
Abstract
Objective. To elucidate the potential role of follicular helper T cells (TFH) and interleukin 10 (IL-10)+ B cells in the development of systemic lupus erythematosus (SLE).
Methods. The numbers of peripheral blood CD27+, CD38+, CD86+, CD95+, IL-10+ B cells, and inducible T cell costimulator (ICOS)+, programmed death-1 (PD-1)+, IL-21+, CXCR5+CD4+ TFH-like cells were examined in 23 patients with new onset SLE and 20 healthy controls (HC).
Results In comparison with HC, significantly reduced numbers of CD19+ and IL-10+ B cells, but increased numbers of CD27high, CD86+, CD95+ B cells, CXCR5+CD4+, ICOS+, PD-1+, and IL-21+ TFH-like cells were detected, which were accompanied by higher levels of serum IL-21, but lower levels of IL-10 in the patients. Treatment with anti-SLE therapy modulated the imbalance of different subsets of B and TFH-like cells. The levels of serum IL-21 and IL-10 were positively correlated with the numbers of CD4+CXCR5+ TFH-like and CD19+CD5+CD1d+ B cells in the patients, respectively. The numbers of CD27high B cells were correlated positively with IL-21+ TFH-like cells, but negatively with IL-10+ B cells. The values of SLE Disease Activity Index, C3, and erythrocyte sedimentation rate were correlated positively with serum IL-21, but negatively with IL-10 in those patients.
Conclusion. Our data indicate that the imbalance of IL-21+ TFH-like, CD27high, and IL-10+ B cells may be associated with the pathogenesis of SLE, and levels of serum IL-21 and IL-10 may be valuable for evaluating disease activity in SLE.
- SYSTEMIC LUPUS ERYTHEMATOSUS
- B CELLS
- REGULATORY B CELL
- FOLLICULAR HELPER T-LIKE CELLS
- INTERLEUKIN 21
- INTERLEUKIN 10
Systemic lupus erythematosus (SLE) is a prototypic systemic autoimmune disease characterized by high levels of serum autoantibodies against different components of cells, leading to systemic tissue damage1,2. Mounting evidence has demonstrated that immunological disturbance plays a role in the pathogenesis of SLE3 and imbalance of different subsets of B cells is crucial for the initiation and perpetuation of SLE in both humans and rodents4,5. Previous studies have shown that decreased numbers of CD19+CD27+ memory B cells and increased numbers of CD19+ CD27high plasmablast B cells are associated with the development of SLE6,7,8. Indeed, the percentages of CD19+ CD27high and CD19+CD38+ plasma B cells are positively correlated with disease severity in patients with SLE9,10. In addition, CD86 and CD95 are expressed by activated and exhausted B cells, respectively11. It is notable that CD19+CD5+CD1d+ B cells can produce interleukin 10 (IL-10) and negatively regulate inflammation in murine models of autoimmune diseases12,13,14. The numbers of IL-10+ B cells are associated negatively with concentrations of serum autoantibodies, suggesting that IL-10 produced by CD19+CD5+CD1d+ B cells may downregulate humoral responses15. However, little is known about how these different subsets of B cells are present in Chinese patients with new-onset SLE.
CCD4+ T cells, particularly CD4+CXCR5+ follicular helper-like T (TFH-like) cells, are crucial regulators of the activation and functional differentiation of B cells16. Activated TFH-like cells express inducible T cell costimulator (ICOS) and programmed death-1 (PD-1) and secrete IL-21, an important factor for their function17,18,19. Previous studies have shown that TFH can help the formation of germinal centers, high-affinity long-living plasma cells, and memory B cells20. Indeed, dysfunction of TFH cells in the germinal centers and aberrant activation of B cells are associated with pathogenic humoral responses and SLE development21. Human peripheral blood CXCR5+CD4+ T cells share properties with TFH cells22. However, it is unclear whether and how the numbers of peripheral blood CXCR5+CD4+ TFH-like cells are associated with the development of SLE in Chinese patients.
Currently, patients with new-onset SLE are treated with disease-modifying antirheumatic drugs and prednisone. Previous studies have shown that the anti-SLE therapies can effectively control clinical symptoms and benefit patients with new-onset SLE23,24,25. However, little is known about how the anti-SLE therapies affect the numbers of different subsets of B cells and TFH-like cells in Chinese patients with SLE.
We investigated the numbers of different subsets of peripheral blood B cells and TFH-like cells, and concentrations of serum IL-10 and IL-21 in patients with new-onset SLE and healthy controls, and the potential association of the numbers of these cells or the concentrations of serum IL-10 and IL-21 with clinical measures in those patients. Further, we examined the dynamic changes in the numbers of these cells and the concentrations of serum IL-10 and IL-21 in those patients following the anti-SLE therapies. Our findings indicate that increased numbers of activated B cells and TFH-like cells, together with deceased numbers of IL-10+ B cells, were present in patients with new onset SLE and that treatment with the anti-SLE therapies not only effectively controlled clinical symptoms, but also modulated the imbalance of different subsets of B cells and TFH-like cells in patients with SLE.
MATERIALS AND METHODS
Subjects
A total of 23 patients with new-onset SLE were recruited from the inpatient service of the First Hospital of Jilin University, Changchun, China. Individual patients with SLE were diagnosed according to the criteria established by the American College of Rheumatology13. The disease activity of individual patients was assessed using the SLE Disease Activity Index (SLEDAI). Another 2 sex-, age-, and ethnicity-matched healthy controls (HC) were also recruited. Individuals were excluded if they had a history of other autoimmune diseases, recent infection, or immunosuppressant or glucocorticoid therapies within the past 6 months. The experimental protocol was established according to the guidelines of the Declaration of Helsinki and was approved by the Human Ethics Committee of Jilin University. Written informed consent was obtained from each participant.
The demographic and clinical data of individual subjects were collected, including age, sex, and current medications. Their demographic and clinical characteristics are summarized in Table 1.
The demographic and clinical characteristics of subjects. Data are median (range) of each group of subjects unless otherwise indicated.
Treatment
After being admitted, individual patients were treated with anti-SLE therapies, dependent on the values of SLEDAI. A total of 13 patients with a SLEDAI value < 9 were treated orally with 5 mg prednisone (Wyeth) daily for 1 week and then with 400 mg hydroxychloroquine (Shanghai Xinyi Pharmacy) daily for 12 weeks; 4 patients with a SLEDAI value of < 14 were treated orally with 10 mg prednisone, 20 mg leflunomide daily (Fujian Huitian Pharmacy) and 10 mg methotrexate (MTX, Shanghai Xinyi Pharmacy) once per week for 12 weeks; 6 patients with a SLEDAI value of ≥ 15 were treated with 10 mg prednisone daily, 10 mg MTX once per week, and 150 mg cyclophosphamide (Boehringer Ingelheim) once per 3 weeks for 12 weeks, as described13. After being discharged, those patients visited the outpatient service of our department for office visits and laboratory tests.
Clinical measurements
Peripheral venous blood samples were obtained from individual patients for laboratory tests before treatment and 4 and 12 weeks after the initial treatment. The routine laboratory investigations included full blood counts, the concentrations of serum C-reactive protein (CRP), and complement factors C3 and C4, which were determined by scattered turbidimetry on a Siemens special protein analyzer (Siemens Healthcare Diagnostics Products).
Flow cytometry analysis
The percentages of different subsets of peripheral blood B cells in individual subjects were analyzed by flow cytometry using specific antibodies, as described26. Briefly, 100 µl of blood samples were stained with PerCP-anti-CD19, phycoerythrin (PE)-anti-CD38, antigen-presenting cell (APC)-anti-CD86, PE-anti-CD27, and APC-anti-CD95 antibodies (BD PharMingen) for 30 min. The matched mouse isotype controls included FITC-IgG1, PE-Ig2a, and PerCP-IgG1. After lysis of erythrocytes, the remaining peripheral blood mononuclear cells (PBMC) were characterized using FACSCalibur (BD Biosciences) and FlowJo software (v7.6.2; TreeStar). At least 20,000 events of each sample were analyzed and the numbers of each type of cells were calculated, based on the total numbers of PBMC. To ensure the quality of each batch of data during the longitudinal study, we used the same lot of antibodies from identical manufacturers, according to the same protocol, in a blinded manner.
Further, the percentages of peripheral blood IL-10+ B cells in individual subjects were also determined by flow cytometry. Briefly, PBMC were isolated from individual subjects by density-gradient centrifugation using Ficoll-Paque Plus (Amersham Biosciences). PBMC (4 × 106/ml) were stimulated in duplicate with 50 ng/ml phorbol myristate acetate (PMA), 1.0 µg/ml ionomycin in the presence or absence of 50 ng/ml lipopolysaccharide (LPS; Sigma-Aldrich) in 10% fetal calf serum RPMI-1640 (Hyclone) in 24-well U-bottom tissue culture plates (Costar) for 2 h, and then cultured for another 4 h in the presence of monensin (2 µl, GolgiStop; BD Biosciences). PBMC in medium alone served as negative controls. The cells were stained in duplicate with APC-anti-CD19, PerCP-anti-CD5, and PE-anti-CD1d (BD Biosciences) or isotype controls for 30 min, fixed, and permeabilized using the permeabilization solution (BD Biosciences), followed by intracellular staining with FITC-anti-IL-10. The percentages of IL-10+ B cells were determined by flow cytometry.
In addition, the percentages of IL-21+ TFH-like cells in individual subjects were determined by flow cytometry. PBMC (2 × 106/well) were stimulated with PMA/ionomycin in the presence of monensin for 6 h, as described above. The cells were stained in duplicate with PerCP-anti-CXCR5 (Biolegend) and APC-anti-CD4 (BD Biosciences) or isotype controls for 30 min, fixed, and permeabilized, followed by intracellular staining with Alexaflour-anti-IL-21 (BD PharMingen). Further, PBMC (5 × 105/tube) were stained in duplicate with PerCP-anti-CXCR5 (Biolegend) and APC-anti-CD4, PE-anti-ICOS, FITC-anti-PD-1, or isotype-matched controls (BD Sciences) for 30 min. After being washed with phosphate buffered saline (PBS), the cells were characterized on a FACSCalibur (BD), and at least 20,000 events were analyzed by FlowJo software (v5.7.2). The numbers of different subsets of peripheral blood lymphocytes were calculated by multiplying the total numbers of PBMC with the percentage of the subset of lymphocytes.
ELISA
The concentrations of serum IL-21 and IL-10 in individual subjects were determined by ELISA using human IL-21 and IL-10 ELISA kits, according to the manufacturers’ instructions (Roche Diagnostics). Briefly, individual wells in flat bottom, high-binding capacity 96-well ELISA plates (Greiner Bio-One) were coated with 100 µl of anti-IL-10 or anti-IL-21 in carbonate bicarbonate buffer (pH 9.6) and kept at 4ºC overnight. After being washed with 0.5% Tween-20 in PBS, individual wells were blocked with 300 µl of 1% bovine serum albumin (BSA, Sigma) for 2 h at room temperature (RT). After being extensively washed with PBS, 100 µl of serum samples at 1:2 dilutions or recombinant standard proteins at different concentrations in 0.1% BSA PBS were added in triplicate into individual wells and incubated at RT for 2 h. After being washed, individual wells were added with 100 µl of biotinylated anti-IL-10 or anti-IL-21, respectively, and incubated at RT for 2 h. The wells were exposed to 100 µl of freshly prepared streptavidin-horseradish peroxidase (1:200 dilutions, R&D Systems) in 0.1% BSA PBS at RT for 1 h. After wells were washed 3 times with PBS, 100 µl of enzyme substrate tetramethylbenzidine containing hydrogen peroxide (R&D Systems) at RT was added for 20 min in the dark, and the reactions were stopped by 100 µl of sulfuric acid, followed by reading the optical density at 450 nm in an microplate reader (Bio-Rad 680). The concentrations of serum IL-21 and IL-10 of individual samples were determined, according to the standard curve established using the recombinant IL-21 and IL-10 provided. The limits of detection for IL-21 and IL-10 were 10 ng/l and 2.5 ng/l, respectively.
Statistical analysis
Data are present as median and range, except when specified. All data were tested for normal distribution by Shapiro-Wilk test and for homogeneity of variances by Levene’s test. If the data were normally distributed and had similar variances, the data were analyzed by 1-way ANOVA and posthoc Tukey HSD test. If the data were not normally distributed and did not have similar variances, the data were analyzed by nonparametric Kruskal-Wallis test and posthoc Bonferroni correction test to compare the medians among multiple groups. If statistically significant, the difference between 2 groups was analyzed by Mann-Whitney U test. The relationship between the variables was evaluated using the Spearman’s rank correlation test. All statistical analyses were performed using the SPSS 18 for Windows. A p value < 0.05 was considered statistically significant.
Results
Increased numbers of circulating activated and exhausted B cells in patients with new-onset SLE
To determine the role of different subsets of B cells in the development of SLE, 23 patients with new-onset SLE and 20 age-, sex-, and ethnicity-matched HC were recruited. As expected, there was no significant difference in the distribution in age and sex between these 2 groups of subjects (Table 1). Further, most patients developed anti-dsDNA and anti-Sm autoantibodies, and all patients had active SLE. As compared with HC in this population, the patients displayed significantly higher concentrations of CRP, erythrocyte sedimentation rate (ESR), C3, and C4, but lower numbers of white blood cells (WBC) and PBMC, thereby demonstrating systemic inflammation.
After treatment for 4 or 12 weeks, there were 16 and 20 patients with SLEDAI values < 6, respectively. Levels of disease activity, positive rates of serum autoantibody detection, and concentrations of serum CRP as well as ESR were significantly reduced in these patients (Table 1) in comparison with levels before treatment. The levels of serum C3 and C4 decreased, while the numbers of WBC and PBMC increased in these patients. Therefore, the anti-SLE treatment effectively controlled systemic inflammation in these patients.
Flow cytometric analysis indicated that there was a significant difference in the percentages of different subsets of B cells among different groups of subjects (p < 0.05, determined by Kruskal-Wallis test and posthoc Bonferroni correction test), except for the frequency of CD19+CD86+CD38- B cells (Figure 1 and 2). The numbers of circulating CD19+ B cells in the patients were significantly lower than those in the HC. Further quantitative analysis revealed that the numbers of circulating CD19+CD38+, CD27highCD19+, CD19+CD86+, CD19+CD86+CD38+, CD19+CD95+, CD19+CD95+CD27+, and CD19+CD95+CD27− B cells in the patients were significantly greater than those in the HC (Figures 1 and 2). However, there was no significant difference in the numbers of CD19+CD27+ and CD19+CD86+CD38- B cells between the patients and HC in this population (data not shown). Following anti-SLE treatment for 12 weeks, the numbers of circulating CD19+CD38+ and CD19+ CD27high B cells were gradually reduced, but the numbers of CD19+ did not significantly change in these patients. Similarly, the numbers of circulating CD19+CD86+, CD19+CD86+CD38+, CD19+CD95+, CD19+CD95+CD27+, and CD19+CD95+CD27−, but not CD19+CD86+CD38- B cells significantly decreased, particularly at 12 weeks post- treatment. Similarly, we observed that the percentages of each type of B cells, except for the percentages of CD19+CD86+CD38− and CD19+CD95+CD27+ B cells, gradually decreased in the patients following treatment (Figure 1). Collectively, the anti-SLE treatment not only effectively improved clinical symptoms, but also mitigated autoimmunity-related B cell activity in patients with SLE.

FACS analyses of B cells. Peripheral blood samples were obtained from individual subjects at the indicated timepoints and stained with PerCP-anti-CD19, PE-anti-CD38, APC-anti-CD86, PE-anti-CD27, and APC-anti-CD95 antibodies or isotype-matched IgG. The cells were gated initially on living mononuclear cells (top left) and then on CD19+ B cells (lower left). Subsequently, the percentages of CD19+CD38+, CD19+CD27high, CD19+CD86+, CD19+CD86+CD38+, CD19+CD86+CD38-, CD19+CD95+, CD19+CD95+CD27+, and CD19+CD95+CD27- B cells in total B cells were determined by flow cytometry. Data shown are representative charts of flow cytometry analysis of different subsets of B cells in individual groups of subjects (n = 20 for the HC, n = 23 for the patients at 0 week, n = 16 for the patients at 4 weeks, and n = 20 for the patients at 12 weeks post-treatment). PE: phycoerythrin; APC: antigen-presenting cells.

Quantitative analysis of different subsets of B cells. The numbers of different subsets of B cells in individual subjects were calculated. Data are expressed as the mean numbers of each type of B cells in individual subjects. The horizontal lines indicate the median values for each group at the indicated timepoints.
Decreased numbers of CD19+CD5+CD1d+IL-10+ B cells in the patients with new-onset SLE
Previous studies have shown that CD5+CD1d+ B cells can produce IL-10 and negatively regulate autoimmune diseases in rodents12,13,14. We characterized the numbers of these B cells and found that there was a significant difference in the numbers of these B cells and in the levels of serum IL-10 among the different groups of subjects (p < 0.05, determined by Kruskal-Wallis test and posthoc Bonferroni correction test, Figure 3). The numbers of CD19+CD5+, CD19+CD5+CD1d+, and CD19+CD5+CD1d+IL-10+ B cells in patients with new-onset SLE were significantly lower than those in HC (Figure 3B, C, and D). The numbers of CD19+CD5+ and CD19+CD5+CD1d+ B cells were significantly correlated in those patients (r = 0.604 p = 0.002). Further analysis indicated that the concentrations of serum IL-10 in the patients were also significantly lower than those in the HC (Figure 3E). The concentrations of serum IL-10 were correlated positively with the numbers of CD19+CD5+CD1d+ B cells in those patients (r = 0.623, p = 0.002). In addition, the concentrations of serum IL-10 and the numbers of CD19+CD5+CD1d+IL-10+ B cells were correlated negatively with the numbers of CD27highCD19+ B cells in those patients (r = −0.475, p = 0.022; r = −0.468, p = 0.024), respectively. Of note, following the anti-SLE treatment for 12 weeks, the numbers of CD19+CD5+, CD19+CD5+CD1d+, and CD19+CD5+CD1d+IL-10+ B cells and the concentrations of serum IL-10 in those patients gradually increased to levels similar to that of HC. Similarly, we observed that the percentages of CD19+CD5+CD1d+ and CD19+CD5+CD1d+IL-10+ regulatory B cells, but not CD19+CD5+, gradually increased following treatment in the patients (Figure 3). These data suggest that decreased numbers of CD19+CD5+CD1d+IL-10+ B cells may be associated with the early process of SLE in Chinese patients.
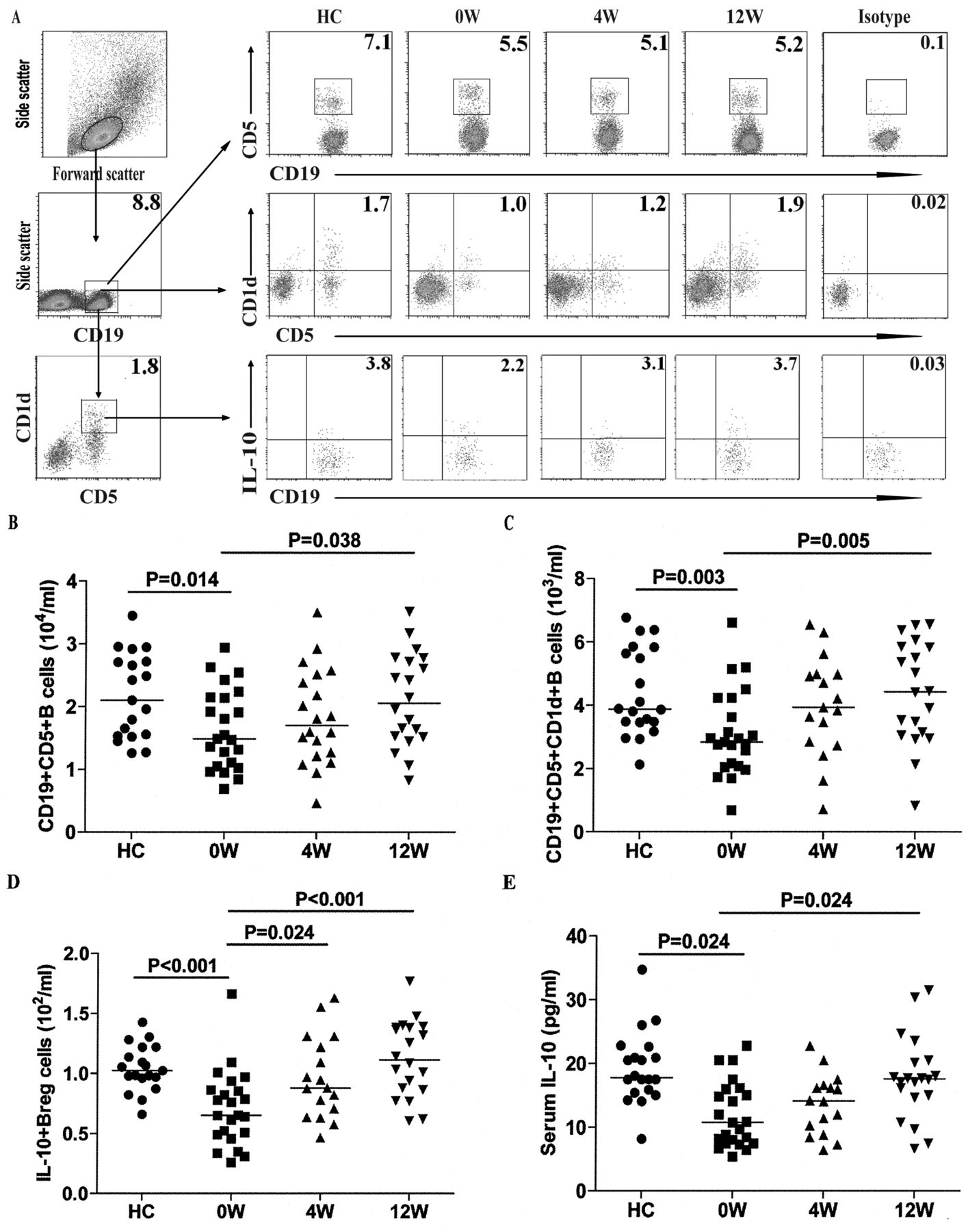
Analysis of peripheral blood interleukin 10+ (IL-10+) B cells and the levels of serum IL-10. PBMC were isolated from individual subjects at the indicated timepoints and stimulated with PMA/ionomycin/LPS in vitro. Subsequently, the cells were stained with APC-anti-CD19, PerCP-anti-CD5, and phycoerythrin-anti-CD1d for 30 min, fixed, and permeabilized, followed by intracellular staining with FITC-anti-IL-10. The numbers of peripheral blood CD19+CD5+, CD19+CD5+CD1d+, and CD19+CD5+CD1d+IL-10+ B cells in individual subjects were characterized by flow cytometry. The concentrations of serum IL-10 in individual subjects were determined by ELISA. Data shown are representative FACS charts or expressed as the mean numbers of each type of cells per ml of peripheral blood in individual subjects from 2 separate experiments. A. Representative FACS charts. B, C, D. Numbers of CD19+CD5+, CD19+CD5+CD1d+, and CD19+CD5+CD1d+IL-10+ B cells. E. Levels of serum IL-10. The horizontal lines indicate the median values for each group at the indicated timepoints. PBMC: peripheral blood mononuclear cells; PMA: phorbol myristate acetate; LPS: lipopolysaccharide; HC: healthy controls.
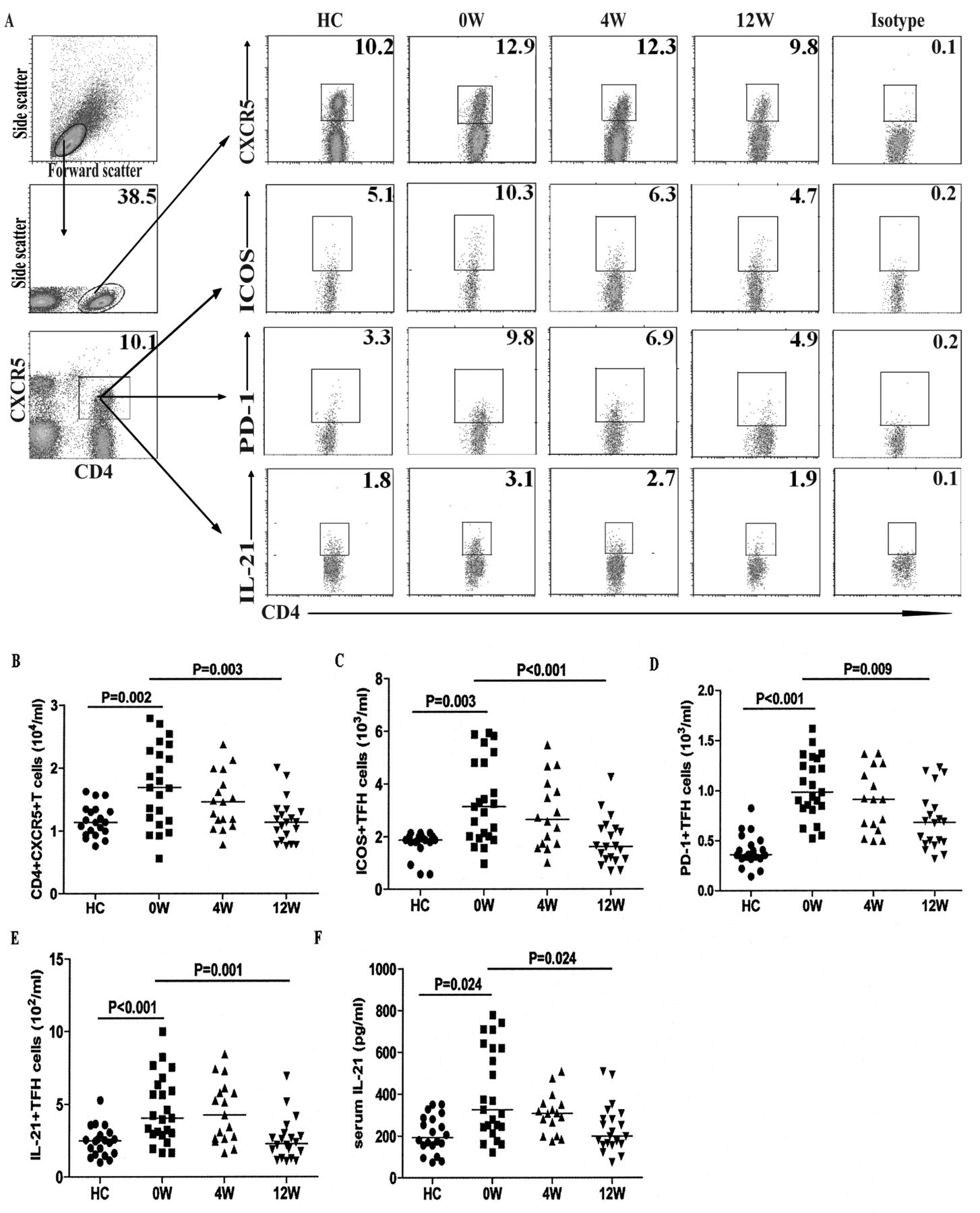
Increased numbers of circulating TFH-like cells in patients with new-onset SLE
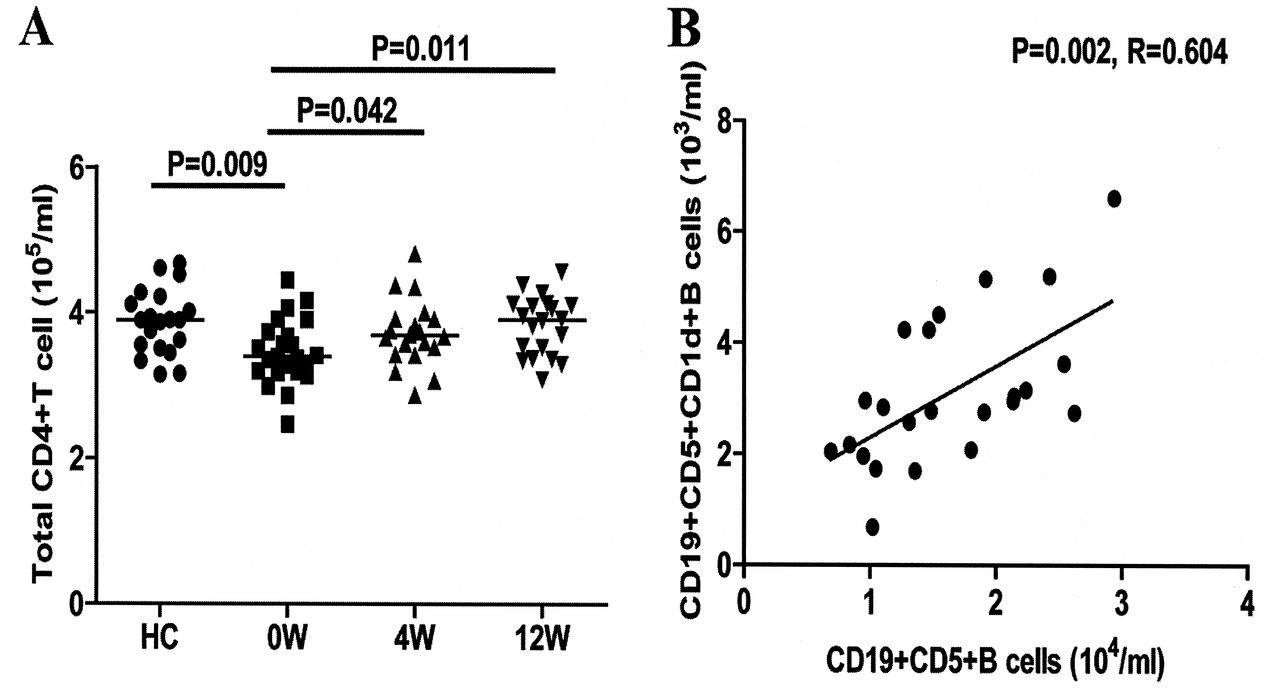
TFH cells are important regulators of humoral responses. Next, we characterized the numbers of circulating CD4+ and CD4+CXCR5+ TFH-like cells by flow cytometry. Our results revealed that there was a significant difference in the numbers of CD4+ and different subsets of CD4+CXCR5+ TFH-like cells, as well as in the levels of serum IL-21 among the different groups of subjects (p < 0.05, determined by Kruskal-Wallis test and posthoc Bonferroni correction test; Appendix 1 and Figure 4). The numbers of circulating CD4+ T cells in the patients with new onset SLE were significantly lower than those in the HC (Appendix 1). However, the numbers of CD4+CXCR5+, CD4+CXCR5+ICOS+, CD4+CXCR5+PD-1+, and CD4+CXCR5+IL-21+ TFH-like cells in the patients were significantly greater than those in the HC (Figure 4B, C, D, E). Further analysis revealed that the concentrations of serum IL-21 in the patients were significantly higher than those in the HC (Figure 4F). The concentrations of serum IL-21 were correlated positively with the numbers of CD4+CXCR5+ TFH-like cells (r = 0.647, p = 0.008). Moreover, the numbers of CD4+CXCR5+ or CD4+CXCR5+IL-21+ TFH-like cells were correlated positively with the numbers of CD19+ or CD27highCD19+ B cells in those patients (r = 0.573, p = 0.004; r = 0.491, p = 0.017), respectively. In addition, the concentrations of serum IL-21 were correlated positively with the numbers of CD27highCD19+ B cells in those patients (r = 0.552, p = 0.006). Following the anti-SLE treatment for 4 and 12 weeks, the numbers of CD4+CXCR5+, CD4+CXCR5+Bcl-6+, and CD4+CXCR5+IL-21+ TFH-like cells and the concentrations of serum IL-21 in those patients were gradually reduced, particularly at 12 weeks after the initial treatment. Similarly, the percentages of each type of TFH-like cell gradually decreased in the patients following treatment (Figure 4). These data suggest that increased numbers of circulating TFH-like cells and higher levels of serum IL-21 may be associated with the early process of SLE in Chinese patients.
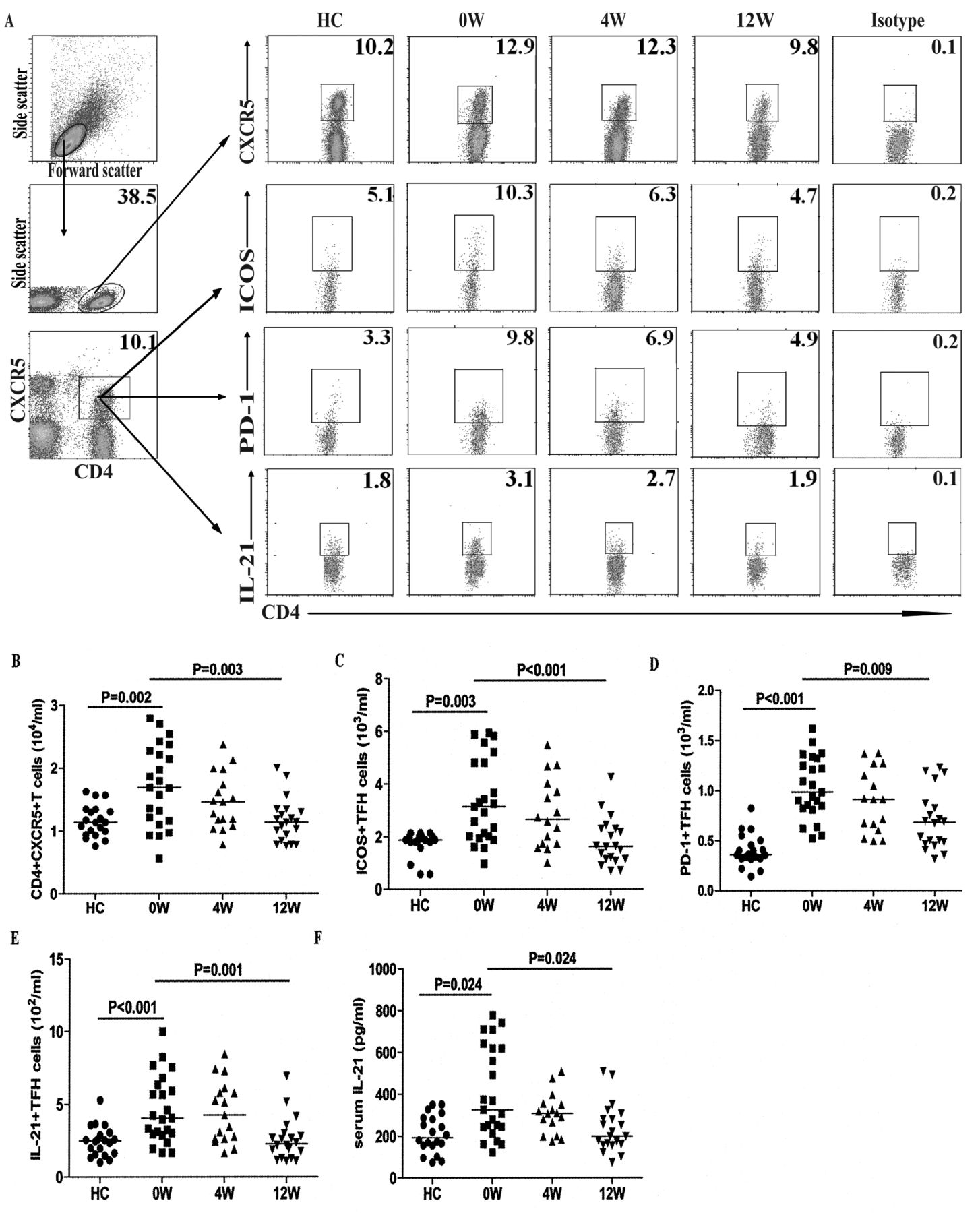
Analysis of peripheral blood TFH-like cells in individual subjects. PBMC were isolated from individual subjects at the indicated timepoints and stimulated with PMA/ionomycin in vitro. The cells were stained with PerCP-anti-CXCR5 and APC-anti-CD4 for 30 min, fixed, and permeabilized, followed by intracellular staining with PE-anti-Bcl6 or Alexa Fluor-anti-IL-21. Additional cells were stained with FITC-antiinducible T cell costimulator, phycoerythrin-anti-PD-1, PerCP-anti-CXCR5, and APC-anti-CD4 for 30 min. The numbers of peripheral blood CD4+CXCR5+, CD4+CXCR5+ICOS+, CD4+CXCR5+PD-1+, and CD4+CXCR5+IL-21+ TFH-like cells in individual subjects were determined by flow cytometry. The concentrations of serum IL-21 were determined by ELISA. Data shown are representative FACS charts or the mean numbers of each type of cells per ml of peripheral blood in individual subjects from 2 separate experiments. A. Representative FACS charts. B, C, D, E. Numbers of CXCR5+CD4+, ICOS+CXCR5+CD4+, PD-1+CXCR5+CD4+, and IL-21+CXCR5+CD4+ TFH-like cells. F. Levels of serum IL-21. The horizontal lines indicate the median values for each group at the indicated timepoints. TFH: follicular helper T cells; PBMC: peripheral blood mononuclear cells; PMA: phorbol myristate acetate; APC: antigen-presenting cell; HC: healthy controls.
The relationship between the numbers of B or TFH-like cells and the values of clinical measures in patients with SLE
We found that the numbers of CD27highCD19+ B cells were correlated positively with the values of SLEDAI and the concentrations of serum IL-21 in the patients with new-onset SLE (Figure 5). However, there was no significant association of the numbers of other types of B cells with any of the clinical measures tested in this population (data not shown). Further, the concentrations of serum IL-10 were correlated negatively with the values of SLEDAI, serum C3, CRP, and ESR in those patients (Figure 5). In addition, the concentrations of serum IL-21 were correlated positively with the values of SLEDAI, serum C3, and ESR in those patients (Figure 5), while there was no significant association between the concentrations of serum IL-21 or IL-10 and other measures tested in those patients (data not shown). These data suggest that the concentrations of serum IL-10 and IL-21 may be valuable markers for the evaluation of disease activity in patients with new-onset SLE.
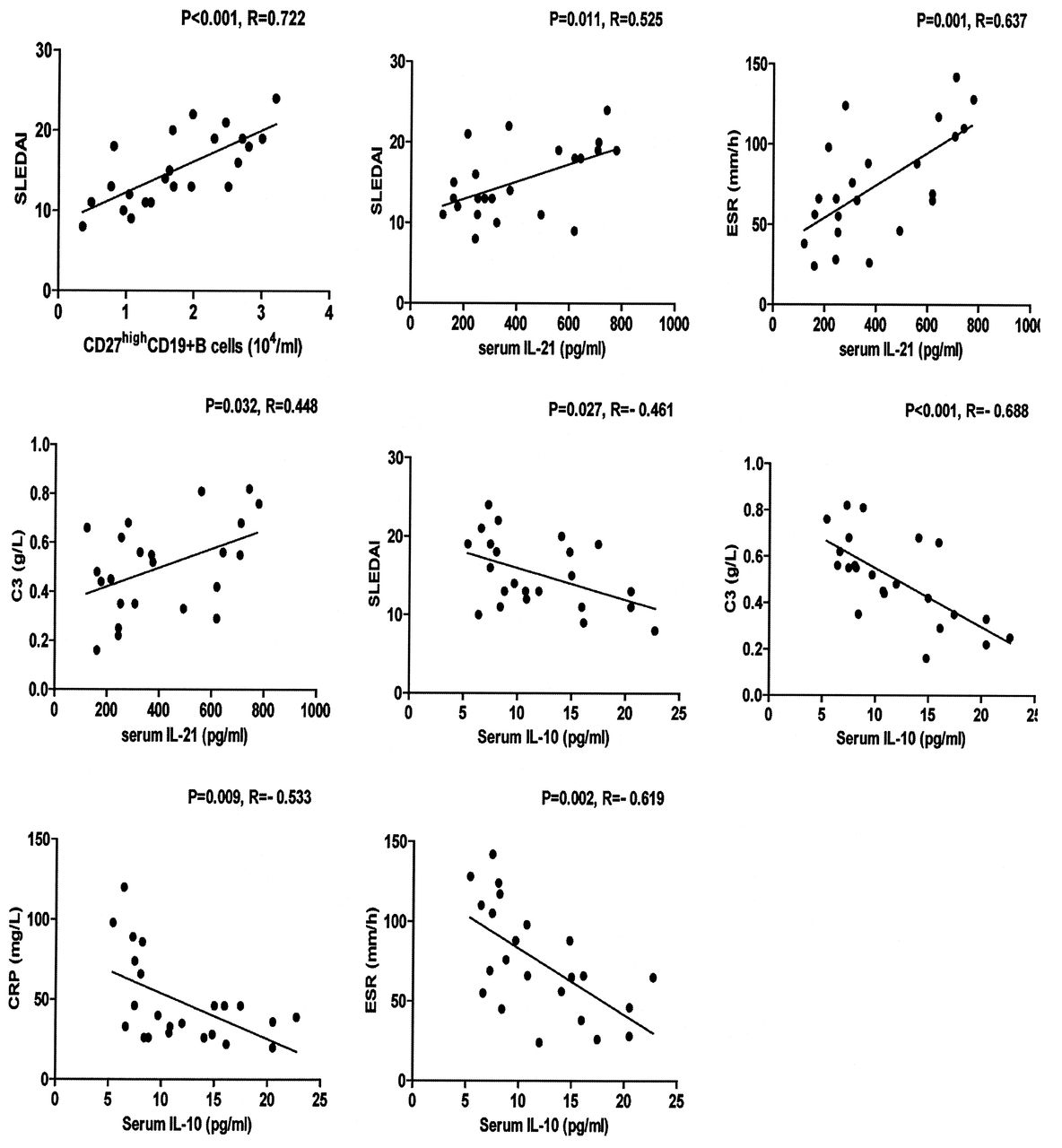
Correlation analysis. The potential association between the numbers of CD19+CD27high B cells, the levels of serum interleukin 10 (IL-10) or IL-21, the values of Systemic Lupus Erythematosus Disease Activity Index (SLEDAI) and erythrocyte sedimentation rate (ESR), and the levels of serum complement level 3 (C3) and C-reactive protein (CRP) in those patients before the anti-SLE therapy were analyzed by the Spearman’s rank correlation test. Data shown are the values of individual patients (n = 23).
DISCUSSION
B cells play a pivotal role in the initiation and perpetuation of autoimmune diseases8,27. Activated autoreactive B cells not only produce autoantibodies, but also secrete cytokines and express surface molecules, such as CD86 and CD95, that present antigen and regulate T cell immunity28. In our study, we examined the numbers of different subsets of B cells in new- onset SLE patients and HC. We found that the numbers of CD19+ B cells were significantly lower than those in the HC, consistent with previous observations29,30. Further, the numbers of CD19+CD27high and CD19+CD38+ but not CD19+CD27low B cells in the patients were significantly greater than those in the HC. It is possible that some CD19+CD27low B cells were converted into CD19+CD27high and CD19+CD38+ B cells during the early process of SLE6,31. In addition, the numbers of CD86+ activated, CD95+ exhausted, CD86+CD38+, CD95+CD27+, and CD95+CD27−, but not CD86+CD38− B cells in the patients were significantly greater than those in the HC. The increased numbers of CD86+ and CD95+ B cells may reflect an enhanced antigen-presenting activity and an increased sensitivity of those B cells to apoptosis induction, respectively32,33. Moreover, we observed a similar pattern of changes in the percentages of different subsets of B cells in patients with SLE. Together, these data indicate that an imbalance of different subsets of B cells exists in the patients with new-onset SLE.
Studies have shown that CD19+CD5+CD1d+ B cells secrete IL-10 and negatively regulate immune responses and autoimmune diseases12,13,14,15. We found that the numbers and percentages of CD19+CD5+, CD19+CD5+CD1d+, and CD19+CD5+CD1d+IL-10+ B cells in the patients were significantly lower than those in the HC. Similarly, the concentrations of serum IL-10 in the patients were significantly lower than those in the HC. Our data were consistent with the findings from a mouse model of SLE-like disease14, but were different from another report34. These conflicting results may stem from the difference of the enrolled patients. While our study centered on new-onset patients, their study included patients receiving immunosuppressants and at varying stages of disease. Further, the numbers of CD19+CD5+, CD19+CD5+CD1d+, and CD19+CD5+CD1d+IL-10+ B cells were positively correlated in those patients, supporting the notion that IL-10+ B cells are derived from B1 cells14. Moreover, the concentrations of serum IL-10 or the numbers of CD19+CD5+CD1d+IL-10+ B cells were correlated negatively with the numbers of CD27highCD19+ B cells in those patients. In addition, the values of SLEDAI were correlated positively with the numbers of CD27highCD19+ B cells, but negatively with the levels of serum IL-10 or the numbers of CD19+CD5+CD1d+IL-10+ B cells in those patients. These data suggest that IL-10+ B cells may negatively regulate antigen-specific B cell activation during the process of SLE18. Therefore, greater numbers of CD27highCD19+ B cells and reduced numbers of IL-10+ B cells as well as decreased levels of serum IL-10 may contribute to the early process of SLE in Chinese patients. To our knowledge, this is the first report about the imbalance of different subsets of B cells in patients with new-onset SLE.
TFH cells regulate germinal center formation and B cell responses as well as autoimmune diseases21,35. TFH cells express the CXCR5 chemokine receptor and secrete IL-2117,18,19,31,36,37,38. We examined the numbers and frequency of different types of TFH cells and found that the numbers of CD4+CXCR5+ and CD4+CXCR5+IL-21+ TFH-like cells in the patients were significantly greater than those in the HC. Similarly, the numbers of ICOS+ and PD-1+ TFH-like cells in the patients were also significantly greater than those in the HC. Our recent study has shown that CD4+CXCR5+Foxp3+ TF receptor cells exist in patients with SLE39. Given that CD4+CXCR5+Foxp3+ TF receptor cells have been demonstrated to negatively regulate inflammation, the increased numbers of TFH-like cells in patients with SLE may not only positively regulate humoral responses, but also provide negative signals to prevent aberrant B cell activation during the early process of SLE38,40. Further, the concentrations of serum IL-21 in the patients were significantly higher than those in the HC. In addition, the concentrations of serum IL-21 were correlated positively with the numbers of CD4+CXCR5+ TFH-like cells, suggesting that IL-21 is predominantly produced by TFH-like cells23,24. Moreover, the numbers of TFH-like cells and the levels of serum IL-21 were correlated positively with the numbers of CD27highCD19+ B cells in those patients, respectively. These data indicate that TFH-like cells and IL-21 are key regulators of functional B cell development18,41,42. More importantly, the levels of serum IL-21 were correlated positively with the values of SLEDAI, ESR, and the levels of serum C3 in those patients. Collectively, our data suggest that increased numbers of TFH-like cells and increased levels of IL-21 may be associated with the pathogenesis of SLE and that the levels of serum IL-21 may be a valuable marker for the evaluation of SLE activity in Chinese patients. Conceivably, TFH-like cells may be targets for the design of new immunotherapies for SLE in Chinese patients.
Anti-SLE therapy has been demonstrated to effectively control symptoms in patients with SLE in the clinic. We found that the anti-SLE therapy modulated the imbalance of different subsets of B cells by reducing the numbers of CD19+CD38+, CD19+CD27high, CD19+86+, and CD19+CD95+ B cells, but increasing the numbers of IL-10+ B cells and the levels of serum IL-10 in those patients. Similarly, the anti-SLE therapy also reduced the numbers of different types of TFH-like cells and the levels of serum IL-21 in those patients, which were consistent with previous findings43. Given that prednisone, MTX, and other immunosuppressants have been demonstrated to affect the activation and function of immunocompetent cells, it is possible that these drugs directly affect the activation and proliferation of B and TFH-like cells. Alternatively, these drugs may affect the bone marrow and the development of B and TFH-like cells, leading to the changes in the numbers and frequency of circulating B and TFH-like cells in the patients.
Although the significant changes in the numbers of different subsets of B and TFH-like cells may contribute to the therapeutic effect, we cannot completely exclude that other anti-SLE treatment-related effects contribute to improvement of clinical symptoms in these patients. Moreover, our findings were in disagreement with a previous report35. The difference between our findings and those of that group may be due to several factors, including the varying criteria for characterizing TFH-like cells, followup periods, and immunosuppression regimens.
Thus, our findings may provide new insights into the mechanisms underlying the therapeutic efficacy of anti-SLE therapy in patients with SLE. We are interested in further investigating how these drugs modulate the numbers and frequency of these cells in patients with SLE.
This was the first study, to our knowledge, to comprehensively analyze the numbers of different subsets of B and TFH-like cells in patients with new-onset SLE. Our data suggest that increased numbers of activated B cells and TFH-like cells and decreased numbers of IL-10+ B cells may be associated with the early process of SLE in Chinese patients. Anti-SLE therapy not only effectively improved clinical symptoms, but also modulated the imbalance of different subsets of B cells and TFH-like cells. Therefore, our findings may provide new insights into the pathogenesis of SLE and aid in the design of new immunotherapies for the intervention of SLE in the clinic.
We recognized that our study had limitations, including a small sample size and the lack of convincing markers and functional studies of different subsets of B and TFH-like cells. Thus, further validation of these findings in a bigger population is warranted.
Acknowledgment
We thank Medjaden Bioscience Limited for assisting in the preparation of this manuscript.
APPENDIX 1.
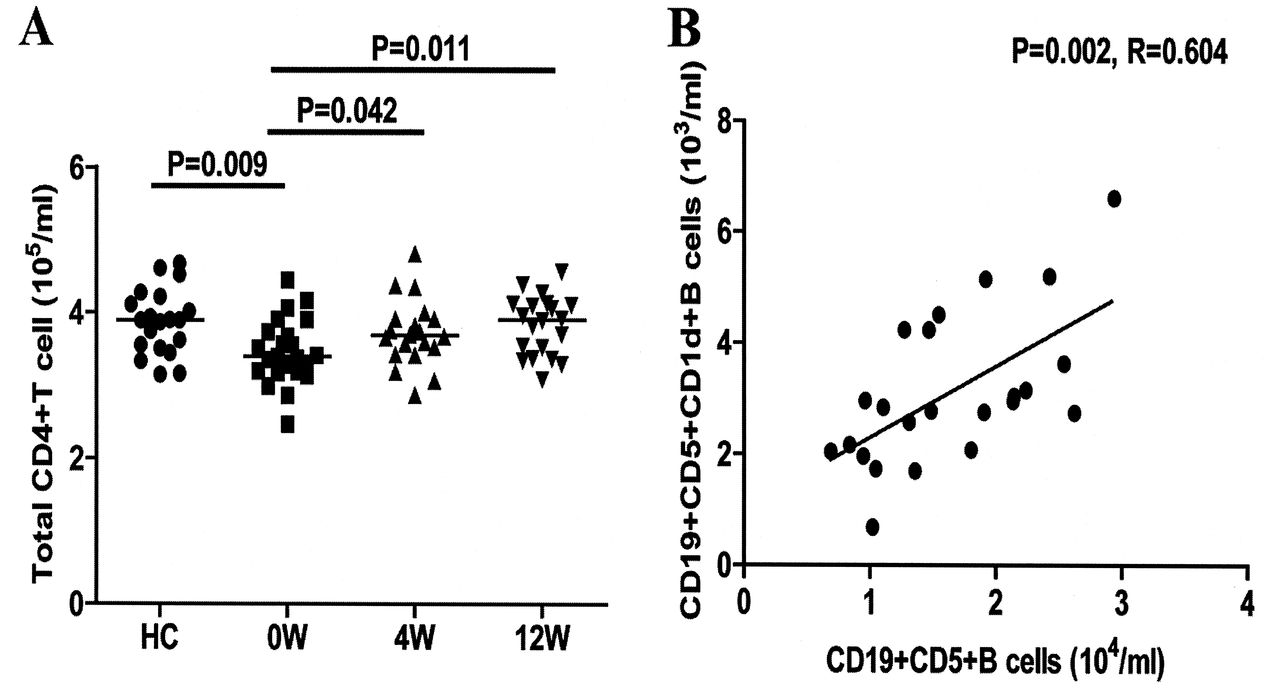
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
A. Total CD4+ T cells. B. Correlation between the numbers of CD19+CD5+ and CD19+CD5+CD1d+ B cells in patients with new onset SLE. Data are values of individual patients at indicated timepoints. Horizontal lines indicate median values for each group at indicated timepoints. SLE: systemic lupus erythematosus; HC: healthy controls.
Footnotes
-
Supported by grants from the National Natural Science Foundation of China (No. 30972610 and 81273240), Jilin Province Science and Technology Agency (No.20110716), the Health Department Research Projects in Jilin Province (2009Z054), and the Norman Bethune Program of Jilin University (2012206).
- Accepted for publication April 4, 2014.
REFERENCES
- 1.
- 2.
- 3.
- 4.
- 5.
- 6.
- 7.
- 8.
- 9.
- 10.
- 11.
- 12.
- 13.
- 14.
- 15.
- 16.
- 17.
- 18.
- 19.
- 20.
- 21.
- 22.
- 23.
- 24.
- 25.
- 26.
- 27.
- 28.
- 29.
- 30.
- 31.
- 32.
- 33.
- 34.
- 35.
- 36.
- 37.
- 38.
- 39.
- 40.
- 41.
- 42.
- 43.