

Abstract
Objective. Knee flexion contractures (KFC) are limitations in the ability to fully extend the knee joint. In people with knee osteoarthritis (OA), KFC are common, impair function, and worsen outcomes after arthroplasty. In KFC, the posterior knee capsule is believed to play a key role, but the pathophysiology remains poorly understood. We sought to identify gene expression differences in the posterior knee capsule of patients with OA with and without KFC.
Methods. Capsule tissue was obtained from the knees of 12 subjects diagnosed with advanced-stage OA at the time of knee arthroplasty surgery. The presence or absence of KFC allocated patients into 2 groups using a case-control design. Genomewide capsular gene expression was compared between the 2 patient groups. Confirmation of differential expression of the corresponding proteins was performed by immunohistochemistry on tissue sections.
Results. There were no significant demographic differences between the patients with OA with KFC and without KFC save for reduced extension in their surgical knee (p < 0.01). KFC patients showed a 6.4-fold decrease in CSN1S1 (p = 0.017) gene expression and a 3.7-, 2.0-, and 2.6-fold increase in CHAD, Sox9, and Cyr61 gene expression, respectively (p = 0.001, 0.004, 0.001, respectively). There were corresponding increases in protein levels for chondroadherin, sex determining region Y-box 9, and casein alphaS1 (all p < 0.05). Functional analysis of the differentially expressed genes indicated a strong association with pathways related to the extracellular matrix and to tissue fibrosis.
Conclusion. Posterior capsules in endstage OA knees with KFC exhibited differential expression of 4 genes all previously documented to be associated with tissue fibrosis.
The knee is one of the joints most commonly affected by osteoarthritis (OA), and about 15% of the population in the United States over 65 years old has symptoms of knee OA1. Pain, stiffness, and loss of function are common symptoms, making knee OA a leading cause of disability1,2. Knee flexion contractures (KFC) are limitations in full passive knee extension3. KFC compound the symptoms of OA by disturbing gait, increasing energy expenditure, and decreasing function3,4. In a group of patients with knee OA representing 5228 knees, more than one-third lacked full extension when presenting for primary total knee arthroplasty (TKA)2. Partial or total TKA is performed to treat endstage knee OA and represents a successful procedure conducted on over 719,000 cases in 2010 in the United States alone5. Over 4.5 million persons are currently living with at least 1 TKA, 95% of whom report satisfaction with their procedure6.
For the 5% of patients with knee OA not cured by TKA, mechanical problems related to KFC, the implant, thrombosis, and infection are the most commonly diagnosed reasons and often necessitate revision surgery2,7. KFC after primary TKA results in poorer outcomes in terms of pain, knee score, and overall satisfaction2. Preoperative KFC is a risk factor for KFC postarthroplasty: 62% of TKA cases with preoperative KFC ≥ 20º did not achieve complete contracture correction at 1 year8. In surveys administered to large cohorts of patients with arthritis, participants identified lack of complete joint mobility as their second most troublesome problem after pain9. Consistently, the second preference for improvement of health for arthritic patients was mobility, with emphasis on walking and handling activities9.
While knowledge about the degenerative processes taking place in the articular cartilage of patients with OA is abundant10, limited information is available regarding the contribution of other knee joint tissues, including the knee capsule, to the pathophysiology of knee OA. When a KFC accompanies knee OA, partial release of the posterior joint capsule limited to the lateral area, as well as release of the lateral ligaments during TKA, improves knee range of motion (ROM)11. Animal joint contracture models, with and without intraarticular damage, provide more direct evidence for the contribution of the posterior knee capsule to KFC. Immobilization of rat knees in flexion over extended periods of time caused the posterior capsule to shorten and become fibrotic, resulting in a KFC3. Full extension was not regained after complete sectioning of transarticular muscles and the limitation knee ROM was attributed to the posterior knee capsule. Unfortunately, sectioning of the posterior knee capsule is not a routine option for patients with knee OA with flexion contractures owing to the risks for joint instability and neurovascular injury. Nonsurgical treatments must therefore be explored.
The goal of our study was to identify a gene expression signature in the posterior capsule of patients with knee OA with KFC. Our objectives were to (1) report changes in the collagenous composition of the posterior joint capsule, (2) identify differentially expressed genes and submit those genes for functional analysis, and (3) confirm corresponding protein product levels for those genes in posterior joint capsules from OA knees with KFC.
MATERIALS AND METHODS
Patient recruitment and data collection
Subjects were prospectively recruited prior to TKA for severe primary knee OA at The Ottawa Hospital, Ottawa, Ontario, Canada. Subjects met the American College of Rheumatology criteria for knee OA12. Exclusion criteria included history of inflammatory arthropathy, hemophilia, cancer treatment, surgery, arthroscopy, significant trauma to the operative knee, or known genetic anomaly. Risk factors for preoperative KFC in patients with OA are not known; therefore selection of recorded demographic data was based on previous studies for knee OA2,11. ROM of both the surgery and contralateral knees were recorded in degrees (Table 1). Based on these demographic features, we matched contracture to noncontracture subjects as closely as possible using a case-control design.
Demographic features of patients whose samples were selected for microarray. Data shown are mean or percentage ± SD.
All patients gave written informed consent to participate. Our study was approved by the institutional research ethics board.
Knee range of motion
Knee ROM was measured with a goniometer by 2 research assistants trained by a physician using a method with good interrater reliability13. Patients were placed supine, hips at neutral, with a rolled towel under the calcaneus to maximize knee extension. The goniometer fulcrum was centered over the lateral femoral condyle, and the goniometer arms were aligned with the greater trochanter and fibular head, respectively. The angle formed by these landmarks was the maximal angle of extension. A lack of 180° knee extension of ≥ 6º was considered a KFC2,13, and has been shown to bring about quadriceps demand while standing4.
Radiographic severity of OA and knee alignment
Radiographic severity was graded independently by 1 investigator (TMC) and a radiologist (MQ), blinded to the patient’s identity and clinical information, prior to obtaining gene expression data using the Kellgren and Lawrence criteria14. Disagreements were resolved by consensus.
Knee alignment was determined on standing knee radiographs using the anatomic axis method as described15. Varus alignment was < 180° and valgus alignment > 180°.
Capsule collection and storage
During TKA, posterolateral and/or the posteromedial knee capsule was collected according to surgical preference. We refer to this tissue as “posterior capsule”. The posterior capsule sample (about 0.5 cm ×1 cm ×1 cm) was immersed in RNAlater solution in the operating room for transportation to the investigators’ laboratory. The capsule was divided into pieces for histology/immunohistochemistry (immersed in 10% buffered formalin) and for RNA extraction (stored in RNAlater solution for 24 h at 4°C before transfer to storage at −80°C). The tissue used for histology and immunohistochemistry was adjacent to that used for RNA extraction.
Histology processing and analysis
We characterized the posterior knee capsule tissue of the 12 patients whose tissue samples were used for RNA extraction. Posterior capsules were paraffinized, cut into 5-μm sections, and stained with Masson trichrome. Digitized microscope color images were analyzed for collagen cross-sectional area using ImageJ (version 1.46, US National Institutes of Health). The Threshold color plugin isolated blue color on the original color images, representing collagen. Images were then converted to 8-bit images, and the threshold function was used to determine collagen area. To calculate percent collagen relative to total area, total area was calculated by including the entire color spectrum on the original color images. For all histologic analyses, the analyzer was blinded to patient identity and clinical information.
Total RNA isolation
Capsule samples for RNA isolation were homogenized using a mortar and pestle, and submerged in TRIzol reagent (Invitrogen/Life Technologies Corp.) using the protocol provided by the manufacturer. RNeasy mini kit columns (Qiagen) were used to remove non-RNA contaminants according to the manufacturer’s instructions. The integrity of the RNA sample was determined using the Experion RNA StdSens Analysis Kit (Bio-Rad) to produce an RNA quality index (RQI). The mean RQI was 8.5.
Microarray hybridization
Genomewide RNA expression was determined using the HumanHT-12 v4 BeadChip (Illumina Inc.). Six subjects with and 6 without contracture were selected based on the 12 arrays available on a single chip, eliminating interchip variability. The hybridization workflow was initiated as described by the manufacturer. The RNA first- and second-strand synthesis step was performed using the TotalPrep RNA amplification kit (Illumina Inc.).
Microarray data analysis
Preprocessing of probe-level data and identification of differentially expressed genes were performed using R statistics software (v.3.0.3). The “lumi” package16 was used in its standard settings in the R environment for background correction, variance stabilization transformation, and quantile normalization across all 12 arrays. Probes with an increased or decreased fold-change of 1.5 and a p value of ≤ 0.05 were considered significant. The empirical base (Wright and Simon) method was used for between-group gene expression comparison17.
We performed functional analysis of differentially expressed genes using the online tool Database for Annotation, Visualization and Integrated Discovery (DAVID, v6.7). Functional annotations were clustered by GO Terms and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways. Cluster selection was based on p values. The selection of candidates for validation was based on the results of the functional analysis and biological terms with the lowest p values. Candidate proteins for validation included chondroadherin (CHAD), sex determining region Y-box 9 (Sox9), casein alphaS1 (CSN1S1), and cysteine-rich angiogenic inducer, 61 (Cyr61).
Levels of protein product by immunohistochemistry
Primary antibodies to CHAD (HPA018241), Sox9 (HPA001758), CSN1S1 (HPA035659), and Cyr61 (HPA029853) were purchased from the Prestige Antibodies collection of polyclonal, monospecific, affinity-isolated rabbit antihuman antibodies (Sigma-Aldrich Canada Co.).
Five-µm posterior knee capsule slides were deparaffinized to water, and heat-induced epitope retrieval (HIER) was performed when needed. Endogenous peroxidases were blocked (Peroxidazed 1, Biocare Medical LLC), and then samples were background-blocked (Background Sniper, Biocare Medical) and incubated with primary antibody. CHAD primary antibody was used at a dilution of 1:300 overnight at room temperature (RT). Primary anti-Sox9 was incubated overnight at a dilution of 1:125 at RT. Primary anti-CSN1S1 was incubated overnight at 1:250 at 4°C. Primary anti-Cyr61 was incubated overnight at a dilution of 1:20 at RT. For Sox9, the HIER method was performed at pH 6 and 110°C for 5 min. For Cyr61, HIER was performed at pH 6, 100°C for 10 min. HIER was not required for CHAD or CSN1S1.
Secondary antibodies were a 2-step anti-rabbit horseradish peroxidase micropolymer conjugate (Mach 3, Biocare Medical). Slides were developed with diaminobenzidine (DAB) chromogen. The slides were counterstained using Shandon instant hematoxylin. For negative controls, we omitted the primary antibody as recommended by the manufacturer18 and as described by others19.
Immunohistochemistry microscopy
Semiquantitative analysis of DAB staining intensity reliably detects relative protein concentration20. Cellular and extracellular matrix (ECM) staining to CHAD, Sox9, CSN1S1, and Cyr61 were assessed under light microscopy at 50× magnification in 4 randomly selected areas of each slide. The number and percentage of cells positive for each antibody were counted manually, and cell type, subcellular location (nuclear or cytoplasmic), and ECM staining intensity were recorded (none = 0, mild = 1, moderate = 2, or intense = 3). Should 1 randomly selected area present no cell or no stained cell, this would affect the average of the 4 areas. We, therefore, also reported the percentage of stained cells only at areas containing stained cells. The analyzer was blinded to sample identification for immunohistochemistry analysis.
Data analysis and statistics
Statistical testing was performed using SPSS 16.0 (SPSS). For demographics, histology, and immunohistochemistry, we used the nonparametric Mann-Whitney U test or Kruskall-Wallis test where appropriate. Differences between contracture and noncontracture groups were considered statistically significantly at a p value of ≤ 0.05.
RESULTS
Demographics of patients and posterior capsule histology
Table 1 shows the demographics of the 12 patients with OA selected for microarray, 6 with a KFC and 6 without, matched according to their demographic features. The mean limitation to full extension was 11.3° ± 3.9° in the contracture group and 1.0° ± 1.1° in the noncontracture group. Knee joints contralateral to a surgical knee with KFC had less range of extension (lacking 5.7° ± 5.0°) than knee joints contralateral to a surgical knee without contracture (lacking 1.0° ± 1.1°, p = 0.09). There were no statistically significant differences between the groups for any of the other demographic features (Table 1).
Cross-sectional area of the posterior capsules of the KFC group for collagenous tissue was 64.6% (SD ± 15.8%), compared to the noncontracture group, which was 54.4% (SD ± 19.0%), p = 0.20 (Table 1, Figure 1A, left and right, respectively).
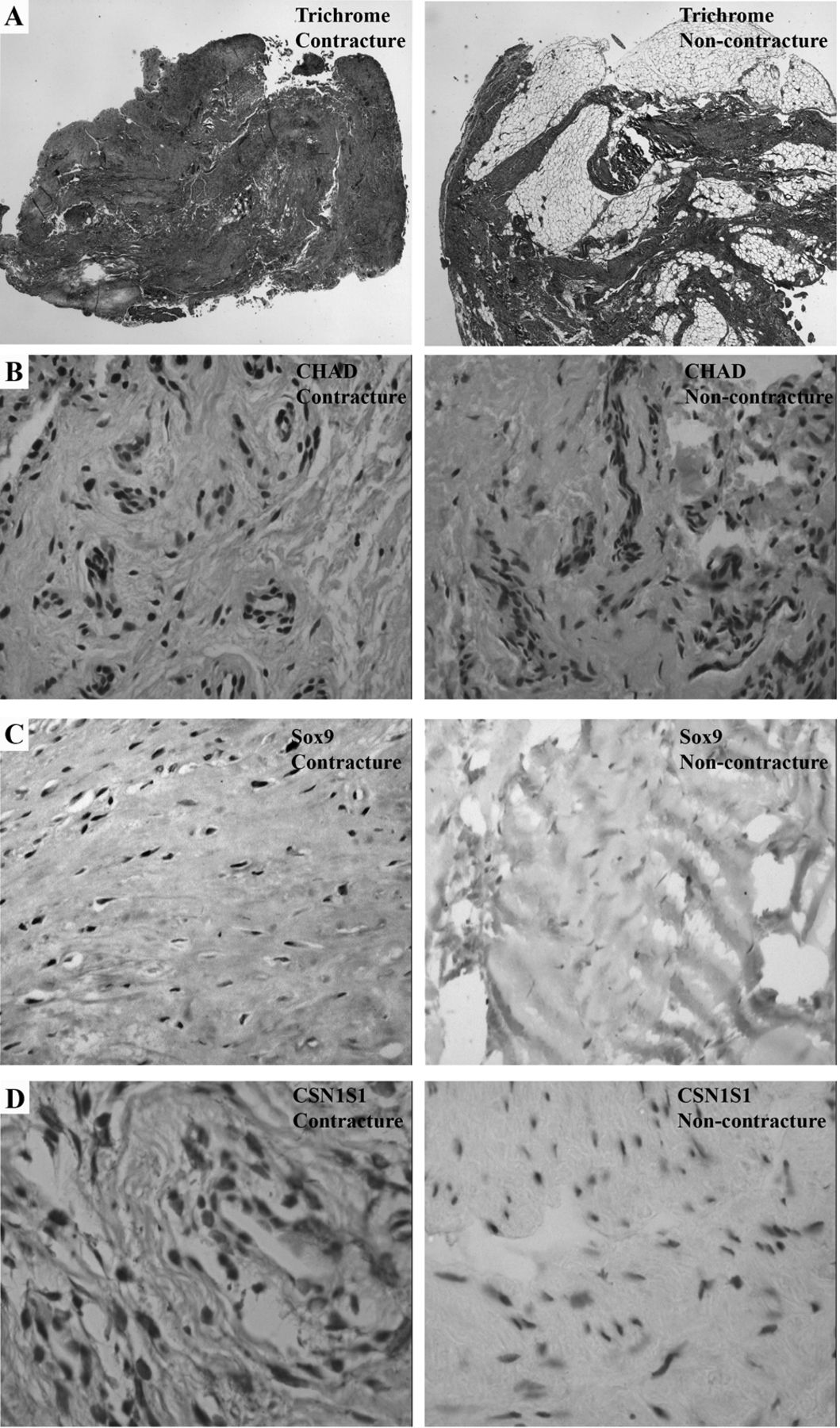
Histology and immunohistochemistry of OA posterior knee capsule from contracture and noncontracture subjects. Left column images: contracture samples. Right column images: noncontracture sample. Areas of lighter shade represent the presence of collagen. A. Trichrome stained samples 20× original magnification. B. CHAD immunohistochemistry, 50× original magnification. C. Sox9 immunohistochemistry, 50× original magnification. D. CSN1S1 immunohistochemistry, 20× original magnification. All immunohistochemistry counterstaining was done using hematoxylin. OA: osteoarthritis; CHAD: chondroadherin; Sox9: sex determining region Y-box 9; CSN1S1: casein alphaS1.
Microarray data analysis
Our initial data analysis revealed 65 differentially expressed genes, 30 of which were represented in our functional analysis (Table 2). Terms with the most counts from the 2 Gene Ontology project biological processes included cell adhesion and biological adhesion. Several cellular component terms were identified relating to ECM homeostasis. A KEGG pathway of ECM-receptor interaction was discovered. While the transforming growth factor-β (TGF-β) pathway was listed, it did not reach statistical significance (p = 0.07; Table 2).
Functional analysis of differentially expressed genes. Differentially expressed genes not represented: ADARB1, ALDH1A1, ALDH1A2, ALDH1A3, AOX1, BIN1, C17orf47, CHST6, CLIC5, CRYAB, DLX5, DOCK9-AS2, ERRFI1, FAM118A, GEM, HIST1H4C, KCNJ6, KYNU, LOC729254, MKX, MT1X, MYO1D, PFKFB3, PPP1R3C, RNF150, SCARA5, SNHG5, TCEAL2, TMEM176B, TMEM98, TSC22D1, TSHZ2, UAP1, ZFP36L2, ZNF385B.
Based on our functional analysis and previous joint contracture findings in settings other than OA21,22, we hypothesized that KFC in OA could involve fibrosis of the joint capsule. We therefore verified the protein expression of 4 genes from our list that have been implicated in ECM alteration and fibrosis: CHAD, Sox9, CSN1S1, and Cyr61 (Table 3).
Statistically significant genes selected from functional analysis.
Tissue levels of gene protein products by immunohistochemistry
Proteins are the functional posttranslational products of mRNA, and quantitative analysis of mRNA using methods such as quantitative real-time PCR (qRT-PCR) correlates poorly with protein product23,24. We therefore confirmed differential microarray gene expression by examining corresponding protein products using immunohistochemistry. We found a higher percentage of CHAD-stained cells in fields with stained cells for the contracture group compared to the noncontracture group (p ≤ 0.05; Table 4, Figure 1B). There was no difference in overall cellularity (cells/field) or CHAD-staining intensity between the 2 groups (p > 0.05). Capsules with KFC showed higher cellular staining intensity for Sox9 than did samples without contracture (p ≤ 0.05; Table 4, Figure 1C), but there was no difference between the 2 groups in the percentage of Sox9-positive cells. For CSN1S1, ECM staining was increased in the contracture group (p < 0.001), contrary to the decreased mRNA expression seen in microarray (Table 4, Figure 1D). There were no statistically significant between-group differences for Cyr61 (Table 4).
Average protein expression of CHAD, Sox9, and Cyr61 measured by immunohistochemistry in the contracture and noncontracture groups. Data in bold face are statistically significant.
DISCUSSION
We found that the posterior capsule of OA knees with KFC demonstrated significantly decreased expression of CSN1S1 and increased expression of CHAD, Sox9, and Cyr61 compared to non-KFC specimens. Increased protein for CHAD, Sox9, and CSN1S1 were confirmed by immunohistochemistry.
CHAD expression was 3.7-fold greater in the contracture group. CHAD is a member of the small leucine-rich repeat proteoglycan (SLRP) family25. Its RNA and protein have been found in many tissues, including cartilage, where it is important for chondrocyte adhesion25. SLRP are essential for collagen matrix architecture and ECM organization26. CHAD may be important for ECM remodeling as it binds to collagen types I and III, and OA synovial fluid fibroblasts26. TGF-β was shown to upregulate CHAD mRNA expression when cultured with bone marrow-isolated mesenchymal stem cells, and this was associated with chondrogenesis27. Other SLRP, including biglycan, decorin, and fibromodulin, are essential for type I collagen architecture28. Mice with disrupted SLRP function develop osteoporosis or osteogenesis imperfecta29. In a human model, expression of SLRP was increased in diabetic fibrotic nephropathy characterized by type I collagen deposition30. Type I collagen makes up > 80% of the collagen in joint capsules31. The overexpression of CHAD in our present study may be linked to a fibrotic joint capsule; however, it is unclear whether increased SLRP expression is the cause of or the response to increased fibrosis28. SLRP’s role in fibrosis may be through an interaction with TGF-β28.
Sox9 expression was increased 2-fold in contracture capsules. Sox9 is a member of the sex-determining region Y-box (Sox) transcription factors, is a transcription activator of collagen, matrilin-1, and aggrecan, and is essential in early chondrogenesis32. Viral-mediated Sox9 overexpression in cultured OA chondrocytes increased gene expression and protein synthesis of proteoglycans and type II collagen to wild-type levels32. Sox9 has been shown to interact with numerous ECM-related molecules including collagens I, II, and XI, and aggrecan33. Inappropriate Sox9 expression led to abnormal ECM deposition in mouse ureter, resulting in hydroureteronephrosis34. Sox9 was overexpressed in human keloids, suggesting a role in abnormal collagen proliferation35, and elevated Sox9 expression resulted in hepatic fibrosis through an osteopontin-dependent mechanism in both rat and human models36. Sox9 expression is regulated by numerous factors, including TGF-β37, and may therefore be a key player in capsular remodeling under conditions such as OA and contracture.
Cyr61 expression was increased 2.6-fold in the contracture group. Cyr61 (CCN1) is a member of the CCN group of proteins, which includes connective tissue growth factor (CCN2)38. This family of proteins is involved in numerous mechanisms, including ECM production and fibrosis38. CCN link cells with the ECM by binding to cellular adhesion molecules38. Constitutively elevated CCN2 expression is a hallmark of fibrosis, and overexpression of CCN2 initiates a potent fibrotic response39. CCN1 may also play a role in fibrosis. In a wound-healing model, treatment with CCN1 protein reduced fibrosis by driving fibroblasts into senescence, upregulated antifibrotic matrix metalloproteinases, and downregulated profibrotic Col1a1 and TGF-β140. Although not essential for liver function, Cyr61 accumulated in patients with liver cirrhosis41. Murine hepatic injury induced expression of Cyr61, triggering cellular senescence, limiting fibrogenesis, and promoting regression of liver fibrosis41. Cyr61 was found to attenuate TGF-β signaling, limiting liver fibrosis in a rat bile duct ligation model, indicating a possible therapeutic role for this protein. The elevated expression of Cyr61 in our patients with contracture may indicate an endogenous attempt at reducing the fibrotic process. In our samples, Cyr61 staining was mainly nuclear and cytoplasmic, possibly indicating inhibition of protein translation or export, precluding its preventive action and perhaps explaining why we did not detect a corresponding increase in Cyr61 protein; however, we did not examine this in our study.
CSN1S1 protein exerts a chaperone action to inhibit the stress-induced amorphous aggregation of client proteins42. CSN1S1 slows 1–40 β amyloid peptide fibrillogenesis by sequestering amyloid peptide on its surface through its chaperone-like activity42. Interestingly, the chaperone heat shock protein 47 has been shown to promote excess collagen secretion through interaction with procollagen and has been proposed as a therapeutic target for the prevention of fibrosis43, indicating that chaperones may have a role to play in this process. In our study, KFC was associated with reduced expression of CSN1S1, but increased expression of its protein product in the ECM. Extracellular CSN1S1 protein accumulation may represent a failed cellular attempt at controlling ECM proliferation, because CSN1S1 is unable to prevent excess collagen deposition. Increased ECM CSN1S1 concentration may trigger an intracellular negative feedback mechanism, inhibiting its own RNA production, but we did not test this in our study.
Our functional analysis identified biological and cellular adhesion as the most represented pathways, and differential expression of CHAD, Sox9, CSN1S1, and Cyr61 has been associated with fibrotic changes including cellular adhesion to ECM molecules26,33,38,42. These genes are members of families that influence fibrotic processes, and their protein products influence and/or are influenced by TGF-β, an important regulator of fibrosis37,44,45,46. Activated TGF-β is a potent stimulator of pulmonary and renal fibrosis, and its inhibition prevented fibrosis in these organs45,46. Our functional analysis also revealed the TGF-β pathway as a possible KEGG candidate, although not reaching statistical significance. Fibrosis has been identified in other models of KFC: immobilized rat knees showed increased deposition of type I and decreased expression of type III collagen compared to controls22. Joint capsules of patients with chronic postfracture elbow contracture showed increased expression of collagen and several tissue inhibitors of metalloproteinase21. Our histomorphometry results also showed more collagen content in contracture capsules, but this did not reach statistical significance. Overall, these results suggest a role for capsule fibrosis in KFC associated with OA.
OA is a leading cause of chronic pain and disability1. While knee OA can be successfully treated with TKA, the presence of KFC increases the chance of poor outcome and impaired function2. Identification of those at risk is thus essential. Microarray results have been used for diagnosis and prognosis for numerous diseases, including arthritis47,48,49. Our list of differentially expressed genes related to ECM adhesion and fibrosis appears to be distinct from those of previous studies48,49, which identified genes related to bone homeostasis and ECM degradation. This supports the notion that the gene signature we discovered is related to contracture in the setting of OA rather than OA alone. Detection of increased expression of CHAD, Sox9, CSN1S1, and Cyr61 may help identify patients with OA with a need for earlier, intense, monitored ROM exercises and physiotherapy to prevent KFC. Arthroscopic or intra-operative biopsy of posterior knee capsules could provide an accessible source for such screening; in fact, recent work has demonstrated that the peripheral blood may represent an easily accessible source of meaningful OA gene expression data48. Pharmacologic treatments may play a role in treating OA KFC and have already been successful in non-human models of renal and wound fibrosis40,46.
Limitations
Samples obtained at TKA represent endstage OA only and give limited information on contracture relative to OA progression. The sample size for our microarray was small; however, meaningful microarray data have been produced using 5 samples per group50, and by using only 1 chip, we eliminated interchip analysis variability. A larger sample size may have identified additional differentially expressed genes and pathways of OA KFC. Joint capsule is a multicellular tissue; therefore, the differentially expressed genes we discovered cannot be attributed to a single cell type. Nine of 12 microarray samples were from women, potentially limiting our results to women with severe OA. We confirmed differential expression using detection of protein product. Verification of the microarray results using qRT-PCR could have strengthened the transcriptional data. We did not harvest “normal” non-OA tissue to compare to OA tissue, and therefore do not comment on OA-specific differential gene expression.
Microarray analysis showed a decrease in CSN1S1 and an increase in CHAD, Sox9, and Cyr61 gene expression in posterior knee capsules of patients with OA with KFC compared to matched patients with OA without KFC. Significant increases in tissue protein levels were found for CHAD, Sox9, and CSN1S1. Each of these genes has been previously associated with fibrosis, suggesting that this may be 1 underlying mechanism leading to KFC in knee OA. Further research into knee capsule changes in OA can identify diagnostic and treatment options of patients with OA with KFC.
Acknowledgment
We thank Dr. Mathew Quon for assistance with radiography; Dr. Alain Stintzi, Dr. Zhara Montazeri, Dr. Julian Little, and Dr. Robert J. Feibel for project direction; Professor Philip Conaghan for manuscript advice; Louise Pelletier and the University of Ottawa Pathology Department, Elizabeth Coletta, Ying Nie Ping, Dr. Natalie Bunimov, and Génome Québec for technical support; Sarah Plamondon and Anna Fazekas for assistance with patient recruitment and ethics approval; Gloria Baker for assistance with manuscript preparation; and Dr. Geoffrey Dervin, Dr. Paul Kim, Dr. Peter Thurston, Dr. Robert J. Feibel, and Dr. Paul Edgar Beaulé for tissue collection.
Footnotes
-
Supported by Canadian Institutes of Health Research grant MOP97831 to Dr. Trudel and Dr. Laneuville.
- Accepted for publication July 10, 2014.








{kind=link}