

Abstract
Objective. To evaluate the expression of Dickkopf-1 protein factor (DKK-1), DKK-2, and β-catenin, components of the Wnt pathway, in human osteoarthritic (OA) and osteoporotic (OP) osteoblasts and to correlate it to cell metabolic activity, proliferation, and receptor activator of nuclear factor-κB ligand/osteoprotegerin (RANKL/OPG) expression.
Methods. Primary human osteoblast cultures were obtained from healthy, OA, and OP donors. In each cell population we evaluated DKK-1, DKK-2, nonphosphorylated β-catenin and RANKL/OPG expression, osteocalcin and alkaline phosphatase (ALP) synthesis, and cell proliferation, both in basal condition and after vitamin D3 stimulation.
Results. DKK-1 and DKK-2 showed opposite patterns of expression in OA and OP osteoblasts. The RANKL/OPG ratio was significantly higher in the OP group because of a greater expression of RANKL, whereas it was significantly lower in the OA group because of a higher expression of OPG. Treatment with vitamin D3 increased the RANKL/OPG ratio and DKK-2 expression and reduced DKK-1 expression in each cell population, but did not affect β-catenin levels. Both osteocalcin and ALP production and cell proliferation were enhanced in OA cells and reduced in the OP ones.
Conclusion. These data confirm that OA and OP are characterized by opposite bone changes, consisting of reduced bone remodeling processes with increased osteoblast activity in OA, and enhanced bone resorptive activity with reduction of osteoblast metabolism in OP, and suggest that the Wnt pathway is involved in the pathogenesis of both diseases.
Osteoporosis (OP) and osteoarthritis (OA) are 2 common diseases that represent a major cause of morbidity in the middle-aged and elderly population.
Whereas OP is characterized by the reduction of bone mass with a deterioration of bone microarchitecture that induces increased bone fragility and risk of fracture, in OA a generalized increase of bone density is observed, together with typical concomitant changes in subchondral and periarticular bone, such as eburnation of the joint surface, subchondral irregular thickness, sclerosis, and osteophyte formation1. These bone changes could be intimately related to the pathogenesis of cartilage degradation, leading to the initiation and progression of cartilage damage2 that is the main hallmark of OA. Whether bone changes initiate or are simply involved in the progression of cartilage loss remains a matter of debate3.
An inverse relationship between OP and OA bone alterations was noted 5 decades ago4,5, with the clinical observations of the absence of OA in the head of the femur excised during the treatment of fractures, the rarity of nontraumatic hip and spine fractures in OA subjects6, and the late appearance of OP fractures in OA subjects, suggesting that OA might have a protective effect on the progression of OP7. Several studies have shown a significant increase in bone mass in subjects with OA compared to sex-matched and age-matched controls8.
By contributing to the control of osteoclast differentiation, maturation, and activity, and owing to their fundamental role in the formation of new bone tissue, osteoblasts are crucial for the modulation of bone remodeling processes, whose alterations are determinant in the loss of bone mass and in the architectural changes observed in OP. Also, osteoblasts play a key role in bone changes typical of OA. It has been shown in vitro that osteoblasts derived from subchondral bone of OA subjects present an altered phenotype, such as an increased production of osteocalcin and alkaline phosphatase (ALP), an altered response to vitamin D stimulation9, and an increased expression of growth factors. These growth factors include insulin-like growth factor-1 and other cytokines10 that determine a change in bone remodeling and can influence and modulate the cartilage degradation processes11. The osteoprotegerin/receptor activator of nuclear factor-κB ligand (OPG/RANKL/RANK) cytokine system is essential for controlling osteoclast biology and bone remodeling processes, in both physiological and pathological conditions, including OA and OP. By binding to RANK on the surface of osteoclast precursors, RANKL leads to the activation of nuclear factor-κB and to determining the transcription of osteoclastogenetic genes, with subsequent induction of preosteoclast differentiation, increased osteoclast activity, and prolonged osteoclast lifespan12. OPG is the soluble receptor of RANK, which acts as a competitive inhibitor of RANKL13, reducing osteoclastogenesis and bone resorption. The RANKL/OPG ratio is a major determinant of bone mass14 and better reflects the bone remodeling status.
Another recently recognized regulator of bone formation and bone remodeling is the Wnt/β-catenin system, which represents one of the major signaling cascades in bone biology; any alteration in this signaling cascade causes imbalance in bone turnover and subsequent alterations of bone mass15. The activation of the Wnt/β-catenin pathway by the binding of the appropriate Wnt ligand with a coreceptor complex involving low-density lipoprotein receptor-related protein-5 (LRP-5) or LRP-6 and one of the frizzled protein family member leads to an accumulation of nonphosphorylated β-catenin. This in turn leads to activation of osteoblast gene transcription that promotes osteoblast commitment, proliferation, and differentiation, enhances osteoblast and osteocyte survival, and acts indirectly on bone-resorbing osteoclast activity. In vitro and in vivo studies suggested that OPG expression gene is regulated by the Wnt/β-catenin signaling pathway16.
It has been shown that the canonical Wnt-frizzled-β-catenin pathway is involved in the development and maintenance of joint bone and cartilage and could play a role in the pathogenesis of OA.
Better comprehension of the mechanisms underlying the complex pathogenic processes of OA and OP are essential for the development of new pharmacological approaches to bone diseases. Particularly, regulation of Wnt and Wnt antagonist actions, which represent a well-recognized new therapeutic target in OP treatment, has recently emerged as a potential strategy for controlling tissue remodeling and regeneration in joint degenerative diseases such as OA.
Our aim was to evaluate the different expression of Wnt signaling components, depending on the cellular context in human OA and OP osteoblasts both in basal conditions and after vitamin D3, and to correlate their expression to cell metabolic activity, proliferation, and OPG/RANKL ratio in culture medium.
MATERIALS AND METHODS
Normal, osteoporotic, and osteoarthritic human osteoblasts
Normal bone specimens were obtained from 11 healthy subjects (7 men, 4 women) aged 55.1 ± 9.6 years (mean ± SD, range 39–67) who were undergoing surgery for reduction of traumatic proximal femur fractures. On histological analysis, the bone tissue fragments showed good trabecular architecture and normal trabecular thickness, consistent with a normal bone structure. None of the recruited healthy subjects was affected by metabolic bone diseases and none received medication, including corticosteroids (which could interfere with bone metabolism), for 6 months prior to bone biopsy.
OA subchondral bone samples were obtained from 14 patients (8 men and 6 women) aged 62.3 ± 11.2 years (range 58–81) who were undergoing total hip joint replacement and who were classified as having hip OA based on clinical history, physical examination, laboratory tests, and radiographs. The bone specimens were selected depending on the integrity of overlying articular cartilage, identifying the areas in which macroscopic damage was visible. In the selected areas the cartilage surface appeared rough and eroded, yellowish, softened, with fibrillation phenomena or even completely absent. The macroscopic characteristics of the cartilage surface from the damaged areas were associated with a high degree of thickening and sclerosis of the corresponding subchondral bone, as confirmed by histological examination, that showed clear evidence of injury, consistent with fibrillation, erosion, ulcerations, cracking, and disruption of the cartilage surface. In the underlying bone, trabecular thickening, subarticular cysts (especially where cartilage was absent), and some focal areas of pressure necrosis were observed.
None of the recruited OA subjects were affected by other metabolic bone diseases and none received medication, including corticosteroids, for 6 months prior to bone biopsy.
Osteoporotic (OP) bone samples were obtained from 12 patients (11 women and 1 man) aged 65.8 ± 8.3 years (range 60–75), undergoing total hip joint replacement for OP fractures of the femoral neck. Patients with OP neck fractures were selected on the basis of the age of fracture onset, and younger patients were included in the study to obtain an OP population matched for age to the other study groups. Among the OP female patients, 4 had early menopause (before 45 years old, mean age 41). All OP subjects had 2 or more risk factors for fracture. In all OP patients, the cartilage surface corresponding to the bone fragments appeared normal, with no sign of degeneration. Histological examination of these bone fragments showed a reduction of trabecular number, thinning, and erosions, and loss of network between trabeculae, with an increase in bone marrow space, confirming the OP expression of the bone disease.
Appropriate informed consent was obtained from each patient and the study was approved by an institutional ethics committee.
Primary subchondral bone osteoblast cell cultures
We obtained for each patient (OA and OP) the entire replaced joint (specifically, the femoral head). From the surgical pieces we obtained very small fragments of cancellous bone using a needle bone biopsy. For healthy subjects (traumatic fracture) we used a small fragment obtained by the surgeon. Each fragment was washed using sterile phosphate buffered saline (PBS) to eliminate bone marrow cells, then digested with 0.5 mg/ml type II collagenase (PAA Laboratories) with Dulbecco modified Eagle’s medium (DMEM) without serum and supplemented with antibiotics (penicillin 100 IU/ml and streptomycin 100 mg/ml) for 1 h at 37°C to remove all fibroblasts and residual blood cells. Bone fragments were subsequently washed in DMEM supplemented with antibiotics and containing 20% fetal calf serum (FCS) to remove collagenase and then transferred in sterile flasks in the same medium at 37°C in a water-saturated atmosphere containing 5% CO2. After 24 h, the medium containing many nonadherent cells was removed, the bone fragments were extensively washed with PBS, and fresh medium was added. When cells were observed in the flasks, the culture medium was replaced every 3 days with fresh medium containing 10% FCS. Osteoblasts began to grow from the bone specimens after about 1 week and proliferated on the flask surface, reaching confluence within 3–4 weeks, as described. In all previous in vitro studies on osteoblasts, pathological phenotypes appear to be maintained after this time17. On reaching confluence, osteoblasts were isolated using trypsin 1% for 5 min, with subsequent temperature inactivation at 37°C, then seeded in 24-well plates in DMEM containing 10% FCS and antibiotics at 60 × 103 cells for every plate until semiconfluence was reached, and then cultures were treated with 1,25-dihydroxy vitamin D3 [1,25(OH)2D3; Roche] 10−8 M for 48 h or left under basal conditions. All experiments were performed at first passage after osteoblast isolation to avoid the dedifferentiation of osteoblasts into fibrobast-like cells.
To characterize osteoblast phenotype and cell metabolic activity, we evaluated osteocalcin and ALP levels18 in cell cultures; further, levels of DKK-1, DKK-2, nonphosphorylated β-catenin, OPG, and RANKL were determined on culture medium. For evaluation of cell proliferation, the isolated osteoblasts were seeded in 96-well plates in the same experimental conditions.
All experiments were performed in triplicate for each patient sample.
Intracellular total protein content
The intracellular total protein content in cell lysate, directly related to a total number of metabolically active cells, was determined by the Bradford method, using a Bio-Rad protein assay kit (Bio-Rad Laboratories), based on the color change of Coomassie Brilliant Blue G-250 dye in response to various concentrations of proteins. The dye reagent was added to the cell lysate, obtained by dissolving in 0.1% sodium dodecyl sulfate, and to 5 dilutions of bovine serum albumin (BSA), used as standard; the absorbance was measured at 595 nm with a spectrophotometer. Once the standard curve was drawn, by plotting the absorbance at 595 nm versus the BSA concentration, the unknown concentration of samples was obtained by interpolation and expressed as mg/ml.
DKK-1, DKK-2, OPG, and RANKL levels
Cell medium of cultures both in basal conditions and after 48 h of 1,25(OH)2D3 exposure was collected and then preserved at −80°C until used for quantification of DKK-1, DKK-2, OPG, and RANKL levels.
DKK-1 and DKK-2 were measured by ELISA kits (R&D Systems and USCN Life Science Inc.). The unknown concentrations of samples, obtained from the standard curves, were expressed as pg/ml for DKK-1 and ng/ml for DKK-2 and were normalized to the mg intracellular protein.
Levels of OPG and RANKL were measured with ELISA kits according to the instructions supplied by the manufacturer, BioVendor Laboratory Medicine. The results, expressed as pmol/l, were directly determined from the calibration curves and normalized to the mg intracellular protein.
All measurements were performed in triplicate for each sample, and the mean values were calculated.
β-catenin qualitative determination
The active, nonphosphorylated form of β-catenin in lysate of treated and untreated cells was detected by qualitative ELISA (Novus Biologicals), according to the manufacturer’s recommendations. Samples were incubated in antibody-coated microwells for 2 h and after washing to remove unbound antibodies, the biotin-conjugated detection antibody was added and incubated a further 2 h. After another washing, streptavidin horseradish peroxidase conjugate was added for 30 min. The last washing step was carried out and the remaining conjugate was allowed to react with the substrate solution in the dark; absorbance of the resulting yellow products was read spectrophotometrically at 450 nm. The positive control, supplied by the manufacturer, was used to assess the performance of the assay. Results were expressed as optical density (OD) ± SD.
Osteocalcin and ALP production
Osteocalcin and ALP production was assessed in osteoblast cultures both in basal condition and after 48 h incubation with 1,25(OH)2D3. Osteocalcin release into the culture medium was measured using an intact human osteocalcin ELISA. Results were normalized per mg/cell proteins and osteocalcin production was expressed as ng osteocalcin/mg cell protein. Cell lysate was tested for ALP production, using a commercial ELISA kit (Cusabio). The concentration of ALP in the samples, determined by comparing the OD, measured spectrophotometrically at 450 nm, to the standard curve, was normalized per mg/cell proteins, and ALP production was expressed as ng ALP/mg cell protein.
All experiments were performed at first passage and in triplicate.
Assessment of cell proliferation
Proliferation rates of both treated and untreated cells were determined using the MTT proliferation assay, based on the conversion of yellow tetrazolium salt MTT to purple formazan crystal by actively proliferating cells; results reflect the number of viable cells. Osteoblasts were plated at a density of 1 × 104 cells/cm2 with and without 1,25(OH)2D3 (10−8 M).
After 48 h incubation, the medium was aspirated. Three hundred microliters of MTT [3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide; stock solution 50 mg/ml; Sigma] was added to each well and cultures were continued for 2 h at 37°C. This permitted cellular conversion of MTT to insoluble formazan. The cells were then lysed and the formazan dissolved with acidic propanol at room temperature for 24 h. Two hundred microliters of supernatant was transferred to microplate wells and colorimetric changes were quantified in a microplate reader at an OD of 540 nm (Multiskan EX, Thermo Electron Corp.). The percentage of viable cells was expressed as percentage ratio between the OD of samples and OD of control (untreated normal cells)19.
Statistical analysis
Results were expressed as means ± SD. The differences in osteocalcin production, OPG/RANKL, and DKK-1 expression between the different cell populations were assessed with the nonparametric Kruskal-Wallis test, using the Dunn test for multiple comparisons. Differences in proliferating activity were assessed using the Bonferroni test. A p value < 0.05 was considered significant.
RESULTS
Results are summarized in Table 1. We found opposite patterns of DKK-1 and DKK-2 production in OA and OP osteoblasts. In basal conditions, DKK-1 production was significantly greater in OP cells compared to the control cells (p < 0.001); conversely, the OA osteoblasts showed a significantly lower production of DKK-1 compared to the control osteoblasts (p < 0.05). The addition of 1,25(OH)2D3 induced a significant decrease of DKK-1 expression in all cell populations (Figure 1A). Conversely, DKK-2 levels were significantly increased in the culture medium of OA osteoblasts compared to the control cells (p < 0.001), whereas the reduction was observed in osteoporotic cells (p < 0.05). Treatment with 1,25(OH)2D3 increased DKK-2 production in each cell population (Figure 1B).

Dickkopf protein factor 1 (DKK-1) and DKK-2 expression in normal (N), osteoporotic (OP), and osteoarthritic (OA) osteoblasts, in basal conditions and after stimulation with 1,25-dihydroxy vitamin D3 [1,25(OH)2D3]. A. DKK-1 production is significantly greater in OP cells compared to normal cells (*p < 0.001); conversely, the OA osteoblasts show a significantly lower production of DKK-1 compared to normal osteoblasts (**p < 0.05). The addition of 1,25(OH)2D3 induced a significant decrease in DKK-1 expression in all cell populations (§p < 0.05). Results are normalized per mg/cell proteins. B. DKK-2 production is significantly greater in OA cells compared to normal cells (**p < 0.001); conversely, significantly lower production of DKK-2 was found in OP cells compared to normal osteoblasts (*p < 0.05). The addition of 1,25(OH)2D3 induced a significant increase in DKK-2 expression in all cell populations (§p < 0.05). Results are normalized per mg/cell proteins.
Study results.
Dephosphorylated β-catenin qualitative determination
The dephosphorylated β-catenin levels were significantly higher in OA osteoblasts compared to control cells (Figure 2; p < 0.01); conversely, OP osteoblasts showed lower levels of dephosphorylated β-catenin compared to control cells (p < 0.05). No significant differences were found after 1,25(OH) 2D3 supplementation.

Quantitative determination of nonphosphorylated form of β-catenin in normal (N), osteoporotic (OP), and osteoarthritic (OA) osteoblasts, in basal conditions and after stimulation with 1,25-dihydroxy vitamin D3 [1,25(OH)2D3]. Optical density (OD) of OP cells is significantly lower compared to normal cells (*p < 0.01); conversely, OA osteoblasts show significantly higher absorbance compared to normal osteoblasts (**p < 0.05). No differences were found after exposure to 1,25(OH)2D3.
RANKL/OPG ratio
The RANKL/OPG ratio was different in the osteoblast populations and showed an opposite feature in OP and OA cells. Indeed, the basal RANKL/OPG ratio in the absence of 1,25(OH)2D3 stimulation was significantly higher in osteoporotic cells compared to the control cells (p < 0.001; Figure 3A). Conversely, the basal RANKL/OPG ratio was significantly lower in OA osteoblasts compared both to the control osteoblasts (p < 0.05) and to the OP osteoblasts (p < 0.001). Treatment with 1,25(OH)2D3 increased the RANKL/OPG ratio in the 3 cell populations (Figure 3A).
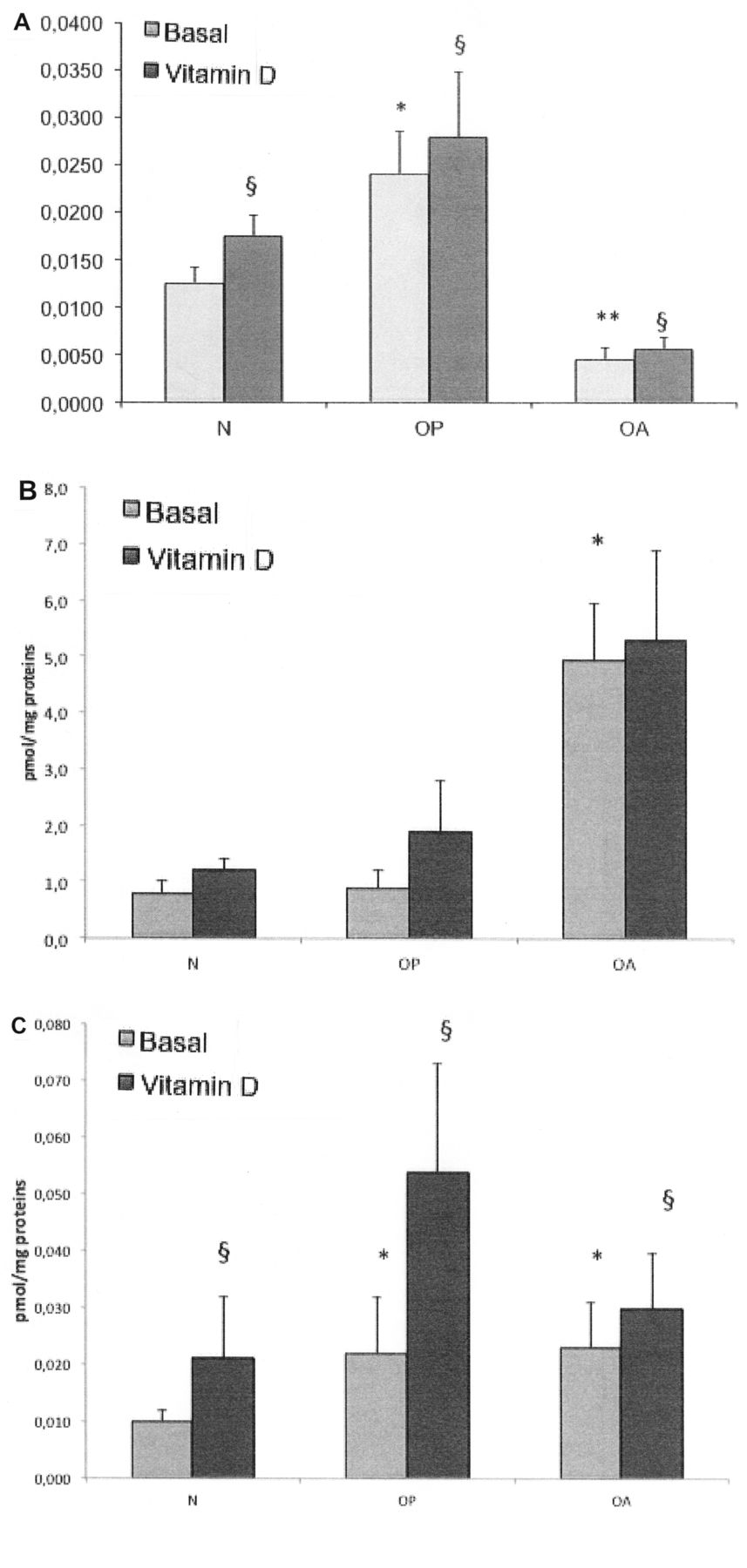
RANKL/OPG levels in normal (N), osteoporotic (OP), and osteoarthritic (OA) osteoblasts, in basal conditions and after stimulation with 1,25(OH)2D3. A. RANKL/OPG ratio. In basal conditions [absence of 1,25(OH)2D3], the RANKL/OPG ratio is significantly lower in OA osteoblasts compared to both normal (**p < 0.05) and OP osteoblasts (*p < 0.001). Treatment with 1,25(OH)2D3 increases the RANKL/OPG ratio in the 3 cell groups (§p < 0.05). B. OPG expression. OPG is significantly higher in OA osteoblasts compared to normal and OP osteoblasts (*p < 0.0001), whereas there are no differences in OPG production between normal and osteoporotic cells. C. RANKL expression. In basal conditions, RANKL production is significantly higher in OP and OA osteoblasts compared to the normal ones (*p < 0.01) without any difference in the 2 groups of pathological cells. Stimulation with 1,25(OH)2D3 significantly enhances RANKL production (§p < 0.0001).
When analyzing the isolated expression of OPG and RANKL in control and pathological osteoblast cultures, we found that OPG was significantly higher in OA osteoblasts compared to both control and OP osteoblasts (p < 0.0001), whereas there were no differences in OPG production between control and OP cells (Figure 3B). Conversely, we found that RANKL production was significantly higher both in OP and in OA osteoblasts compared to the control cells, without any difference in the 2 groups of the pathological cells (Figure 3C). Of note, although in OA osteoblasts both RANKL and OPG levels were higher compared to the control cells, OPG levels were much more elevated than those of RANKL (about 3-fold greater), thus explaining the lower ratio of RANKL/OPG in these cells. Stimulation with 1,25(OH)2D3 enhanced both OPG and RANKL production, but in a greater extent for RANKL, determining the increase of the RANKL/OPG ratio in all cell populations.
Cell proliferation
In basal conditions, OP osteoblasts showed a significantly lower proliferating activity compared to control osteoblasts (–66.37%, or corresponding to 33.63% of proliferating activity of normal osteoblastic cells). Conversely, OA osteoblast proliferation was slightly greater compared to proliferation of the control cells (+3.78%) but did not reach a significant difference. Cell proliferation was significantly enhanced by 1,25(OH)2D3 stimulation in OP cell cultures (p < 0.05) and in OA cell cultures (p < 0.01; Figure 4).

Proliferating activity of osteoporotic (OP) and osteoarthritic (OA) osteoblasts, in basal conditions and after stimulation with 1,25(OH)2D3. In basal conditions, cell proliferation of OP osteoblasts is significantly lower compared to normal osteoblasts (*p < 0.01). Conversely, OA osteoblast proliferation is slightly greater compared to proliferation of normal cells but does not reach a significant difference. Cell proliferation is significantly enhanced by 1,25(OH)2D3 stimulation in OP and in OA cell cultures (§p < 0.01). Results are expressed as percentage ratio compared to proliferating activity of control cells (untreated normal cells).
Osteocalcin production and ALP activity
Stimulation with 1,25(OH)2D3 increased osteocalcin synthesis in all cell populations (OA, OP, and control osteoblasts). According to published data, in OA osteoblasts, osteocalcin synthesis was significantly higher compared to the control osteoblasts, both before and after 1,25(OH)2D3 culture supplementation. Conversely, in OP osteoblasts we found a significantly lower osteocalcin synthesis compared to the control cells (Figure 5). ALP was synthesized by all cell populations, and was enhanced by 1,25(OH)2D3 stimulation, confirming the osteoblast phenotype of studied cells. As shown in Figure 6, OA osteoblasts produced significantly higher levels of ALP compared to the normal cells, whereas in osteoporotic cells ALP levels were significantly lower.

Osteocalcin production of normal (N), osteoporotic (OP), and osteoarthritic (OA) osteoblasts, in basal conditions and after stimulation with 1,25(OH)2D3. In OP osteoblast cultures, osteocalcin synthesis is significantly lower compared to normal cells (*p < 0.05), both before and after 1,25(OH)2D3 culture supplementation; osteocalcin synthesis is significantly higher in OA osteoblasts compared to the normal osteoblasts, both in basal conditions and after 1,25(OH)2D3 stimulation (**p < 0.001). In all cell groups, osteocalcin synthesis was enhanced by 1,25(OH)2D3. Osteocalcin levels are normalized per mg intracellular proteins.
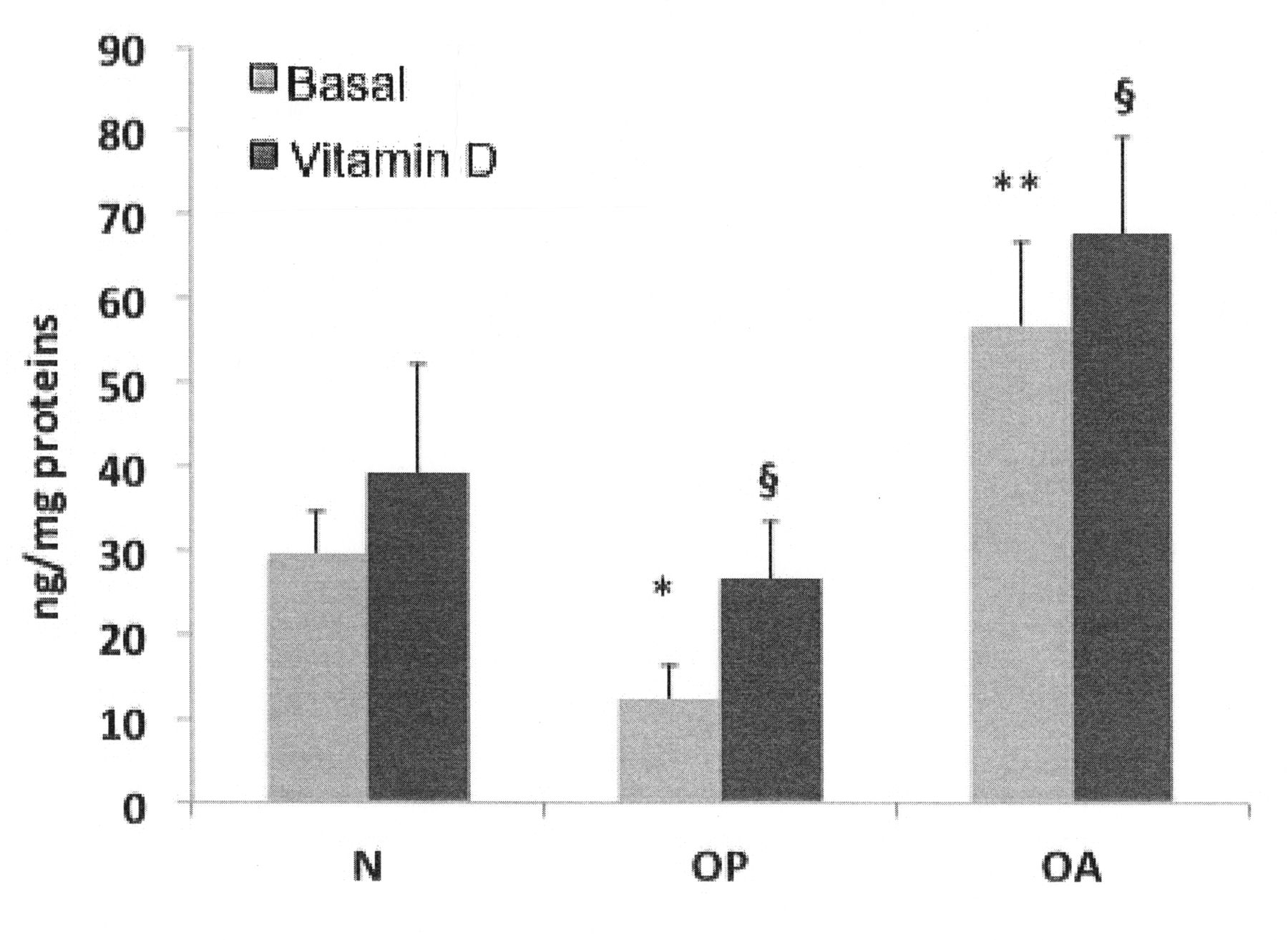
Alkaline phosphatase (ALP) production of normal (N), osteoporotic (OP), and osteoarthritic (OA) osteoblasts, in basal conditions and after stimulation with 1,25(OH)2D3. OA osteoblasts produce significantly higher levels of ALP compared to the normal osteoblasts, both in basal conditions and after 1,25(OH)2D3 stimulation (**p < 0.05). In OP osteoblast cultures, ALP production is significantly lower compared to normal cells (*p < 0.01), both before and after 1,25(OH)2D3 culture supplementation. Stimulation with 1,25(OH)2D3 increases ALP synthesis in both normal and pathological osteoblasts. ALP levels are normalized per mg intracellular proteins.
DISCUSSION
OP and OA are 2 chronic diseases in which opposite alterations of bone structural characteristics and bone remodeling processes have been described. Osteoblasts play a key role in bone remodeling processes because they are directly responsible for bone formation and are essential in controlling bone resorption by regulating the recruitment, proliferation, and differentiation of osteoclasts. These processes are strictly modulated by various systemic and local growth factors, such as the components of the RANK/RANKL/OPG system and Wnt pathway. The canonical Wnt pathway is regulated by various elements exerting multiple and often redundant effects, including ligands, receptors, coreceptors, and several inhibitors, such as members of the DKK family. This family consists of 4 proteins with different patterns of expression and different biological activity20. Whereas DKK-1 and DKK-4 have an inhibitory effect on Wnt signaling, a dual role of DKK-2 as an inhibitor or an activator of Wnt signaling has been described; the DKK-3 effects remain unknown20. It has been shown that various regulatory components of the Wnt system, such as LRP-5, LRP-6, DKK-1, and DKK-2, are expressed in primary cultured human osteoblasts in vitro21. The key role of Wnt signaling in the pathogenesis of OP has been clearly established, but recently several studies showed its involvement in OA pathogenesis, too. Allelic variants of an antagonist of Wnt signaling, the secreted frizzled-related protein, can predispose to hip OA by altering the development or stability of cartilage or bone in weight-bearing loss22; further, the mechanical injury to cartilage explants results in downregulation of the secreted frizzled-related protein and upregulation of Wnt target genes such as axin 223, type II collagen, and aggrecan expression24. The expression of the Wnt-induced signaling protein 1 was increased in an experimental model of OA and in human OA cartilage25. It has been suggested that high circulating levels of DKK-1 may be associated with a slower progression of hip OA26 and that plasma and synovial fluid levels of DKK-1 are inversely related to radiographic grading of knee OA27.
We have found that primary human osteoblasts from patients with hip OA and osteoporotic hip fracture behave abnormally compared to the normal cells involved in the regulation of the Wnt pathway. In particular, we found that OA osteoblasts are characterized by an increased proliferative and metabolic activity in terms of osteocalcin production, and that this might generate the typical alterations observed in subchondral bone in this disease, such as thickness, sclerosis, and increased stiffness.
Conversely, we observed that OP osteoblasts are metabolically less active cells and present a reduced proliferative cell activity, thus contributing to the reduction of bone mass. Interestingly, we found that the alterations in pathological osteoblasts involve both the modulation of the Wnt system and the OPG/RANKL system. Indeed, β-catenin levels were upregulated in OA osteoblasts and downregulated in OP osteoblasts when compared to those of control cells. Further, the DKK-1 expression was lower in osteoblasts from patients with OA compared to osteoblasts from healthy subjects, whereas it was significantly higher in osteoblasts from patients with OP hip fractures. Conversely, the DKK-2 expression was significantly higher in OA osteoblasts and lower in OP cells. These data seem to confirm the hypothesis that the Wnt system is involved in the pathogenesis of both diseases and particularly that it is downregulated in OP and upregulated in OA. Osteoblasts can participate in the regulation of the Wnt system in these pathological conditions, through the synthesis of the Wnt antagonist DKK-1 and DKK-2, whose pattern of expression appears to be opposite when comparing OA and OP cells. These data agree with the findings of a study showing that decreased levels of DKK-1 are essential for local new bone formation and the development of osteophytes, which are the hallmark of OA28. Moreover, the increase of DKK-2 observed in the same cells further supports the hypothesis that this factor could contribute to the abnormal phenotype of OA osteoblasts. The different regulation of the Wnt system could reflect the different expression of osteoblast-specific proteins such as osteocalcin, whose synthesis is enhanced in OA osteoblasts and is reduced in the OP ones, according to published data9.
We found that RANKL/OPG ratio in cell cultures is reduced in OA osteoblasts, confirming that in subchondral OA bone an imbalance of bone turnover favoring a new bone formation occurs, and this is supported by a greater production of OPG rather than RANKL. Conversely, the RANKL/OPG ratio is increased in osteoporotic osteoblasts as a result of the increase in RANKL, whereas in these cells the OPG expression is similar to that of normal cells, consistent with a condition of increased bone remodeling with an enhanced bone resorbing activity.
Indeed, when absolute levels of OPG and RANKL were evaluated we found that OPG was significantly higher in OA osteoblasts compared to the normal and OP cells, whereas RANKL was significantly higher in OA and OP osteoblasts compared to the normal ones, without any differences between the pathological cells. Other authors demonstrated an increased production of OPG in a subgroup of OA osteoblasts that concurred with the histologic aspect of subchondral bone that appears to favor bone formation29.
The production of OPG and RANKL is enhanced by 1,25(OH)2D3, but the increase in the latter is much higher, determining the increase of the RANKL/OPG ratio in all cell population groups, which is indicative of a possible increase of bone remodeling. These findings agree with previous reports showing that 1,25(OH)2D3 increases messenger RNA (mRNA) expression of RANKL and RANKL/OPG ratio in various osteoblast cell lines and human primary cells29,30,31,32, and that 1,25(OH)2D3 is able to promote osteoclastogenesis by acting on the differentiation of osteoclast precursors through osteoblasts33. According to the presented data, 2 different groups found that on biopsies of the iliac crest from patients with OP, the mRNA levels of OPG were lower, and RANKL and RANKL/OPG ratio levels were higher compared to iliac crest bone biopsies from patients with OA34,35. Further, it has been shown that the OPG/RANKL ratio is altered in OA osteoblasts and differs according to the metabolic state of these cells29. Another study demonstrated that OPG secretion in osteoblasts from patients with hip fractures is higher compared to OPG secretion in osteoblasts from patients with OA36. In other studies, the expression of OPG and RANKL genes has been evaluated in both OA and OP, prevalently in cell pools but not in isolated osteoblasts, and under different experimental conditions, with inconsistent results32,37. These conflicting findings may be related to the use of different experimental models, or to the different cell origin and culture conditions. The normal and pathological cells we analyzed were derived from the same anatomical region, so that the comparison between the study groups would not be biased by the anatomical site of cell origin.
The differences we found between OP and OA osteoblasts in proliferating activity, osteocalcin production, DKK-1, DKK-2, and β-catenin expression and OPG/RANKL ratio could reflect differences in cell metabolism related to the specific pathology. OP osteoblasts present a reduced synthesis of the bone-specific protein osteocalcin and are responsible for a reduced bone-forming activity, whereas stimulated bone resorption through an increased expression of RANKL. Conversely, OA osteoblasts show a higher synthesis of osteocalcin, which is the expression of higher metabolic activity compared to OP osteoblasts. OA osteoblasts also present a higher proliferation rate and affect bone remodeling, causing an imbalance toward a bone-forming activity through a proportionally greater expression of OPG rather than RANKL.
The main limitations of our study were the restricted number of healthy and pathological subjects, and the in vitro experimental conditions, which might not truly represent the complete in vivo conditions, in which many other factors (such as growth factors, cytokines, and interactions with other cell types) might be involved.
Nevertheless, these preliminary data support the hypothesis that both OA and OP osteoblasts might contribute to the regulation of the Wnt pathway through an autocrine/paracrine mechanism that involves the synthesis of DKK-1. In OA, the Wnt pathway upregulation may have a stimulatory effect on bone formation and could be related to the bone sclerosis and osteophyte formation observed in the affected joints.
Our results support the hypothesis for the existence of a pathogenic link between DKK family members and the pathogenic processes underlying OA. Additional investigation will be required to clarify the roles of DKK-1, DKK-2, and the other components of the Wnt system in OA pathogenesis, to determine both their real potential roles as biochemical markers and to develop effective pharmacological agents to inhibit the disease progression.
- Accepted for publication December 27, 2012.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}