p = 0.08), 0.60 μg/ml [interquartile range (IQR) 0.52–1.15] versus 0.51 μg/ml (IQR 0.47–0.62), and a higher level of 22:4n-6 (adrenic acid; p = 0.039), 0.15 μg/ml (IQR 0.11–0.21) versus 0.19 μg/ml (IQR 0.12–0.27). Adrenic acid was the last measurable n-6 PUFA prior to the second Δ6 desaturase catalyzed step. However, no significant differences were observed in any other fatty acid after anti-TNF treatment, including all the longer chain n-3 fatty acids (data not shown), and therefore subsequent analyses used the average of baseline and Week 12 measurements.
Relationship of plasma phosphatidylcholine fatty acids with change in DAS28
Table 2 shows the correlations between individual fatty acids, total n-3, n-3/n-6, and EPA/AA ratios, and desaturase product/precursor ratios with change in DAS28 after 3 months of anti-TNF therapy. The most significant correlations with time-averaged levels were seen with EPA (−0.51, p = 0.007) and the EPA/AA ratio (20:4n-6; −0.48, p = 0.01), indicating that higher plasma levels of EPA were associated with a greater reduction in DAS28 at 3 months after anti-TNF. This association with EPA was still statistically significant when restricting the analysis to subjects treated with ETN (r = −0.54, p = 0.04), with a trend seen in the IFX group (r = −0.42, p = 0.10).
Correlations (Spearman ϱ) between plasma phosphatidylcholine n-3 and n-6 fatty acids (μg/ml) at baseline, Week 12, and the averaged concentration from both timepoints, with change in DAS28 score following 12 weeks of anti-TNF therapy. Long-chain n-3 PUFA were considered 20:5n-3 (EPA), 22:5n-3 (DPA), and 22:6n-3 (DHA). Product to precursor ratios were used to estimate the activity of the Δ5 (20:4n-6/20:3n-6) and Δ6 (18:3n-6/18:2n-6) desaturases.
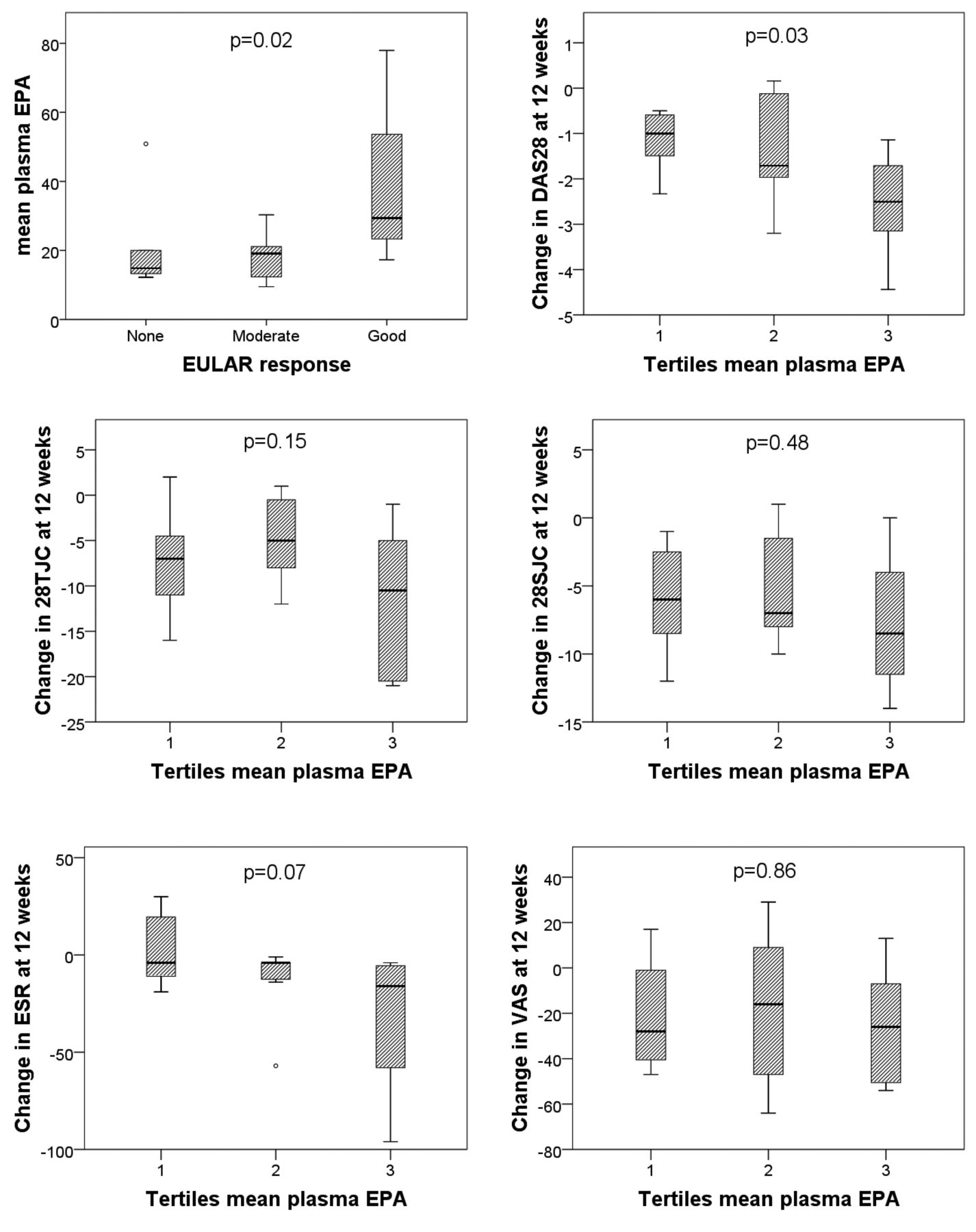
Relationship of plasma phosphatidylcholine EPA levels with EULAR response and components of DAS28
At 12 weeks, 6 subjects were EULAR nonresponders30, and 13 and 3 patients were moderate and good responders, respectively. Median EPA levels in these groups were 14.8 µg/ml, 19.1 µg/ml, and 29.4 µg/ml, respectively (p = 0.02; Figure 1). EPA levels were next divided into tertiles and the highest tertile was associated with a larger fall in DAS28 scores at 12 weeks (p = 0.03; Figure 1). Figure 1 also shows the 4 components that contain the DAS28 score, in relation to tertiles of EPA, with trends being observed for erythrocyte sedimentation rate (p = 0.07) and TJC (p = 0.15).

Time-averaged plasma EPA levels in patients who were EULAR nonresponders, moderate responders, and good responders at 3 months. Relationships between tertiles of time-averaged plasma EPA, and change in Disease Activity Score at 28 joints (DAS28) and its 4 constituents (28TJC, 28SJC, patient’s VAS for global disease activity, and ESR), and Health Assessment Questionnaire at 3 months are also shown. EPA: eicosapentaenoic acid; EULAR: European League Against Rheumatism; 28TJC: tender joint count of 28 joints; 28SJC: swollen joint count of 28 joints; VAS: visual analog scale; ESR: erythrocyte sedimentation rate.
Effect of ETN and EPA on Th17 differentiation
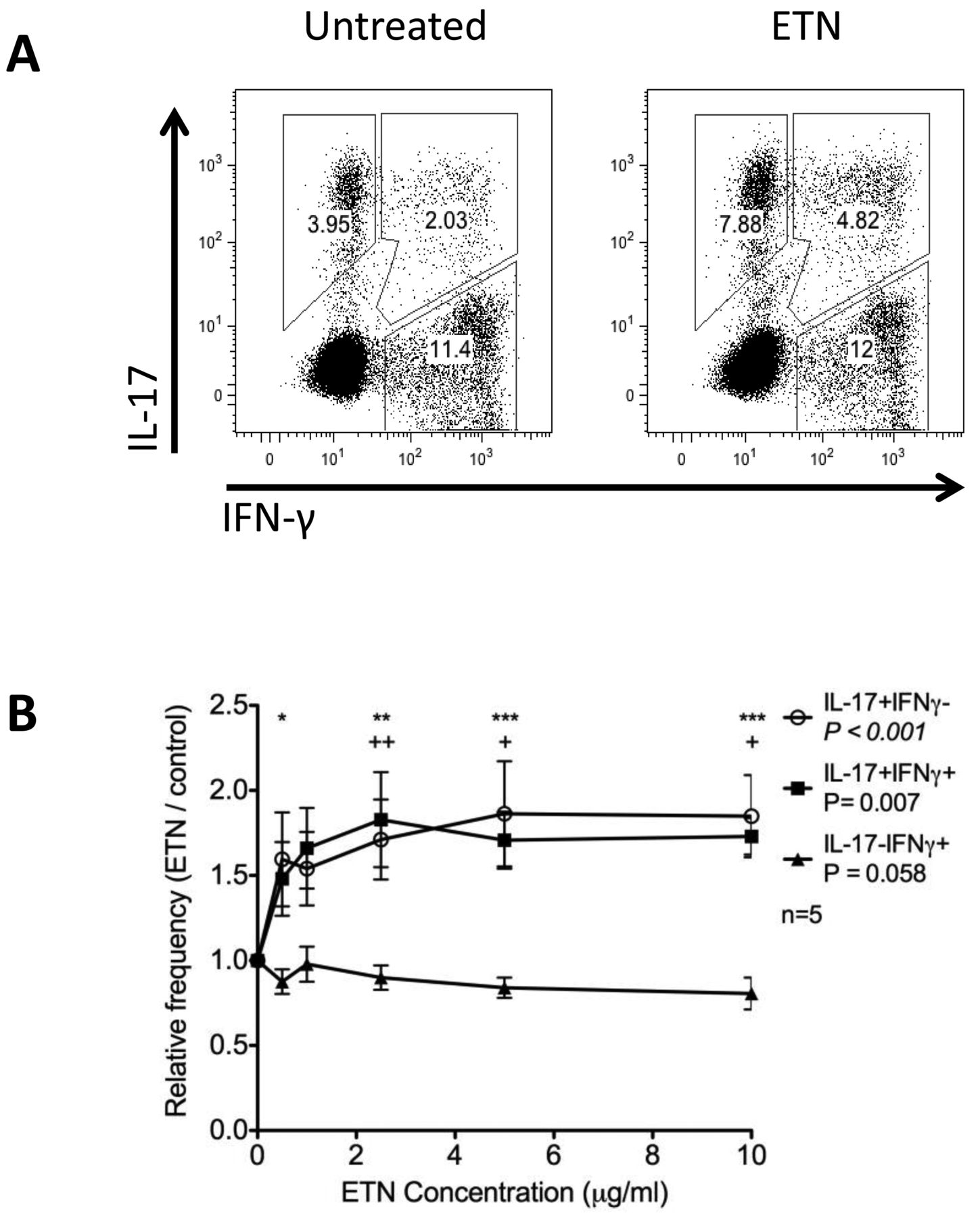
Poor responses to anti-TNF therapy have been associated with increased IL-17 production and the frequency of Th17 cells3,31. Therefore, using our established monocyte-driven T cell stimulation system to study Th17 differentiation in vitro32, we investigated the effect of anti-TNF on Th17 differentiation. At pharmacologically relevant concentrations, ETN promoted a significant, 1.6-fold increase in the frequency of cells expressing IL-17 (Figure 2A and Figure 2B). This increase in frequency included cells that expressed IL-17 alone, as well as those that expressed IL-17 together with IFN-γ. By contrast, the frequency of cells that expressed IFN-γ alone was not affected (Figure 2B). Treatment with IFX did not significantly increase Th17 frequencies in this assay (data not shown). Given that elevated levels of EPA were associated with improved responses to anti-TNF, we were interested to see whether this effect could involve downregulation of IL-17 by EPA. Thus, we repeated the monocyte-driven T cell stimulations in the presence of EPA or the control n-6 fatty acid, LA. Neither EPA nor LA affected the frequency of Th17 cells relative to ethanol carrier control. Importantly, EPA but not LA prevented the ETN-induced increase in Th17 frequency (Figure 3).

ETN increases the frequency of IL-17+ T cells. CD4+CD25− T cells were stimulated with autologous monocytes for 7 days under increasing concentrations of ETN, and the frequency of cells expressing IL-17 and IFN-γ measured by flow cytometry. Representative data for control and 2.5 μg/ml ETN are shown in panel A and relative frequencies of IL-17+ and IFNγ+ cells across ETN concentrations 0–10 μg/ml are summarized in panel B for 5 donors. Significance was tested by ANOVA with Bonferroni posthoc analysis. Asterisks and plus signs indicate significance regarding minus ETN. Asterisks are used for IL-17+IFNy− and plus signs for IL-17+IFNy+ cells. * p < 0.05. ** p < 0.01. *** p < 0.001. ETN: etanercept; IL-17: interleukin 17; IFN-γ: interferon-γ.

EPA inhibits induction of IL-17+ T cells by etanercept (ETN). CD4+CD25− T cells were stimulated with autologous monocytes for 7 days in the presence of ethanol (carrier control), 5 μg/ml LA (control fatty acid), or 5 μg/ml EPA with or without 2.5 μg/ml ETN. The frequency of IL-17+ T cells was measured by flow cytometry. Bars show the mean frequencies for 7 donors. Error bars indicate SD. Significance was tested using paired T tests. EPA: eicosapentaenoic acid; IL-17: interleukin 17; LA: linoleic acid; ns: not significant; EtOH: ethanol.
DISCUSSION
In this cohort of patients with RA receiving either ETN or IFX, higher plasma levels of EPA were associated with a greater reduction in DAS28 scores following treatment compared with patients with lower plasma EPA. These differences were more significant in patients receiving ETN, and it is possible that differences in construct, avidity, and immunogenicity may influence these findings33. Our observed association may relate to the modest benefits previously seen with n-3 PUFA supplementation in patients with RA, although a key difference is that there was no n-3 PUFA supplementation in our study13,14,15. We therefore hypothesized that higher levels of EPA may have additional mechanisms relevant to anti-TNF treatment.
One potentially detrimental effect of anti-TNF therapy is a variable increase in Th17 cells. This is thought to result from the reversal of TNF-mediated p40 suppression, p40 being a subunit of IL-23, which is important in Th17 cell differentiation34. Higher production of IL-17 has been associated with nonresponse to anti-TNF3,31, and there is currently great interest in dual targeting of TNF and IL-17 to optimize biological responses in the face of cytokine compensation35. Were EPA to suppress Th17 differentiation in the context of anti-TNF therapy, supplementation with fish oil might present an alternative, and potentially safer, combination approach.
We were able to replicate this anti-TNF–mediated effect on Th17 frequencies in vitro, using a coculture assay of CD4+CD25− T cells stimulated with autologous monocytes. Addition of ETN, a TNF-receptor fusion protein, resulted in increased frequencies of IL-17+ cells in a dose-dependent manner. Addition of EPA, but not the n-6 LA control, prevented this ETN-driven increase in Th17 frequency.
The mechanism behind this observation has not been established, but eicosanoid metabolites of the n-6 fatty acid AA have also been associated with promotion of Th17 generation. Prostaglandin E2 (PGE2) stimulates IL-23 and IL-1β production by macrophages and dendritic cells while inhibiting IL-12, and it also increases the expression of IL-23 and IL-1β receptors on T cells, so regulating Th17 differentiation36. EPA competes with AA for the prostaglandin-generating COX enzymes, has an inhibitory effect on this enzyme, and yields the generally less proinflammatory PGE337. If alteration of eicosanoid metabolites does influence response to anti-TNF therapy, then one might also expect synergism between anti-TNF drugs and NSAID. Notably, some registry studies have indeed found the use of NSAID to predict better response to anti-TNF38,39, although this association was not considered at the time to be causal, and other studies have not confirmed this40. Against this eicosanoid hypothesis might be the observation that inhibition of PGE2 production by monocytes ex vivo required a relatively high intake of EPA in a dose-response study, although this was conducted in healthy volunteers rather than patients with RA41.
Other mechanisms may also be important. In mouse models of colitis, n-3 fatty acids have been demonstrated to reduce Th17 cell numbers42,43, thought in part to reflect reduced membrane-raft responsiveness to the Th17-polarizing cytokine IL-644. Indeed, the ability of MTX to reduce IL-6 levels may in part explain the benefits of combining this drug with anti-TNF45.
Plasma phosphatidylcholine fatty acids are a better indicator of dietary intake in comparison with dietary questionnaires because of the measurement error inherent in the latter46, but plasma phosphatidylcholine fatty acids reflect only dietary intake over the preceding few days or weeks47,48,49,50. We therefore used the average of baseline and Week 12 measurements to obtain a more reliable biomarker of PUFA status. It should be noted that an individual’s long-chain n-3 PUFA status may be influenced not just by dietary n-3 intake, but also by the amount of n-6 PUFA in the diet because these compete for the same desaturase and elongase enzymes, and by polymorphisms in the desaturase enzymes29. Although our primary analysis focused on time-averaged plasma fatty acids, it is of interest that Week 12 levels of DPA were also negatively correlated with DAS28 levels. The effect of DPA on inflammation is relatively underinvestigated compared with EPA and DHA, but a study of complete Freund’s adjuvant-induced arthritis in rats found that monoglyceride EPA or DPA, but not DHA, reduced paw swelling, arthritis score, and levels of proinflammatory cytokines51. Further investigation of DPA antiinflammatory effects would be warranted.
D6D is considered a rate-limiting step in long-chain PUFA metabolism. The transcription and therefore activity of D6D may be upregulated by insulin and statins52 and downregulated by long-chain PUFA53, glucagon, adrenaline, steroids, and smoking54,55. Interestingly, there is a recognized association of D6D activity and later development of Type 2 diabetes, independent of disturbances in glucose metabolism56. The reason for this is not clear, but inflammation is a risk factor for insulin resistance57 and our data imply that inflammation may also increase D6D activity, providing a possible explanation for this association. Our findings are consistent with the observation that TNF-α increases hepatic expression of sterol regulatory element-binding protein-1c in mice58, because SRBP-1c is involved in upregulating D6D gene transcription53. The alteration in D6D activity we observed does not explain the association of anti-TNF response with EPA levels, for several reasons: such an association would predict an opposite relationship, baseline levels of EPA are also predictive of response, and D6D activity plays only a small role in the conversion of longer chain n-3 fatty acids9,56, as confirmed by the comparison of fatty acid levels pre- and post-anti-TNF. The desaturase activity inferred from these fatty acid ratios generally reflects conversion in the liver4; however, monocytes also demonstrate D6D and D5D activity55,59,60, and we cannot rule out the possibility that an alteration of desaturase activity may be more pronounced in these cells, which rely on AA, a long-chain n-6 PUFA, for eicosanoid production following stimulation.
There are some limitations in our study. The sample size is small and therefore the study requires replication. In view of the sample size and the pilot design of these data, we did not apply a Bonferroni correction. We did not measure the fatty acid profile in mononuclear cell membranes, which arguably may be more physiologically relevant, although these correlate well with plasma levels61. Finally, a causal relationship cannot be assumed from this data. The strengths are a well-characterized population with a biomarker of fatty acid intake at more than 1 timepoint.
We have presented an association of EPA levels with anti-TNF response in RA. If replicated in further studies and causality confirmed, dietary modulation may provide a simple method for improving outcomes with this therapy.
Footnotes
An unrestricted grant from Schering-Plough (now Merck) supported the clinical trial upon which this study is based.
- Accepted for publication December 21, 2016.








{kind=link}
{kind=link}
{kind=link}