

Abstract
Objective. To identify shared and differential molecular pathways in blood and affected muscle between adult dermatomyositis (DM) and juvenile DM, and their association with clinical disease activity measures.
Methods. Gene expression of transcription factors and cytokines involved in differentiation and effector function of T cell subsets, regulatory T cells and follicular Th cells, were analyzed in the blood from 21 newly diagnosed adult and 26 juvenile DM subjects and in 15 muscle specimens (7 adult and 8 juvenile DM) using a custom RT2 Profiler PCR Array. Disease activity was determined and measured by established disease activity tools.
Results. The most prominent finding was the higher blood expression of Th17-related cytokines [retinoic acid-related orphan receptor-γ, interferon regulatory factor 4, interleukin (IL)-23A, IL-6, IL-17F, and IL-21] in juvenile DM at baseline. In contrast, adult patients with DM showed increased blood levels of STAT3 and BCL6 compared with juvenile DM. In muscle, GATA3, IL-13, and STAT5B were found at higher levels in juvenile patients with DM compared with adult DM. Among 25 patients (11 adult and 14 juvenile DM) who had blood samples at baseline and at 6 months, increased expression of IL-1β, STAT3, STAT6, STAT5B, and BCL6 was associated with an improvement in global extramuscular disease activity.
Conclusion. We observed differences in gene expression profiling in blood and muscle between new-onset adult and juvenile DM. Cytokine expression in the blood of juvenile patients with new-onset DM was dominated by Th17-related cytokines compared with adult patients with DM. This may reflect the activation of different Th pathways between muscle and blood.
Dermatomyositis (DM) is a systemic, inflammatory disorder primarily affecting muscle and skin in children and adults. Although the pathogenesis of adult and juvenile DM appear similar, there are important differences in the clinical features and pathophysiology. Juvenile DM has more cutaneous features such as nailfold telangiectasia and calcinosis with a good outcome in the majority of the cases1, whereas adult DM is more resistant to treatment and complete remission is infrequent with a higher mortality rate2,3,4. Similarly, adult patients with DM have a higher risk of developing cancer, but malignancies are rarely reported in juvenile DM5. Interstitial lung disease is a common occurrence in adult DM in up to 23.8% of cases6, but it is uncommon in juvenile DM.
The cellular infiltrates are reported to be similar in adult and juvenile DM and consist of CD4+ T cells, plasmacytoid dendritic cells, B cells, macrophages, and Th17 cells.
Naive CD4+ T cells develop into functionally mature effector cells upon stimulation with relevant antigenic peptides that induce differentiation into at least 4 major Th lineages including Th1, Th2, Th17, and Foxp3+ Treg cells, which produce lineage-indicating cytokines and perform distinct functions in regulating immunity and inflammation7,8,9. In addition, other Th cell subsets have been recently described, such as follicular Th (TFH) cells, Th9 cells, and interleukin (IL) 22-expressing Th22 cells10,11,12.
The main way that differentiated CD4 T cells regulate inflammation is by the release of cytokines. Cytokines and transcription factors are critical for determining CD4+ T cell fates and the effector cytokine production. For instance, IL-12 induces expression of the transcription factors T-bet (TBX21) and signal transducer activator of transcription 4 (STAT4), which mediates the differentiation of Th1 cells with production of interferon-γ (IFN-γ), IL-2, and tumor necrosis factor-α (TNF-α) and -β, whereas IL-4 induces the transcription factors GATA-binding protein 3 (GATA3) and STAT6 with subsequent differentiation of Th2 cells and production of cytokines IL-4, IL-5, IL-13, and IL-1013,14. Th17 cells express the master transcription regulator retinoic acid-related orphan receptor-γ (RORC) and produce canonical IL-17A and IL-17F cytokines. The cytokines IL-6, IL-21, IL-23, and transforming growth factor-β1 (TGF-β1) are crucial factors for the differentiation of Th17 cells8,15. Cytokine production of these Th subsets can be heterogeneous and overlapping, with individual cells within a given polarized Th population at any 1 timepoint not necessarily secreting the full range of cytokine for that subset16.
Studies concerning cytokine expression in muscle tissue of patients with inflammatory myopathies including DM have shown the dominant presence of IL-1α, TGF-β1, IL-6, and proinflammatory Th1 cytokines (IFN-γ, IL-2), and a weak expression of Th2-derived cytokines17,18,19,20, suggesting a Th1/Th2 balance in muscle tissue with a greater Th1 response in DM. Interestingly, a study showed that CD4+IFN-γ+ cells were decreased whereas the CD4+IL-4+ cells (Th2 cells) were increased in the blood of patients with new-onset DM21, suggesting that Th2 cells may predominate in the blood of active DM. In parallel to Th1 and Th2 cells, studies have begun to elucidate the importance of IL-17–producing Th17 cells in the pathogenesis of myositis18,22,23.
Although it appears that the pathogenesis of DM in adults and children is similar, striking differences in clinical features, outcome, and response to treatment exist and suggest that different mechanisms may be at least partially involved. Only a few studies have previously compared the immunopathological features of adult and juvenile DM; most studies combine these diseases. In our study, we sought to compare mRNA expression differences between adults and children with new-onset DM in genes related to Th cell signaling pathways and/or innate immune response using gene expression profiles obtained from peripheral blood mononuclear cells (PBMC) and affected muscle tissues. We also sought to determine whether changes existed in gene expression profiles between baseline visit and 6 months of followup in both DM groups and how they related to changes in clinical disease activity. The purpose of our study was to extend our previous findings24 to include both gene expression data obtained from blood and affected muscle samples to assess which “pathways” or individual genes were deregulated in juvenile and adult DM.
MATERIALS AND METHODS
Blood samples and clinical data were obtained prospectively from 26 children and 21 adults with new-onset disease (< 6 mos of clinical symptoms) who fulfilled the Bohan and Peter criteria25 for the diagnosis of DM at baseline. Blood samples were available in 25 patients (11 adults and 14 juveniles) at baseline and 6 months. Of the 47 patients, 8 were included in a previous report24 and 39 were new samples. All subjects had a definitive diagnosis of DM and were seen at the Division of Rheumatology at the Mayo Clinic, Rochester, Minnesota. The Mayo Clinic Institutional Review Board approved this study (10-005501), and informed consent was obtained from each participant. Disease activity measures included the extramuscular disease activity, physician’s global activity (PGA), and the manual muscle testing of 8 muscle groups24. PBMC were collected at baseline and at a 6-month followup visit. In addition, paired muscle biopsies were obtained from 7 adults and 8 juvenile patients with DM at baseline visit. Biopsies were obtained from the vastus lateralis in 7 patients (all juvenile DM) and from the deltoid and triceps muscles for 8 (7 adult DM and 1 juvenile DM). These specimens were reviewed and confirmed to be DM by a neuromuscular pathologist (neuromuscular laboratory, Mayo Clinic, Rochester, Minnesota, USA).
Gene expression profiling
Whole blood was collected in tubes treated with an RNA stabilization agent (PAXgene; PreAnalytiX). Total RNA was isolated according to the manufacturer’s protocol with on-column DNase treatment. RNA yield and integrity were assessed using an Agilent Lab-on-a-chip Bioanalyzer (Agilent Technologies Inc.). Muscle was homogenized using a PowerGen 700 (Fisher Scientific) and RNA isolated by an organic extraction method (TRIzol; Life Technologies), and was further purified with RNeasy Mini Kit (Qiagen) and an on-column DNase treatment. We measured mRNA expression of IL-1β, IL-6, IL-23A, IL-27, IL-17A, IL-17D, IL-17F, IL-21, IL-22, RORC, TGF-β1, IFN-γ, TNF-α, IL-2, TBX21, STAT4, IL-4, IL-5, IL-10, IL-12B, IL-13, STAT6, GATA3, interferon regulatory factor 4 (IRF4), forkhead box P3 (FOXP3), STAT5B, IL-10, IL-9, B cell CLL/lymphoma 6 (BCL6), STAT3, lymphotoxin-α (LTA/TNFSF1), and the housekeeping genes GAPDH, B2M, and ACTB using a custom RT2 Profiler PCR Array and amplified on an ABI 7900HT PCR system. The genes we analyzed were grouped in “pathways” or functionally related groups of genes according to the particular cytokines secreted by the individual Th cell subset as well as innate immune cells (Table 1). The transcription factors examined were unique to specific Th cell pathways such as TBX21 (Th1), GATA3 (Th2), RORC (Th17), and FOXP3 (Treg). However, we are aware that most genes can act as part of 1 or more pathways. For instance, IRF4 expression, a transcription factor essential for Th2 effector cell differentiation, is also known to interact with STAT3 to induce RORγt expression, suggesting its effect on Th17 cells26.
Functional grouping of genes analyzed in blood and muscle tissue of adult and juvenile patients with dermatomyositis.
Statistical analysis
Data are expressed as the median (range) where indicated. A Wilcoxon rank sum test was performed for comparisons between 2 groups (e.g., adult vs juvenile DM). Paired Student t tests were performed to compare gene expression levels between baseline and 6 months as well as between blood and muscle samples. Spearman correlation coefficients were used to examine correlations between variables. P values < 0.05 were considered statistically significant. Multiple comparisons involved a maximum of 30 comparisons for any specific hypothesis; thus, the Bonferroni correction would consider p values < 0.0017 to be significant. Analyses were performed using SAS version 9.3 (SAS Institute Inc.) and R 3.1.1 (R Foundation for Statistical Computing).
RESULTS
Demographic and clinical characteristics of the patients with DM
Demographic and clinical characteristics of the patients with DM are summarized in Table 2. At baseline, patients in both groups had similar disease activity as evidenced by the values for the disease activity core measures. Ten adult patients with DM (48%) and 6 juvenile patients with DM (29%) had begun treatment with 1 or more traditional disease-modifying antirheumatic drugs (DMARD) in the 2 months prior to the baseline visit. Concomitant corticosteroid use was reported for 14 adults (70%) and 8 children (38%) with DM.
Characteristics at baseline of adult and juvenile patients with DM. Values are mean ± SD unless otherwise specified.
Overall, these patients improved substantially on all disease activity measures between baseline and the 6-month followup visit, with a median improvement of 20/100 in the extramuscular visual analog scale (VAS), 22/100 in the muscle VAS, and 34/100 in the global VAS. Changes in disease activity measures were not significantly different between adults and children with DM (p = 0.14, p = 0.51, and p = 0.07 for PGA, muscle activity, and extramuscular disease activity VAS scores, respectively).
T cell lineage gene expression in blood of new-onset adult and juvenile DM
We investigated whether the T cell lineage gene expression profiling in blood would reveal distinct T cell pathways in juvenile versus adult DM. Gene expression levels of RORC/RORγt, the master regulator for Th17 cells, and IRF4, another transcription factor required for Th17 cell differentiation, were significantly increased in blood of juvenile compared with adult patients with DM at baseline (p = 0.001 and p < 0.001, respectively; Figure 1). In addition, juvenile DM had significantly higher levels of IL-23A (p < 0.001), IL-6 (p < 0.001), IL-17F (p = 0.005), and IL-21 (p = 0.012) mRNA at the baseline visit when compared with adult patients with DM. Similarly, FOXP3 and STAT4 gene levels were significantly increased in the blood of juvenile DM compared with adult DM (p = 0.016 and p = 0.009, respectively). In adult DM blood, increased levels of STAT3 and BCL6 were observed compared with juvenile DM blood (p = 0.028 and p = 0.002, respectively; Figure 1).

Gene expression profile in peripheral blood mononuclear cells of adult and juvenile patients with DM at baseline. The plot depicts the gene expression data from genes that were differentially expressed in blood between juvenile and adult DM. P values < 0.05 were considered statistically significant. Genes expressed as relative quantification. DM: dermatomyositis; JDM: juvenile DM.
T cell lineage gene expression in affected muscle of patients with new-onset DM
As a step toward understanding the differences between juvenile and adult DM, we analyzed gene expression in paired muscle biopsies collected from 15 patients with DM (7 adults and 8 children) at baseline. All DM samples were analyzed histologically to confirm DM. Perivascular lymphocytic infiltration was seen in all DM samples. A gene expression profile was also evaluated in the skeletal muscle tissues from otherwise healthy patients undergoing an orthopedic procedure. The majority of transcripts were found at higher levels in affected muscle tissue of both adult and juvenile DM compared with normal muscle tissues (data not shown).
Evidence of Th2 T cells in muscle tissue was demonstrated by increased expression levels of GATA3 (p = 0.004) and IL-13 (p = 0.049), and Treg cells by increased STAT5B expression (p = 0.049) in affected muscle of juvenile compared with adult patients with DM.
Correlation of T cell lineage gene expression with medication use in patients with DM
Glucocorticoids are critical regulators of several inflammatory cytokines27,28, but our previous data on cytokine protein levels suggest that glucocorticoid treatment does not statistically alter the levels except for IL-1β24. Herein, we analyzed the association between glucocorticoid treatment and the expression of transcription factors. GATA3, IRF4, and FOXP3 levels were lower in juvenile DM blood from patients who were receiving glucocorticoids versus those not taking glucocorticoids (p = 0.015, p = 0.001, and p < 0.001, respectively). If we looked at just patients (both juvenile and adult DM) who were taking glucocorticoids, IL-23A, IL-6, RORC, IRF4, and BCL6 gene expression levels remained significant when comparing juvenile and adult DM; however, other comparisons that no longer reached statistical significance were marginal: IL-17F (p = 0.056), IL-21 (p = 0.12), STAT4 (p = 0.088), and FOXP3 (p = 0.056), which could be related to the limited sample size. Only STAT3 was clearly no longer significant (p = 0.45).
Correlation of blood gene expression profiles with clinical features in juvenile and adult DM at baseline and at 6-month followup visit
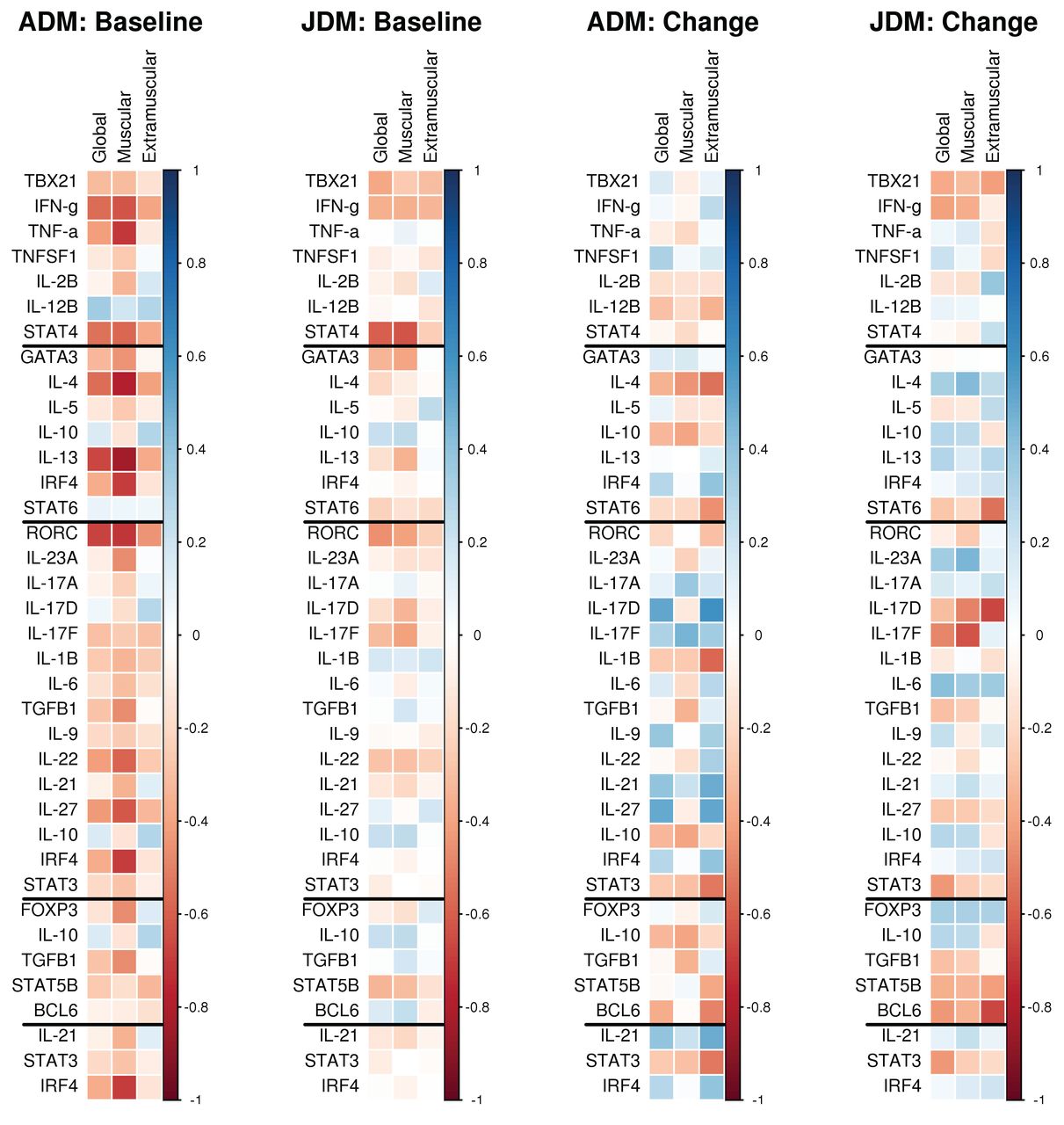
To examine whether differential gene expression was associated with DM disease activity, we compared expression with disease activity at baseline, and examined whether changes in gene expression correlated with changes in the disease activity over time. At baseline in the adults, the gene expression levels of these were found to correlate significantly with muscle disease activity measured at baseline (Figure 2): IL-23A (p = 0.033), RORC (p < 0.001), TGF-β1 (p = 0.035), IL-27 (p = 0.003), IL-22 (p = 0.006), GATA3 (p = 0.045), IRF4 (p < 0.001), IL-4 (p < 0.001), IL-13 (p < 0.001), IFN-γ (p = 0.003), TNF-α (p < 0.001), STAT4 (p = 0.006), and FOXP3 (p = 0.032). Among adults, levels of RORC (p < 0.001), IFN-γ (p = 0.008), STAT4 (p = 0.011), IL-4 (p = 0.009), and IL-13 (p < 0.001) were found to correlate significantly with PGA at baseline and only RORC (p = 0.045) correlated significantly with extramuscular disease activity. In contrast, at baseline in the juvenile DM group, gene expression levels of RORC (p = 0.042), IL-17F (p = 0.040), GATA3 (p = 0.044), and STAT4 (p < 0.001) were found to correlate significantly with muscle disease activity measured at baseline (Figure 2). Only RORC (p = 0.018) and STAT4 (p = 0.001) correlated significantly with global disease activity among the juvenile DM and no significant correlations with extramuscular disease were found.

Correlation of mRNA expressions of T cell lineage transcription factors against measures of disease activity (global, muscular, and extramuscular) cytokines in whole blood of juvenile and adult patients with DM. The 2 leftmost panels correlate baseline gene expression values with baseline disease activity measures. The 2 rightmost panels correlate changes in gene expression from baseline to the 6-month visit with changes in disease activity measures over the same period. Negative correlations between changes in gene expression and changes in disease activity indicate that improvements in disease activity were associated with increases in gene expression levels. ADM: adult dermatomyositis; JDM: juvenile dermatomyositis.
Followup data were available in a total of 25 patients (11 adult and 14 juvenile DM). Among the adults, changes in IL-17D (p = 0.048) between baseline and the 6-month visit were positively correlated with changes in global extramuscular disease activity, indicating that improved disease activity was associated with decreases in gene expression levels (Figure 2). Among the juvenile patients with DM, changes in STAT6 (p = 0.044), IL17-D (p = 0.010), and BCL6 (p = 0.009) correlated negatively with changes in global extramuscular disease activity. No significant correlations between changes in gene expression levels and changes in muscle or PGA measures were found for either group.
Associations between changes in treatment from baseline to 6 months and cytokine gene expression levels were examined longitudinally in the adult and juvenile DM groups combined to enhance statistical power. The addition of DMARD (n = 22) was associated with increased levels of IL-1β (p = 0.012), STAT3 (p = 0.005), STAT6 (p = 0.001), and STAT5B (p = 0.037), and decreased levels of IFN-γ (p = 0.027), IL-22 (p = 0.044), and IRF4 (p = 0.023). Use of glucocorticoids (n = 21) was associated with increases in FOXP3 (p = 0.020), IL-23A (p = 0.013), IRF4 (p = 0.013), and TGF-β1 (p = 0.022), and decreases in IL-2 (p = 0.017). Comparisons of cytokines and transcription factors between blood and muscle did now show significant differences (data not shown).
DISCUSSION
Classically, immunohistochemical and molecular (i.e., real-time PCR) analyses in DM muscle show predominant expression of macrophage-derived cytokines (i.e., IL-1, TNFSF1, and TNF-α), as well as cytokines that can originate from either T cells or macrophages (i.e., IL-6 and TGF-β1)29,30,31. Further, current evidence increasingly supports alternative pathogenic mechanisms, including both adaptive and innate immune-mediated responses. An innate immune response characterized by infiltration of plasmacytoid dendritic cells in DM muscle lesions and IFN-α/β–inducible gene signature in whole blood and muscle may be an important part of the pathogenesis of DM17,32,33,34,35,36.
At present, PCR-based arrays allow facile analysis of expression of a relatively large number of pathway- or disease-focused genes with remarkable sensitivity. Here, we analyzed the expression profile of 30 genes in whole blood from 47 individuals with newly diagnosed DM and in 15 paired muscle samples. To our knowledge, our study constitutes the most comprehensive analysis contrasting the gene expression signatures between adults and children with DM to date.
Cellular and protein effectors of innate and adaptive immunity are found in the blood and inflamed muscle lesions of patients with DM, and an increasing body of evidence indicates that they are directly involved in myofiber injury32,37. We have previously identified a strong correlation between IL-6 serum levels and Type I IFN signature in adult and juvenile DM23. Similarly, a strong expression of IL-1α, IL-1β, TGF-β, and TNF-α has been observed in the affected muscle of patients with DM and other types of myositis, suggesting that these cytokines are important contributors to the pathogenesis of DM19,38,39,40. On the side of the adaptive immune system, despite that muscle-infiltrating T cells are predominantly CD4+ T cells in both adults and children with DM, Th1-derived cytokine IFN-γ and Th2-derived cytokine IL-4 are less abundantly expressed compared with other cytokines30,41, suggesting that other effector T cell lineages might be involved in the pathogenesis or the possible perpetuation of the inflammatory response in DM. Although juvenile and adult DM have a set of common genetic risk alleles and abnormalities in both B and T cell functions, they also have important differences in clinical features and associated disorders, suggesting that specific immunological aberrations may differ for each disease. Our combined analysis of gene expression data in peripheral blood and muscle tissues obtained simultaneously provides a deeper understanding of the T cell pathways that regulate the pathogenesis of both myositis subsets, a finding that eventually could lead to new strategies to facilitate improvement of the therapy.
Herein, we observed differences in gene expression profiling in the blood between juvenile and adult patients with DM. The gene expression profiling that we found in the blood of juvenile patients with DM indicates a Th17 type of inflammatory process. The proportion of patients using glucocorticoids at baseline was higher in adult DM. Despite this difference, after adjusting for glucocorticoid use, we found that IL-23A, IL-6, RORC, and IRF4 gene expression levels remained significantly different in juvenile DM compared with adult DM. The upregulation of Th2-related genes was apparent in the affected muscle of juvenile patients with DM compared with adult DM. However, the interpretation of gene expression profiling in muscle biopsies can be affected by several factors such as a small amount of muscle tissue or the variability of biopsy sampling of inflammatory cells.
Our study also examined the effect of DMARD and glucocorticoid use on the cytokine levels from baseline to 6-month followup. Use of DMARD was associated with increases in blood levels of IL-1B, STAT3, STAT6, and STAT5B, and with decreases in blood levels of IFN-γ, IL-22, and IRF4. Use of glucocorticoids was associated with increases in blood levels of FOXP3, IL-23A, IRF4, and TGF-β1, and with decreases in IL-2, suggesting that various potential immune pathways are altered with treatment.
In addition to the cytokines they produce, effector T cells can be distinguished by their differential expression of specific transcription factors and STAT proteins, which induce gene expression of distinct inflammatory pathways. However, although 1 transcription factor might be the predominant initiator of a differentiation pathway, effector T cells might respond to additional cytokines such as IL-2, IL-6, and IL-21 within the inflammatory environment, which might activate additional T cell lineages. For instance, FOXP3+ Tregs can produce IL-17 when activated in the presence of IL-1β and IL-6, losing their suppressive activity and promoting inflammation42. Additionally, there is much more T cell plasticity than previously recognized wherein a transcription factor may be involved in more than 1 particular differentiation pathway. For example, a study showed that TBX21 (Th1 lineage) and RORC (Th17 lineage) transcription factors were found to be expressed in Th17- and Th1-like Treg cells, respectively43.
In our study, in addition to increased expression of Th17 cytokine genes, RORC and IRF4 (both with critical involvement in Th17 development) were increased in juvenile DM blood compared with adult DM blood. FOXP3 (Tregs) and STAT4 (Th1) gene levels were also elevated in juvenile DM blood.
In contrast, we found higher expression of BCL6 in adult DM blood compared with juvenile DM, which remained significant after adjusting for glucocorticoid use. We observed that STAT3 expression was higher in adult DM blood compared with juvenile DM, although not significant after adjusting for glucocorticoid use. STAT3 is a critical factor for not only activating the proinflammatory Th17 lineage, but for promoting BCL6 transcription and TFH differentiation44. TFH cells are required for germinal center formation, and studies have indicated that TFH is critical in the pathogenic autoantibody production of several autoimmune diseases45,46. BCL6 is upregulated in TFH cells and is considered a master regulator for this lineage. TFH cells have been documented in peripheral blood of juvenile and adult patients with DM47,48. Additionally, we have previously shown that CD4+ T cells and activated B cells come together in ectopic lymphoid aggregates in affected muscle tissue of juvenile patients with DM34, and others have documented similar findings in adult DM49. Together, it seems likely that circulating TFH cells could migrate and local TFH cell-B cell interactions in inflamed muscle tissue may contribute to extranodal immune activation.
In addition to its established role as a transcription factor in TFH differentiation, IL-6–Janus kinase and signal transducer and activator of transcription 3 (JAK-STAT3) activation in muscle cells is strongly associated with skeletal muscle wasting in cancer and other conditions associated with high IL-6 levels50. Cancer and muscle wasting are common features in adult patients with DM; however, whether the IL-6–JAK-STAT3 pathway is a primary mediator of these clinical manifestations in adults with DM needs further investigation.
The differences in gene expression profiling found between juvenile and adult DM can subsequently be applied to gain a better understanding of the complex cellular components involved in the pathogenesis. This can be applied to further studies using emerging technologies such as single cell analysis of circulating immune cell subset and/or molecular analysis of selected cell aggregates in affected muscle tissue using laser identification microdissection. Those analyses could clarify which pathways are shared between juvenile and adult DM, and which differ, identifying particular immune pathways that are active in subsets of patients with DM leading to individualized patient treatment.
Acknowledgment
We thank Jane Jaquith, Heidi Hanf, Jenni Sletten, and Fran Anderson for their efforts in identifying and recruiting patients; and Dr. Andrew G. Engel, Julianna Berge, and Abby Kaehler for their assistance in collecting muscle biopsies.
Footnotes
Supported by the National Institute of Arthritis and Musculoskeletal and Skin Diseases (5R01AR057781-04), the National Center for Advancing Translational Science (NCATS; UL1 TR000135), and the Mayo Foundation for Medical Education and Research.
- Accepted for publication September 30, 2016.








{kind=link}
{kind=link}