

Dis-moi ce que tu manges, je te dirai ce que tu es. [Tell me what you eat and I will tell you what you are.]
Jean Anthelme Brillat-Savarin, 1826
The so-called paleolithic diet has become popular in recent years among some diet enthusiasts. While this diet may soon become one of many passing fads, it is nevertheless true that our human ancestors consumed a diet that was very different from that of today1. The paleolithic diet had about as much cholesterol as is consumed by modern humans, but also contained far more polyunsaturated fat derived from wild game. Our hunter-gatherer ancestors survived by consuming wild game as their primary source of protein. Fat of wild animals contains about 9% eicosapentaenoic acid (EPA) and 5 times more polyunsaturated fat per gram than is found in domesticated livestock2, which contains almost undetectable amounts of omega-3 (also known as n-3) fatty acids (FA). The current FA intake must be properly viewed as a more recent alteration of longstanding dietary patterns to which humans had adapted during tens of thousands of years1,3 (Figure 1).

There are good sources of n-3 FA in certain leafy plants and vegetables as well as flax, canola, soybean, evening primrose, and borage seed oil supplements; walnuts are another good source. These terrestrial n-3 sources contain α-linolenic acid (ALA; 18:3,n-3). The 18 represents the number of carbon atoms in the molecule; the 3 immediately after the colon stands for the number of double bonds; the n-3 means that the first double bond is found 3 carbon atoms removed from the terminal methyl group of the FA molecule. The structure of various dietary long-chain FA is depicted in Figure 21,3. The food chain leading to human consumption of these FA is seen in Figure 3.
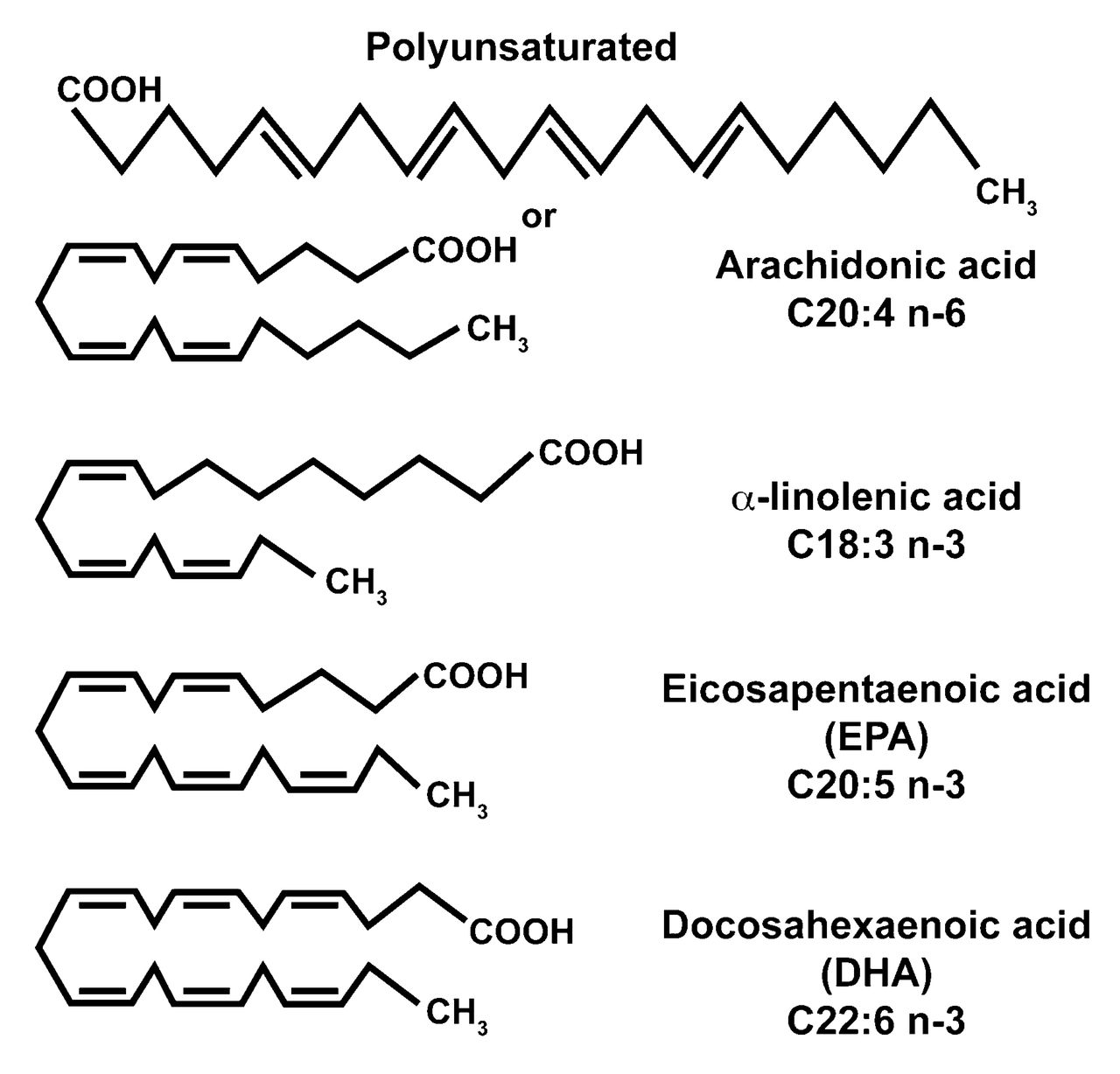
Nomenclature and structure of long-chain dietary fatty acids. The Omega (n-3) status is derived from the position of the first double bond from the terminal methyl group.
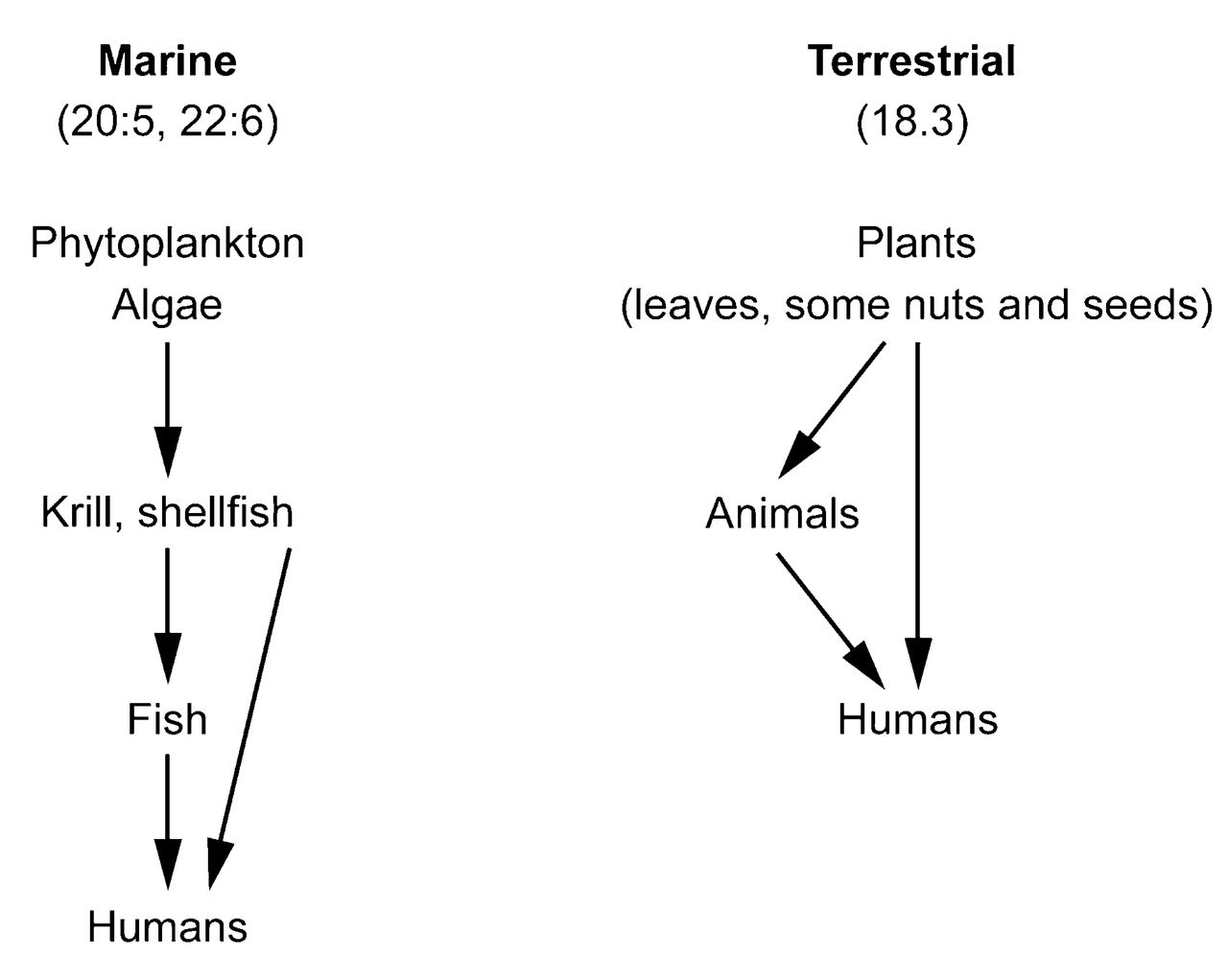
Marine and terrestrial sources of n-3 fatty acids in the food chain to the point of human consumption.
However, as described by Jeffery and colleagues in this issue of The Journal4, it is only the longer chain FA 20:5,n-3 (EPA) and 20:6,n-3 [docosahexaenoic acid, or DHA, and to a lesser extent 22:5,n-3 (docosapentaenoic acid)] that are biologically active. These highly polyunsaturated molecules can significantly alter eicosanoid metabolism by competing with arachidonate (20:4,n-6) for the production of prostaglandins and leukotrienes, resulting in eicosanoid mediators with significantly less inflammatory potential. In addition, dietary supplementation of EPA and DHA is associated with significant alterations in certain cytokines, including interleukin 1 (IL-1)5, also affecting production of other inflammatory cytokines6.
The conversion of ALA (18:3,n-3) to the longer chain EPA and DHA molecules occurs through the Δ-6 desaturase and elongase enzymes, which are inefficient in humans. Thus, unless one ingests the supplements described above, or has a terrestrial diet unusually rich in 18:3,n-3, the biologically active highly polyunsaturated n-3 FA EPA and DHA will be primarily from marine sources. Fatty fish from cold waters such as salmon, tuna, and mackerel are particularly rich in these biologically active n-3 FA. Besides marine sources in the diet there are also an abundance of fish oil (FO) supplements in retail and online nutrition and pharmacy outlets. These supplements are commonly consumed by individuals for general health reasons and have been shown to be effective in a variety of autoimmune diseases7.
Although still controversial, it is interesting to once again consider that rheumatoid arthritis (RA) itself may be a disease of relative modernity. The dropoff in naturally occurring n-3 in the modern diet depicted in Figure 1 may be purely coincidental, of course, and does not prove a connection between diet and RA susceptibility. Nevertheless, it is tempting to consider a link.
Early studies on the effect of dietary supplements of EPA and DHA in NZBxNZW F1 mice showed a significant amelioration of proteinuria and renal failure when compared to mice consuming a diet with beef tallow8 as well as amelioration of collagen arthritis9. These animal studies were soon followed by the first studies of dietary FO supplementation in patients with active RA10. There have been about 20 published trials of FO supplementation in RA and most are at least 20–30 years old10,11,12,13,14,15. These studies documented common alterations in eicosanoid and IL-1 production11,12,13 associated with clinical improvements in tender joints in patients with the disease. These studies were all conducted in patients with RA receiving established conventional disease-modifying antirheumatic drug and nonsteroidal antiinflammatory drug treatments, rather than as substitutes for these medications. All of these earlier trials of n-3 dietary supplementation were conducted in the prebiologic era. More recently, it has been demonstrated that patients with early RA do better with supplementation of n-3 FA16.
It is therefore of some particular note that Jeffery and colleagues4 have observed a relationship between plasma levels of n-3 FA and response to tumor necrosis factor (TNF) inhibitors, which are linked to production of IL-173. Building on earlier reported effects of TNF inhibition on the IL-17 pathway17, the authors have observed that increased levels of plasma n-3 FA were associated with decreased expression of IL-17, resulting in a consequent enhanced clinical response to TNF inhibition. It is interesting that the patients in this small trial were not described as consuming a diet high in n-3 and were not given supplements containing these FA.
If non-supplemented patients with RA have diminished enhancement of IL-17 production that is inversely related to their plasma n-3 levels (the greater the plasma n-3, the less IL-17 is produced), what additional benefit might be derived from taking daily FO supplements in the ranges previously reported10,11,12,13,14,15?
Indeed, if these observations can be replicated, it is possible that routine supplementation of FO given with TNF inhibitors could ameliorate some of the poor responses sometimes seen with this class of biologic agents. As noted, the authors place their observation in the context of previously established inhibitory mechanisms of TNF itself on the production of IL-1717. Thus inhibition of TNF would be expected to enhance IL-17 production with the potential to abrogate the beneficial clinical effects of TNF inhibition.
Although the number of patients reported by Jeffery and colleagues is quite small, their observations are fascinating and potentially relevant for improving TNF responses. We really do not entirely understand the reasons why some patients fail TNF inhibition, although immunogenicity and neutralizing antibodies to monoclonal TNF inhibitors have been well described18, along with a variety of more loosely linked genetic traits19.
Prospective investigations in large cohorts of the effects of FO supplementation in subjects receiving TNF inhibitors for the first time should be conducted with measurement of IL-17 production in plasma. The studies can simultaneously examine the correlation of clinical response with the level of n-3 FA in plasma, and membrane phospholipids. The hypothetical investigations could seek to determine whether strong correlations are found between n-3 levels and diminution of IL-17 production compared to a control group receiving n-6 FA (such as corn oil) while being treated with TNF inhibitors. If the subjects with RA receiving n-3 supplements do indeed have superior clinical responses compared with matched controls, then the small study by Jeffery will have led the way to a probable explanation of why some patients fail TNF inhibitor drugs, while providing a mechanism to overcome the problem.








{kind=link}
{kind=link}
{kind=link}