

Abstract
Objective. To investigate the effects of the endothelin 1 (ET-1) receptor antagonists (ETRA) macitentan, its active metabolite ACT-132577, and bosentan on myofibroblast activation and extracellular matrix production induced by ET-1 in cultured systemic sclerosis (SSc) and control skin fibroblasts.
Methods. Fibroblasts were obtained from skin biopsies of 6 patients with SSc and 5 healthy subjects. Some cultured cells were untreated or treated with macitentan, ACT-132577, or bosentan alone (10 μM). Other cultured cells were treated with ET-1 alone (100 nM) or with ETRA, and after 1 h, also with ET-1. After 48 h of treatment, myofibroblast activation was investigated to evaluate the α-smooth muscle actin (α-SMA) expression by immunofluorescence; type I collagen (COL-1) and fibronectin (FN) were investigated by immunocytochemistry, Western blotting, and quantitative real-time PCR (qRT-PCR). Statistical analysis was performed by the nonparametric Mann-Whitney U test.
Results. In cultured SSc skin fibroblasts, only the treatment with macitentan significantly reduced the basal level of α-SMA expression (p = 0.03 vs untreated cells). Macitentan also significantly reduced the basal level of COL-1 synthesis, similarly to bosentan (p < 0.05 vs untreated cells). Macitentan or ACT-132577 antagonized the ability of ET-1 to further induce α-SMA expression (p = 0.03), COL-1, and FN synthesis (p = 0.03, p = 0.005); bosentan showed similar effects. These results obtained by immunofluorescence and immunocytochemistry were confirmed by Western blotting and qRT-PCR. The downregulatory effects exerted by ETRA were observed also in cultured human control skin fibroblasts.
Conclusion. Macitentan and ACT-132577 seem to downregulate in vitro the profibrotic myofibroblast phenotype induced by ET-1 in cultured human SSc skin fibroblasts.
- SKIN FIBROSIS
- ENDOTHELIN RECEPTOR ANTAGONISTS
- ENDOTHELIN 1
- SYSTEMIC SCLEROSIS
- EXTRACELLULAR MATRIX
- HUMAN SKIN FIBROBLASTS
Fibrosis is considered to be one of the advanced pathological features of selected chronic autoimmune diseases, including systemic sclerosis (SSc)1. In SSc lesional skin, the progressive tissue fibrosis involves the excessive deposition of elastin fibrils, fibronectin (FN), fibrillar collagens (type I and III), as well as the alteration of enzymes that mediate the posttranslational collagen modifications1. All these events are associated with an increased presence of α-smooth muscle actin (α-SMA)-positive myofibroblasts, which are key mediators of fibrotic tissue remodeling2,3,4,5,6. Therefore, although the pathogenesis of SSc remains unclear, myofibroblast activation has been thought to be the most advanced step following microvascular damage, autoimmune reactivity, and inflammatory reaction3.
Endothelin 1 (ET-1) is considered one of the known mediators of fibrosis, mainly inducing the transition of fibroblasts into myofibroblasts and then increasing the extracellular matrix (ECM) protein production7. Constitutive ET-1 signaling was shown to be necessary to activate the profibrotic gene program induced by transforming growth factor-β1 in both human lung and skin fibroblasts8,9. Because the direct link between ET-1 and its receptors (ETA and ETB) is fundamental to its effects, blocking this interaction with ET receptor antagonists (ETRA) is suggested as a possible strategy to counter the profibrotic process. Both skin fibrosis and the occurrence of new digital ulcers are reduced by treatment with ETRA bosentan in patients with SSc10,11.
Macitentan (ACT-064992) is a new orally active tissue-targeting molecule that inhibits ETA and ETB receptors. Compared to other ETRA, macitentan has a higher tissue penetration and affinity for the lipophilic setting12. In plasma, the circulating metabolite ACT-132577 has been identified as well as macitentan; this pharmacologically active metabolite is the result of the oxidative depropylation of macitentan, and is characterized by lower potency, similar volume of distribution, and a longer half-life compared to its precursor (8.4 h for ACT-132577 vs 2 h for macitentan)13.
Our in vitro study aimed to investigate the effects of the ETRA macitentan and its active metabolite ACT-132577 on both myofibroblast induction and ECM protein production mediated by ET-1 in cultured human skin fibroblasts from patients with SSc and control subjects. Bosentan effects were also tested.
MATERIALS AND METHODS
Patients and healthy subjects
Full-thickness skin biopsies were obtained from the clinically involved skin of one-third of the distal forearm of 6 patients with SSc (4 women and 2 men, mean age 65 ± 7 yrs) recruited from the Division of Rheumatology at the University of Genoa, Italy. Five age-matched healthy subjects (4 women, 1 man) were enrolled from the Division of Dermatology of the same university during routine diagnostic procedures. All patients and healthy subjects gave informed consent, and the study was conducted according to the Declaration of Helsinki and in compliance with the internal ethics board of the Division of Dermatology. The group of patients with SSc fulfilled the new European League Against Rheumatism/American College of Rheumatology criteria for SSc14. No severe clinical SSc complications were present at the time of skin sampling, and patients were treated only with various vasodilators (cyclic prostanoids). Clinical and demographic data of the patients with SSc enrolled into the study are summarized in Supplementary Figure 1 (available online at jrheum.org).
Cell cultures and treatments
Human SSc and control skin fibroblasts were grown in RPMI 1640 medium supplemented with 10% of fetal bovine serum (FBS), 1% penicillin-streptomycin, and L-glutamine (Lonza Clonetic). For the in vitro experiments, human SSc and control skin fibroblasts were used between passages 2 and 5.
Fibroblasts were cultured up to 80% of confluence and then maintained for 18 h in serum-free medium. After serum starvation, some cultured cells were treated with macitentan, or ACT-132577 or bosentan alone (all 10 µm, Actelion Pharmaceuticals). Other cultured fibroblasts were treated as mentioned, and after 1 h, they were also stimulated with ET-1 (100 nM, Enzo Life Science) to evaluate the antagonizing action of ETRA on the profibrotic effects of ET-1 in accordance with previous studies9,15. Moreover, other cultured cells were only treated with ET-1. All treatments were performed in growth medium at 5% of FBS and maintained for 48 h based on the results obtained in our previous in vitro study16. Untreated SSc and control skin fibroblasts, cultured in growth medium at 5% of FBS only, were used as controls.
In a preliminary step of our study, dilution ranges from 1 nM to 10 µm for macitentan, and from 10 nM to 100 µm for ACT-132577, were tested to choose the optimal concentrations able to contrast the ET-1 effects. In particular, the concentrations of 100 nM and 1 μM for both macitentan and ACT-132577 were tested together with the concentration of 10 μM to investigate their contrasting actions on myofibroblast activation and ECM protein synthesis in cultured SSc and control skin fibroblasts in accordance with a recent study17. In these preliminary experiments, concentrations of 100 nM and 1 μM for both macitentan and ACT-132577 did not induce any contrasting effects on the basal levels of α-SMA, type I collagen (COL-1), and FN, or on the increase in these ECM protein syntheses mediated by ET-1 in cultured human SSc and control skin fibroblasts (data not shown).
The concentration of bosentan was established in accordance with previous in vitro studies15,16.
Test using 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
Human control skin fibroblasts were plated into a 96-well tissue culture test plate (2 × 103 cells/well) and maintained for 18 h in serum-free medium. After serum starvation, some cultured cells were treated with ET-1 alone (100 nM) or with different concentrations of macitentan (ranging from 1 nM to 10 µm) either with or without the addition of ET-1 after 1 h. This experimental design was also used to treat other cultured fibroblasts with different concentrations of ACT-132577 (range from 10 nM to 100 µm). All treatments were performed in growth medium at 5% of FBS and maintained for 48 h. Untreated control skin fibroblasts, cultured in growth medium at 5% of FBS only, were used as controls. At the end of treatments, the culture medium was removed and cells were incubated at 37°C, 5% CO2 in presence of MTT labeling reagent (Sigma-Aldrich) for 3 h. Finally, 100 µl of dimethyl sulfoxide were added to each well18. The absorbance was measured at 490/620 nm. Five independent experiments were performed in triplicate.
Immunofluorescence and immunocytochemistry
SSc and control skin fibroblasts were cultured in Flexi PERM chamber slides (5 × 103 cells/spot, Millipore) and treated as described in the “Cell cultures and treatments” paragraph. At the end of treatment, cells were fixed in methanol and incubated with primary antibody to human α-SMA (dilution 1:100, Dako Cytomation), a marker of the myofibroblast phenotype7. FITC-immunoglobulin G (IgG) secondary antibody (dilution 1:200, Cell Signaling Technology) was used to detect the generated fluorescence. Nuclei were counterstained with 4′,6-diamidin-2-phenylindole (DAPI, Sigma-Aldrich).
Immunocytochemistry was performed by incubating the cells with primary antibodies to human COL-1 (dilution 1:100, Abcam) and FN (dilution 1:100, Sigma-Aldrich). Linked antibodies were detected by biotinylated universal secondary antibody and subsequently with a horseradish peroxidase (HRP)-streptavidine complex (Vector Laboratories). The analysis of α-SMA expression and ECM protein synthesis were performed in each experimental condition, evaluating the same number of cells by light microscopy (magnification 20×, Leica). The subsequent computerized image analysis was performed by Leica Q500MC Image Analysis System (Leica). The values of α-SMA expression obtained from each treatment were normalized to that of the untreated cells (taken as unit value by definition) and indicated as a level of α-SMA expression. The values of COL-1 and FN synthesis obtained from each experimental condition were indicated as levels of protein synthesis. The results of immunofluorescence and immunocytochemistry were obtained from 6 independent experiments on cultured SSc skin fibroblasts and 5 independent experiments on cultured human control skin fibroblasts.
Western blotting
SSc and control skin fibroblasts were cultured up to 80% of confluence and treated as previously described. At the end of treatments, cells were lysed with NucleoSpin RNA/protein (Macherey-Nagel). The protein quantification was performed by the Bradford method. For every condition, 40 µg of protein were separated by electrophoresis on 8% Tris-Glycine gel and transferred onto Hybond-C-nitrocellulose membrane (Life Technologies Ltd.).
After 1 h in blocking solution (PBS1×, 0.1% triton-X, and 5% nonfat powdered milk), membranes were incubated overnight at 4°C with primary antibodies anti-human COL-1 (dilution 1:500, Enzo Life Science) and anti-human FN (dilution 1:1000, Sigma-Aldrich). Membranes were also incubated with primary HRP-conjugated antibody to human-actin (dilution 1:10,000, Santa-Cruz Biotechnology) to confirm similar loading of gels and efficiency in the electrophoretic transfer.
Membranes were subsequently incubated with secondary antibodies antirabbit IgG for COL-1 (dilution 1:2000, Cell Signaling Technology) and antimouse IgG for FN (dilution 1:5000, GE Healthcare Europe). Protein synthesis was detected using the enhanced chemiluminescence system (Luminata Crescendo). Densitometric analysis was performed by Leica Q500MC Image Analysis System (Leica).
For each experimental condition, the values of COL-1 and FN synthesis were normalized to those of the corresponding actin. The resulting values of each treatment were compared to that of the untreated cells (taken as unit value by definition) to obtain the level of protein synthesis. Western blotting was performed on 5 independent experiments on cultured human skin fibroblasts from patients with SSc and 4 independent experiments on cultured human skin fibroblasts from healthy subjects.
Quantitative real-time PCR (qRT-PCR)
SSc skin fibroblasts were cultured up to 80% of confluence and treated as previously described. Total RNA was extracted with NucleoSpin RNA/protein (Macherey-Nagel) and quantified by NanoDrop (Thermo Scientific), which also evaluates RNA integrity, in accordance with the manufacturer’s protocol. For each experimental condition, first-strand cDNA was synthesized from 1 µg of total RNA using QuantiTect Reverse Transcription Kit (Qiagen).
The qRT-PCR was performed on an Eppendorf Realplex 4 Mastercycler using Real MasterMix SYBR Green detection system (Eppendorf) in a total volume of 10 µl loaded in triplicate. Primers for COL-1 (NM_000088), FN (NM_002026), and β-actin (NM_001101, housekeeping gene) were supplied by Primerdesign.
Gene expression values were calculated using the comparative ΔΔCT method and they corresponded to the expression level (fold increase) of the target gene compared to the calibrator sample (untreated cells) taken as unit value by definition19. In all qRT-PCR, the melting curve was performed to confirm the specificity of the SYBR green assay. The qRT-PCR was performed on 5 independent experiments on SSc skin fibroblasts.
Statistical analysis
The statistical analysis was carried out by the nonparametric Mann-Whitney U test to compare unpaired treatments. Any p value lower than 0.05 was considered statistically significant. Final results of the MTT test, immunofluorescence, immunocytochemistry, Western blotting, and qRT-PCR were the mean of the results obtained from the independent experiments performed on in vitro cultures of skin fibroblasts isolated from each patient with SSc and healthy subject. These results were indicated as mean ± SD.
RESULTS
Determination of functional concentrations of macitentan and ACT-132577 able to antagonize the mitogenic effects of ET-1
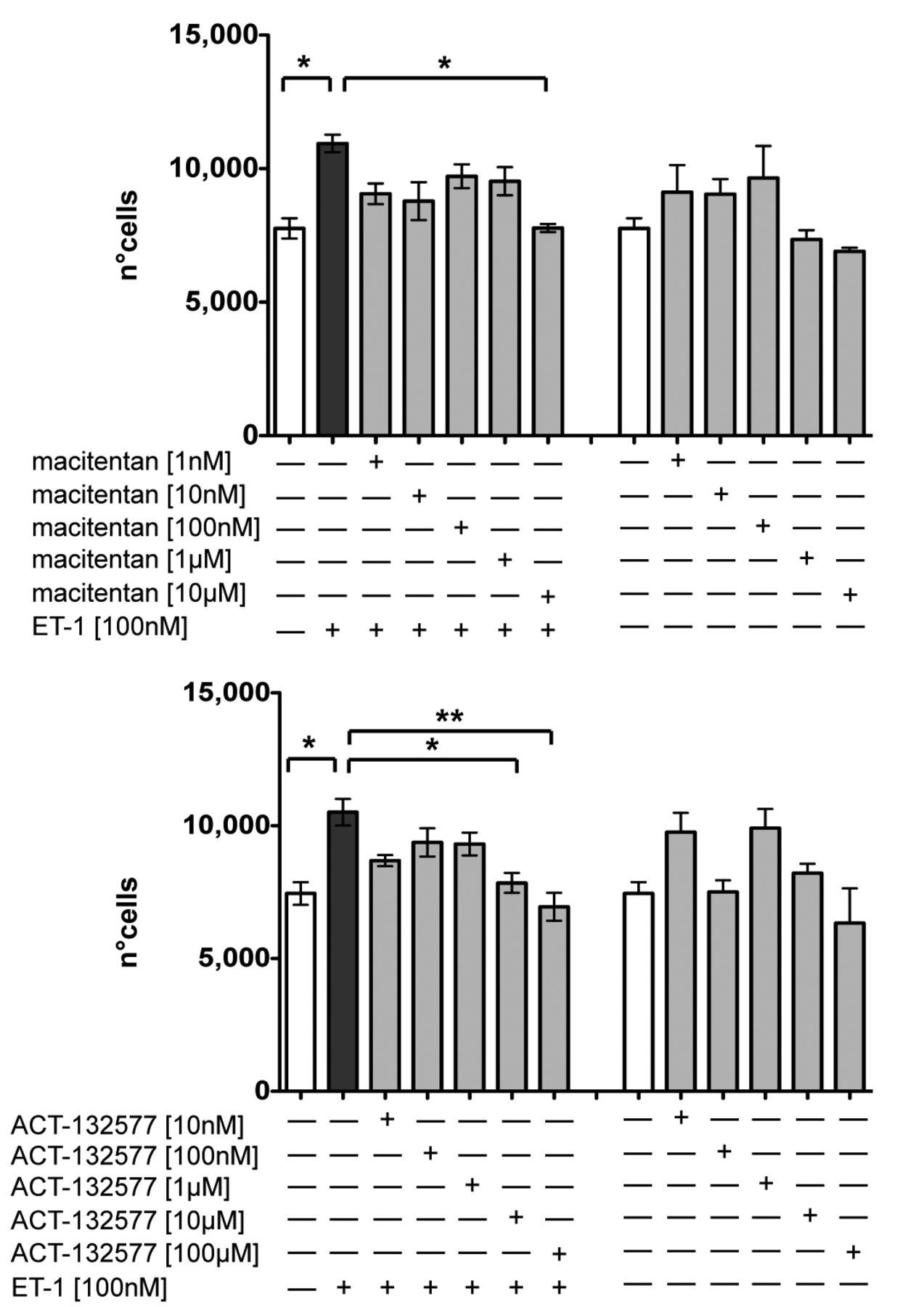
The MTT test showed that 10 µm was the only concentration of macitentan that could significantly antagonize the ET-1–induced increase in cell proliferation (macitentan 10 µm/ET-1 vs ET-1–treated cells, p = 0.03, Figure 1). This concentration did not show any cytotoxic effect, although a limited reduction in cell growth (6.5%) was observed versus untreated skin fibroblasts (Figure 1). The concentrations of ACT-132577 that significantly contrasted the increase in cell proliferation induced by ET-1 were 10 µm and 100 µm (p = 0.016 for ACT-132577 10 µm/ET-1 and p = 0.008 for ACT-132577 100 µm/ET-1 and vs ET-1–treated cells, Figure 1). Compared to untreated cells, the ACT-132577 concentration of 100 µm determined an important cell growth decrease (21%), while the concentration of 10 µm did not show any cytotoxic effect or reduction in fibroblast growth (Figure 1).

Evaluation of cell growth in cultured human control skin fibroblasts. Cell proliferation analysis by methyltetrazolium salt test (MTT test) in cultured skin fibroblasts from healthy subjects (n = 5) after 48 h of treatment. Cultured cells were untreated, treated with ET-1 alone (100 nM), treated with different concentrations of macitentan (range from 1 nM to 10 μM) and after 1 h also stimulated with ET-1, or treated only with different concentrations of macitentan (1 nM to 10 μM; top panel). Cultured cells were untreated, treated with ET-1 alone (100 nM), treated with different concentrations of ACT-132577 (10 nM to 100 μM) and after 1 h also stimulated with ET-1, or treated only with different concentrations of ACT-132577 (10 nM to 100 μM; bottom panel). Data were shown as mean cell number ± SD. * p < 0.05. ** p < 0.01. MTT: 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide test; ET-1: endothelin 1.
Therefore, both macitentan and ACT-132577 seemed to be functionally active at the same concentration (10 µm) in contrasting the ET-1–induced effects on cell proliferation in cultured human control skin fibroblasts. The concentration of 10 µm was the same used for the in vitro treatment of skin fibroblasts with bosentan, because it was already demonstrated in several other studies15,16. Based on MTT results, the concentration of macitentan and ACT-132577 used in subsequent experiments was 10 µm.
Evaluation of α-SMA expression and ECM protein synthesis in cultured SSc and control skin fibroblasts
At basal condition (untreated cells), SSc skin fibroblasts showed a higher expression of α-SMA and a significantly higher COL-1 synthesis compared to control skin fibroblasts (p = 0.016), as observed by immunofluorescence and immunocytochemistry (Supplementary Figure 2 and Supplementary Figure 3A, available online at jrheum.org, and Figure 3). No differences in FN synthesis were observed between both cell types (Supplementary Figure 3A).
Effects of macitentan, ACT-132577, and bosentan on α-SMA expression and ECM protein synthesis in cultured SSc and control skin fibroblasts
As assessed by immunofluorescence and immunocytochemistry, the treatment with macitentan induced a significant reduction in α-SMA expression in cultured SSc skin fibroblasts compared to untreated cells (p = 0.03), whereas ACT-132577 or bosentan did not significantly reduce the expression of this activated myofibroblast marker (Figure 2; Supplementary Figure 2). On these cultured cells, treatment with macitentan also induced a significant decrease in COL-1 synthesis compared to the untreated cells, similar to the decrease observed in bosentan treatment (p = 0.02 and p = 0.016, respectively; Figure 3; Supplementary Figure 3A). ACT-132577 induced a nonsignificant reduction in COL-1 synthesis compared to SSc untreated cells (Figure 3; Supplementary Figure 3A).
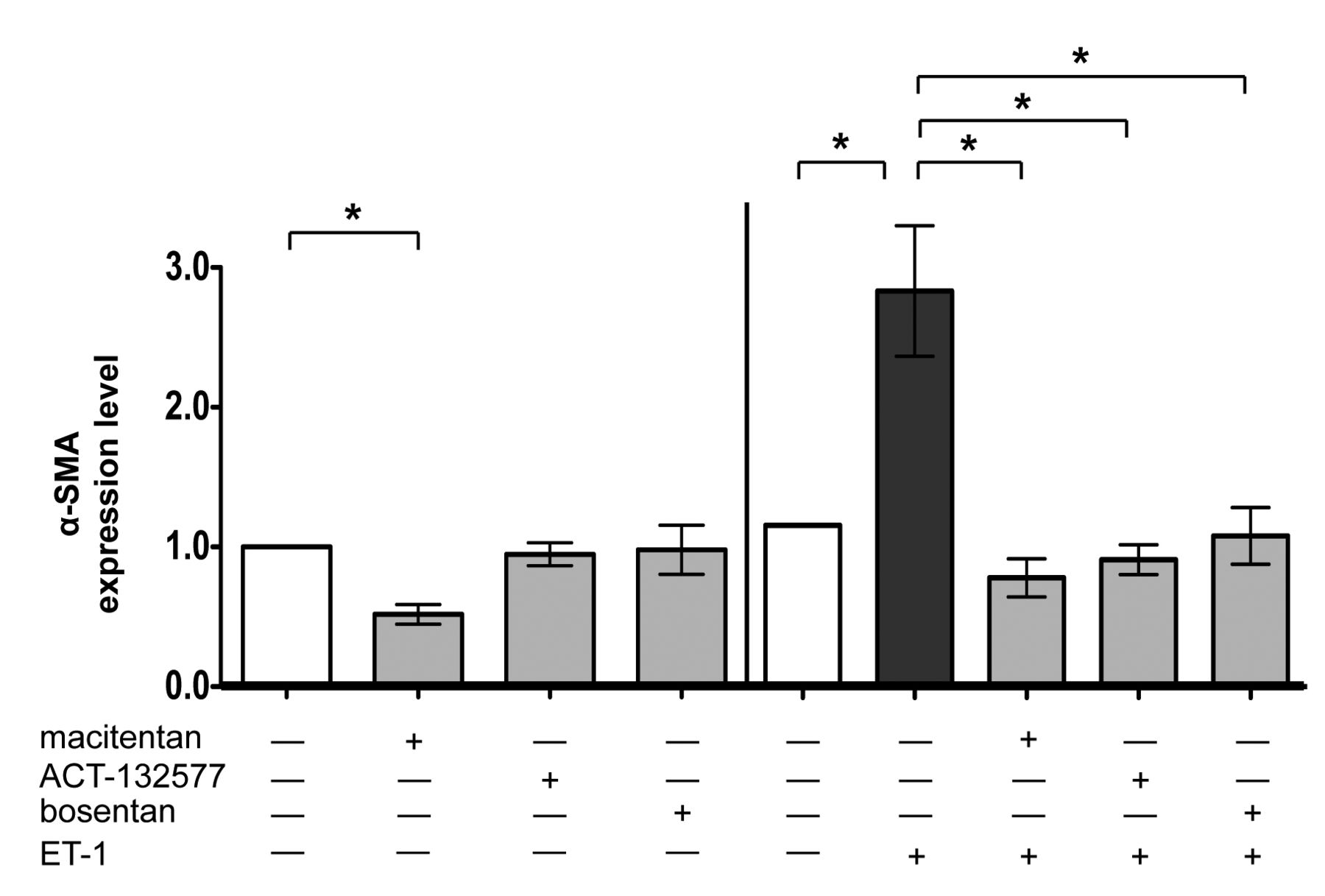
Evaluation of α-SMA expression in cultured SSc skin fibroblasts by immunofluorescence. Immunofluorescence image analysis of α-SMA expression in cultured human skin fibroblasts from patients with SSc (n = 6) after 48 h of treatment. Cultured skin fibroblasts were either untreated or treated with macitentan (10 μM), ACT-132577 (10 μM), or bosentan (10 μM) alone (left part of histogram). Cultured skin fibroblasts were untreated, treated with ET-1 (100 nM), treated with macitentan, ACT-132577, or bosentan (all 10 μM) and after 1 h, also stimulated with ET-1 (right part of histogram). The image analysis was performed by evaluating the same number of cells for each experimental condition by light microscopy (magnification 20×). The values of α-SMA expression obtained from each treatment were normalized to that of untreated cells (taken as unit value by definition) and showed as mean ± SD of α-SMA expression level. * p < 0.05. α-SMA: α-smooth muscle actin; SSc: systemic sclerosis; ET-1: endothelin 1.

Evaluation of ECM protein synthesis in cultured SSc and control skin fibroblasts by immunocytochemistry. Immunocytochemistry image analysis of COL-1 and FN synthesis in cultured skin fibroblasts from patients with SSc (n = 6) and healthy subjects (n = 5) after 48 h of treatment. Cultured SSc and control skin fibroblasts were untreated or treated with macitentan, ACT-132577, or bosentan (all 10 μM, left part of histogram). Cultured SSc and control skin fibroblasts were untreated, treated with ET-1 alone (100 nM), macitentan, ACT-132577, or bosentan (all 10 μM) and after 1 h also stimulated with ET-1 (middle and right parts of histogram). The values of protein synthesis were obtained by evaluating the same number of cells in each experimental condition. The data of COL-1 and FN synthesis were shown as mean ± SD and indicated as level of COL-1. * p < 0.05. ** p < 0.01. ECM: extracellular matrix; COL-1: type I collagen; FN: fibronectin; SSc: systemic sclerosis; ET-1: endothelin 1.
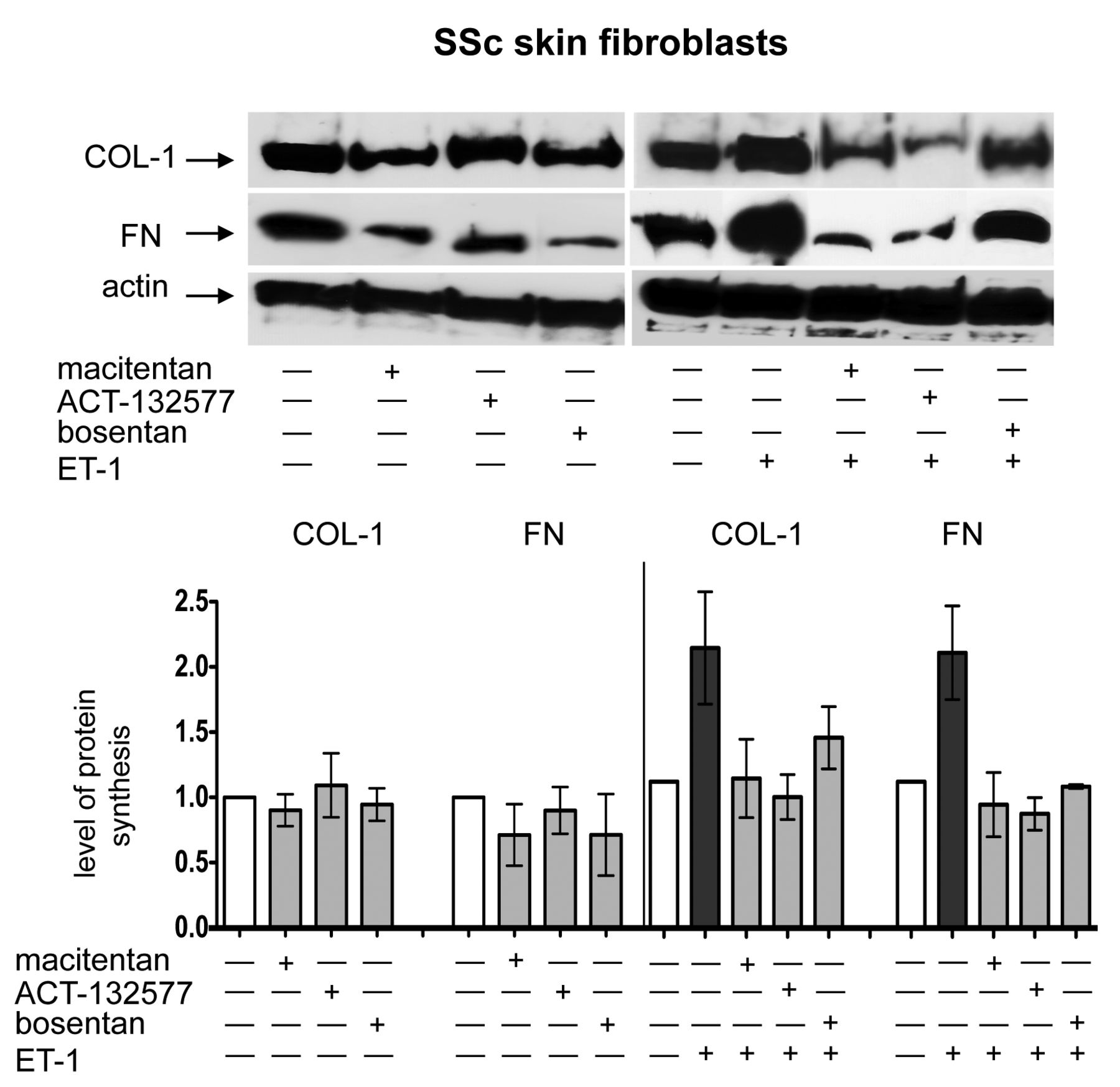
At the same time, no effects were observed by treatment with macitentan, ACT-132577, or bosentan on FN synthesis compared to untreated cells (Supplementary Figure 3A). However, Western blotting showed that treatment with either macitentan or bosentan for 48 h induced a nonsignificant reduction in COL-1 and FN synthesis compared to untreated SSc skin fibroblasts (Figure 4).
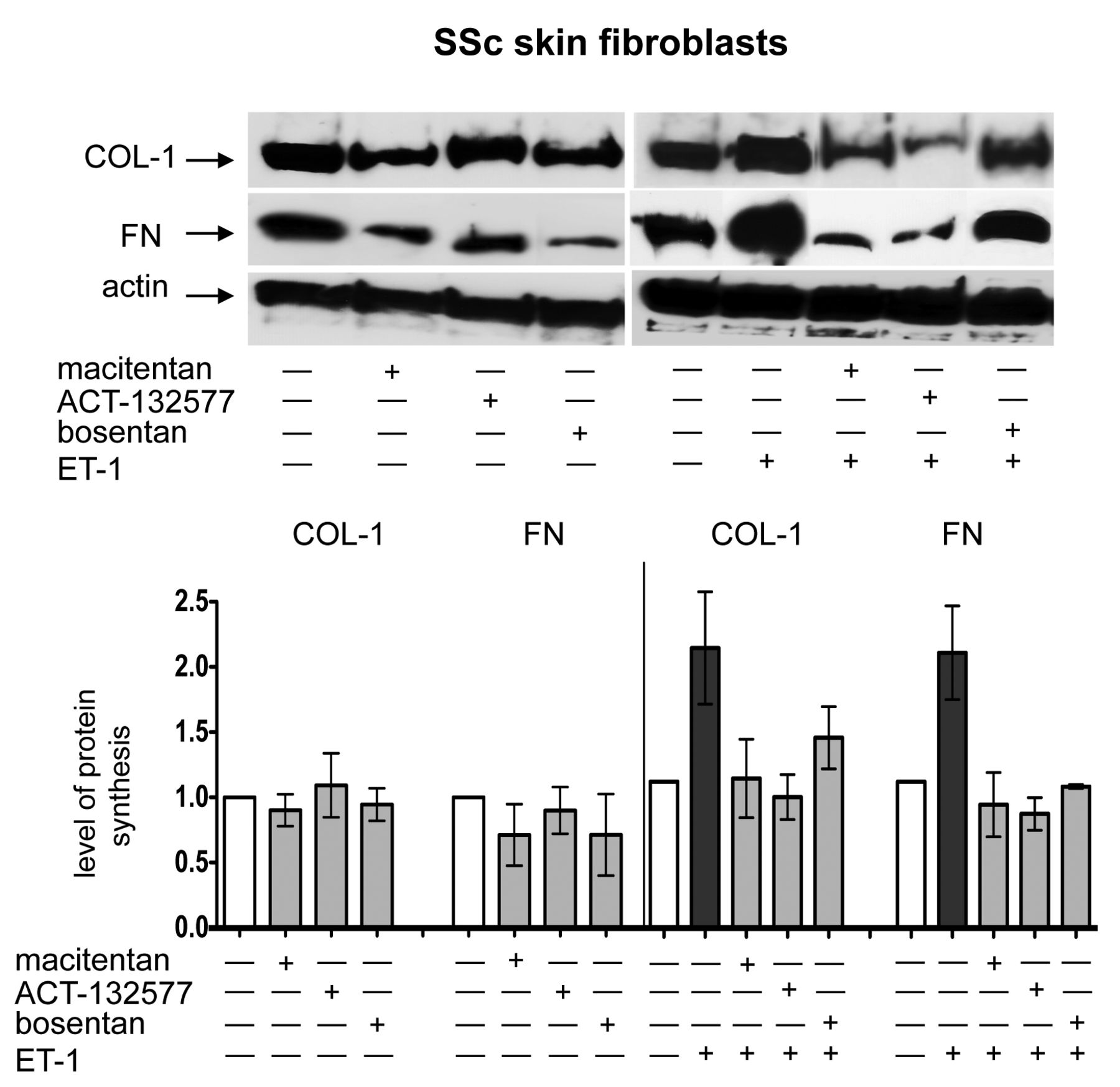
Evaluation of ECM protein synthesis in cultured SSc skin fibroblasts by Western blotting. Western blotting and relative densitometric analysis of COL-1 and FN synthesis in cultured skin fibroblasts from patients with SSc (n = 5) after 48 h of treatment. Cultured skin fibroblasts were either untreated or treated with macitentan (10 μM), ACT-132577 (10 μM), or bosentan (10 μM) alone (left image and part of histogram). Cultured SSc skin fibroblasts were untreated, treated with ET-1 (100 nM), macitentan, ACT-132577, or bosentan (all 10 μM) and after 1 h also stimulated with ET-1 (right image and part of histogram). For each experimental condition, the values of COL-1 and FN synthesis were normalized to that of the corresponding actin. The values of protein synthesis obtained for each treatment were normalized to that of the untreated cells taken as unit value by definition. The data of COL-1 and FN synthesis were shown as mean ± SD and indicated as level of protein synthesis. ECM: extracellular matrix; SSc: systemic sclerosis; COL-1: type I collagen; FN: fibronectin; ET-1: endothelin 1.
On the contrary, in cultured control skin fibroblasts, treatment with macitentan, ACT-132577, or bosentan did not induce any modulatory effects on either α-SMA expression or ECM protein synthesis compared to untreated cells, as observed by immunofluorescence and immunocytochemistry (Supplementary Figure 2 and Supplementary Figure 3A, available online at jrheum.org, and Figure 3).
Effects of macitentan, ACT-132577, and bosentan on ET-1–induced expression of α-SMA and ECM protein synthesis in cultured SSc and control skin fibroblasts
The stimulation by ET-1 induced a significant increase in α-SMA expression, as well as in COL-1 and FN synthesis in cultured SSc skin fibroblasts compared to untreated cells (p = 0.03 for α-SMA, p = 0.02 for COL-1, p = 0.005 for FN; Figure 2 and Figure 3; Supplementary Figures 2 and 3B). Of note, in these cultured cells, the treatment with macitentan significantly antagonized the ability of ET-1 to increase the expression of α-SMA as well as the synthesis of both ECM proteins (macitentan/ET-1 vs ET-1–treated cells, p = 0.03 for α-SMA and COL-1, p = 0.005 for FN; Figure 2 and Figure 3; Supplementary Figures 2 and 3B). Similar results were observed by treatment with ACT-132577, which significantly contrasted the ET-1–induced increase in α-SMA expression and ECM protein synthesis (ACT-132577/ET-1 vs ET-1–treated cells, p = 0.03 for α-SMA and COL-1, p = 0.005 for FN; Figure 2 and Figure 3; Supplementary Figures 2 and 3B). Treatment with bosentan was confirmed to significantly antagonize the ability of ET-1 to increase the α-SMA expression, COL-1, and FN synthesis in cultured SSc skin fibroblasts (bosentan/ET-1 vs ET-1–treated cells, p = 0.03 for all ECM proteins; Figures 2 and 3; Supplementary Figures 2 and 3B)8. All these data were obtained by immunofluorescence and immunocytochemistry.
The effects of ET-1, macitentan, ACT-132577, and bosentan on α-SMA expression and ECM protein synthesis were also evaluated in cultured control skin fibroblasts. On these cells, ET-1 induced the expression of α-SMA and a significant increase in COL-1 and FN synthesis compared to untreated fibroblasts (p = 0.03 for both ECM proteins; Supplementary Figure 2, Supplementary Figure 3B, and Figure 3). Treatment with macitentan, ACT-132577, or bosentan inhibited the expression of this myofibroblast phenotype marker and significantly antagonized the increase in ECM protein synthesis when the cells were subsequently stimulated with ET-1 (vs ET-1–treated cells for COL-1 and FN, p = 0.03; Supplementary Figure 2, Supplementary Figure 3B, and Figure 3).
No significant differences were observed between the ETRA in contrasting the ET-1–induced increase in ECM protein synthesis in both cultured SSc and control skin fibroblasts (Figure 3). All these data were obtained by immunofluorescence and immunocytochemistry.
Western blotting analysis showed similar results to those reported above, primarily on COL-1 and FN synthesis (Figure 4; Supplementary Figure 4, available online at jrheum.org).
Effects of macitentan, ACT-132577, and bosentan on ECM gene expression in cultured SSc skin fibroblasts
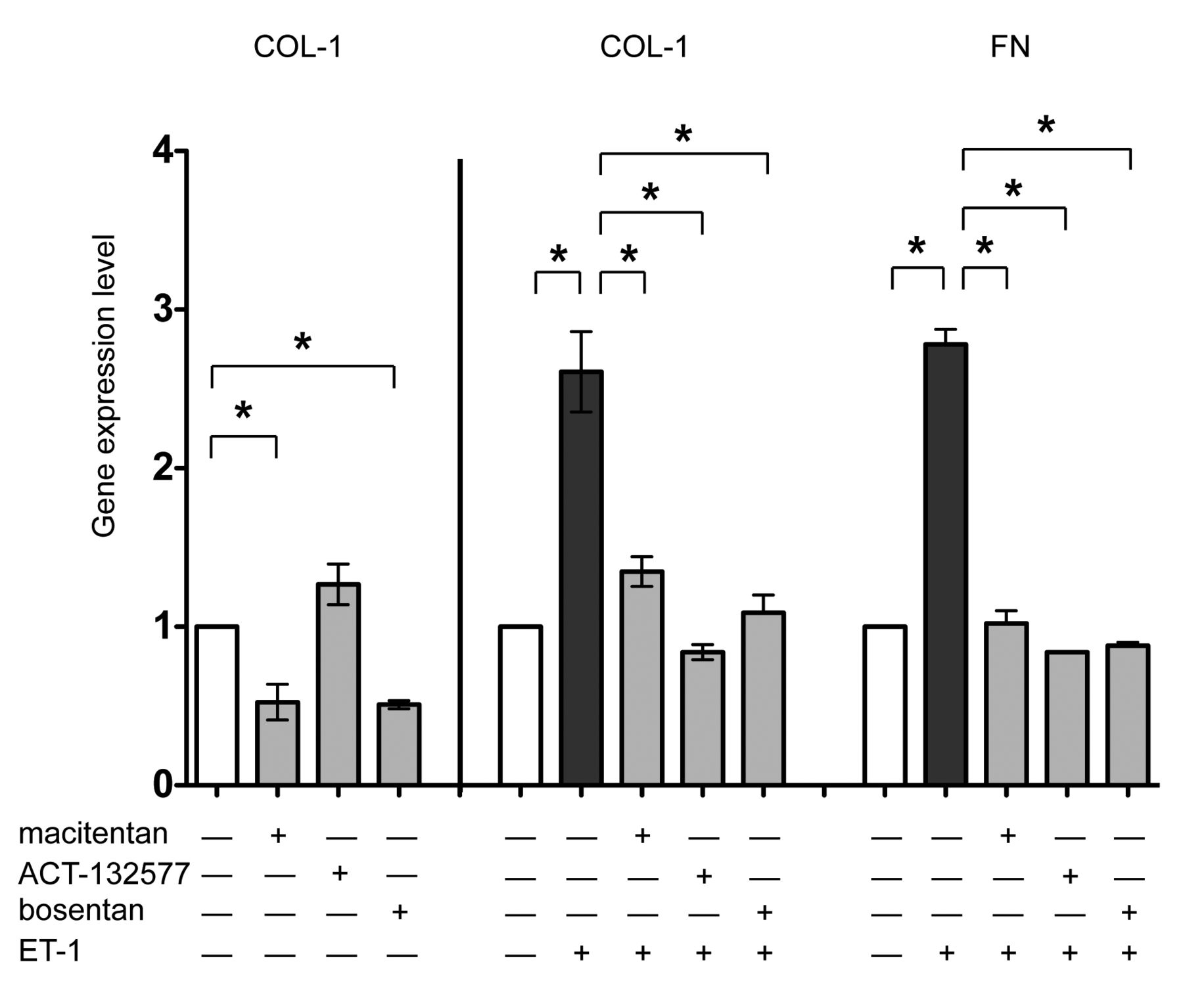
In cultured SSc skin fibroblasts, qRT-PCR showed that treatment with macitentan as well as bosentan significantly reduced the gene expression of COL-1 compared to the untreated cells (p = 0.047 for macitentan; p = 0.03 for bosentan), confirming the results obtained by immunocytochemistry (Figure 5). Conversely, ET-1 induced a significant increase in the gene expression of COL-1 and FN compared to the untreated cells (p = 0.03 for both; Figure 5). Treatment with macitentan, ACT-132577, or bosentan significantly antagonized the ability of ET-1 to increase the gene expression of these ECM molecules (macitentan/ET-1, ACT-132577/ET-1, and bosentan/ET-1 vs ET-1, p = 0.03 for COL-1, p < 0.05 for FN; Figure 5).
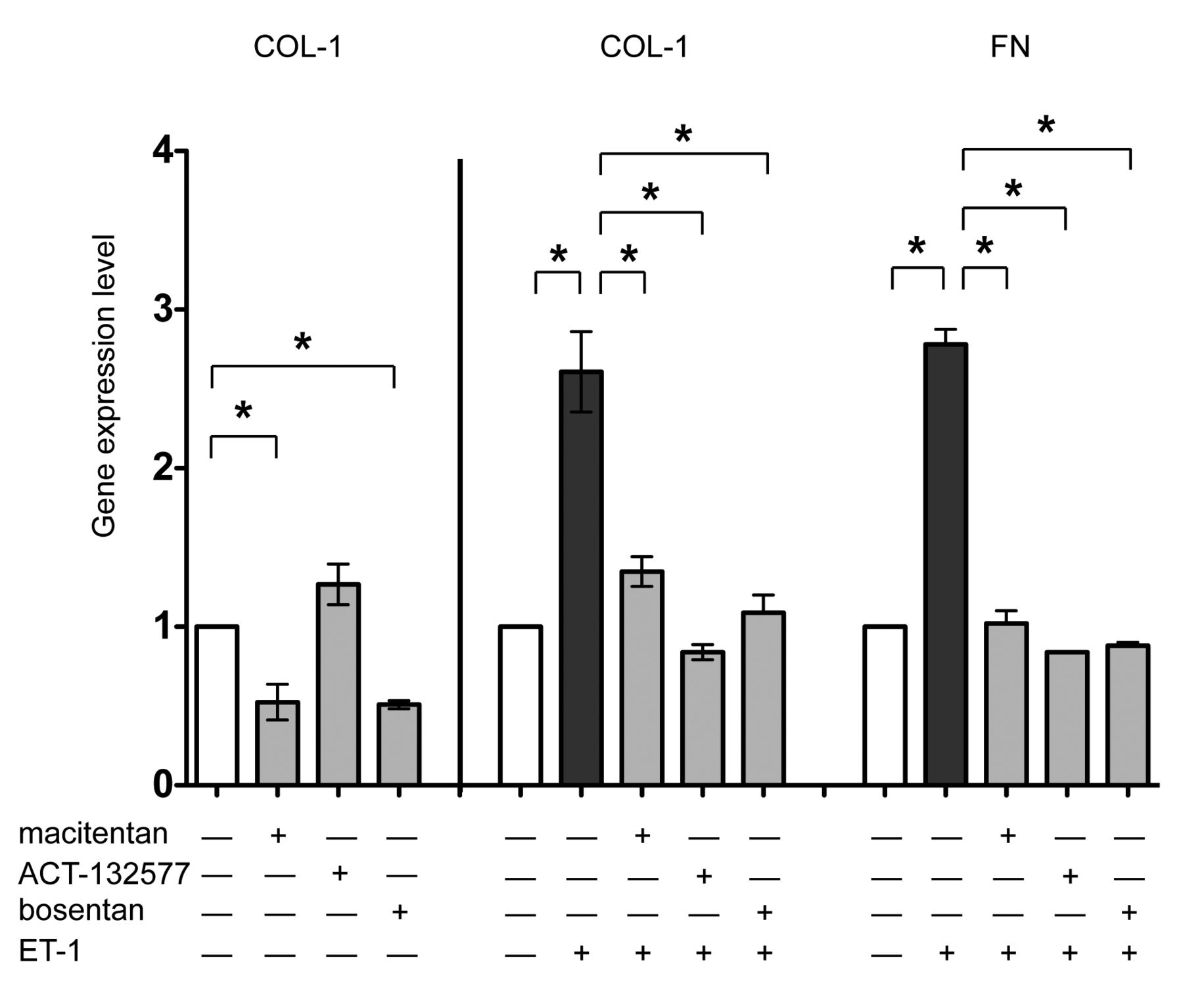
Evaluation of ECM protein expression in cultured SSc skin fibroblasts by qRT-PCR. qRT-PCR analysis of COL-1 and FN gene expression in cultured skin fibroblasts from patients with SSc (n = 5) after 48 h of treatment. Cultured skin fibroblasts were either untreated or treated with macitentan (10 μM), or ACT-132577 (10 μM), or bosentan (10 μM) alone (left part of histogram). Cultured skin fibroblasts were untreated, treated with ET-1 (100 nM), macitentan, ACT-132577, or bosentan (all 10 μM) and after 1 h also stimulated with ET-1 (middle and right part of histogram). For each treatment, the gene expression values correspond to a fold expression (expression level or fold increase) of the target gene compared to the calibrator sample (untreated cells), taken as unit value by definition. The data of COL-1 and FN expression were shown as mean ± SD and indicated as gene expression levels. * p < 0.05. ECM: extracellular matrix; SSc: systemic sclerosis; qRT-PCR: quantitative real-time PCR; COL-1: type I collagen; FN: fibronectin; ET-1: endothelin 1.
DISCUSSION
The results of our in vitro study suggest that macitentan, a new tissue-targeting dual ETRA, may downregulate the expression of α-SMA as a marker of an activated myofibroblast phenotype in cultured SSc skin fibroblasts. The downregulatory effect of macitentan on the α-SMA expression was found to be greater than that mediated by its active metabolite ACT-132577 or bosentan. This ability might be attributed to the higher affinity of macitentan for ET receptors, especially for ETA, which seems involved in the in vitro myofibroblast phenotype induction12,20. It might be interesting to evaluate the effects of macitentan on α-SMA expression in comparison with selected ETA receptor antagonists such as sitaxentan or ambrisentan. Of note, similar to bosentan, macitentan reduced the basal level of COL-1 synthesis, suggesting that both ETRA may attenuate the transition into the myofibroblast phenotype from human SSc skin fibroblasts, at least in vitro.
The ability of macitentan to antagonize the increase in α-SMA expression and ECM protein synthesis was also maintained after ET-1 stimulation both in cultured SSc and control skin fibroblasts. In the same way, the treatment with ACT-132577 as well as bosentan blocked the ET-1–induced increase in α-SMA expression and ECM protein synthesis.
All these observations suggest that ET-1 might be an important molecule involved in the activation and maintenance of the activated myofibroblast phenotype in SSc8,9.
One recent study confirms inhibitory effects of macitentan (limited to the α-SMA expression) in cultured skin fibroblasts from patients with SSc and healthy subjects using a combination of very low concentrations of both macitentan and ACT-132577 (concentrations of 1 µm and 6 µm, respectively)17. Conversely, our study found inhibitory effects on α-SMA expression and also on ECM protein synthesis for both compounds tested individually in cultured skin fibroblasts using slightly higher concentrations (macitentan and ACT-132577 10 µm).
ACT-132577 was found to be active in vitro and without any toxic effect when used in the same effective concentration of the parent compound (macitentan), even if it was about 5- to 10-fold less effective than macitentan in its capability to inhibit the binding between ET-1 and its receptors12,21,22.
The results of our study are in agreement with investigations showing that ET-1 mediates in vitro myofibroblast differentiation of lung fibroblasts and their ECM protein synthesis through the interaction with its receptors7,8,9,23.
Interestingly, one study showed that ET-1 promotes experimental cardiac fibrosis, inducing the activation of the endothelial-to-mesenchymal transition (EndoMT) process24. The EndoMT process is characterized by the ability of vascular endothelial cells to express myofibroblast markers (i.e., α-SMA, vimentin, S100A4/fibroblast specific protein 1) and to differentiate into myofibroblasts. This mechanism seems to be implicated in several fibrotic conditions, such as idiopathic pulmonary hypertension, as well as cardiac, renal, and skin fibrosis in diabetes25,26. Moreover, EndoMT has been suggested to have a role in the pathogenesis of SSc27,28. These observations further support that ET-1 might have a role in the early phase of the fibrotic process, following the microvascular damage3,25,29,30.
ET-1 was confirmed to increase the expression of α-SMA, COL-1, and FN in cultured SSc and control skin fibroblasts, further suggesting a possible role in the complex pathogenesis of SSc. Based on these observations, ET-1 might contribute to the transition from the early microvascular damage to the fibrotic progression of the disease. Macitentan seems to have important effects in contrasting the profibrotic myofibroblast phenotype in cultured SSc skin fibroblasts, reducing the basal levels of α-SMA and COL-1 expressions. Therefore, macitentan, its active metabolite ACT-132577, and bosentan might have downregulatory effects in the early phases of the fibrotic process that characterizes SSc disease progression31,32,33. Based on present results, a new study is in progress to investigate the functional relevance of macitentan and its active metabolite, evaluating their ability to contrast the activation of intracellular signaling pathways linked to the profibrotic effects mediated by ET-1 and to contrast the enhanced contractile fibrotic phenotype of SSc skin fibroblasts.
ONLINE SUPPLEMENT
Supplementary data for this article are available online at jrheum.org.
Footnotes
-
Supported by a research grant from Actelion Pharmaceuticals Ltd.
- Accepted for publication November 20, 2014.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}