

Abstract
Objective. To investigate B cell survival cytokine messenger RNA (mRNA) levels as biomarkers of idiopathic inflammatory myopathies (IIM).
Methods. We measured and compared mRNA levels of B cell survival cytokines by quantitative real-time polymerase chain reaction in 98 patients with IIM, 38 patients with systemic lupus erythematosus, and 21 healthy controls. The cytokines were B cell-activating factor belonging to the tumor necrosis factor family (BAFF); ΔBAFF; and a proliferation-inducing ligand (APRIL); and their receptors BAFF-R, transmembrane activator and calcium modulator and cyclophilin ligand interactor, and B cell maturation antigen (BCMA). We also identified autoantibodies, including anti-Sm, anti-RNP, anti-SSA/Ro, anti-SSB/La, anti-topoisomerase 1, anti-hystidyl-tRNA synthetase, anti-ribosomal P, and anti-chromatin. Clinical disease activity was assessed by the International Myositis Assessment and Clinical Studies core set tool. We examined correlation of mRNA with disease activity, medication use, and autoantibodies.
Results. We found a positive correlation of BAFF and ΔBAFF expression with 3 disease activity measures, with ΔBAFF having a stronger correlation. Similarly, anti-SSA/Ro-52 and/or anti-SSA/Ro-60 had a strong positive correlation with mRNA levels of BAFF and ΔBAFF, and with relative ratios of BAFF/APRIL and BCMA/BAFF-R.
Conclusion. These findings highlight the potential importance of BAFF, ΔBAFF, and BAFF-R in the pathogenesis of IIM, and suggest an important role in the assessment of disease activity.
- AUTOANTIBODIES
- B CELL ACTIVATING FACTOR
- B CELL ACTIVATION FACTOR RECEPTOR
- B CELL MATURATION ANTIGEN
- IDIOPATHIC INFLAMMATORY MYOPATHIES
- PROLIFERATION-INDUCING LIGAND
The idiopathic inflammatory myopathies (IIM) are systemic connective tissue diseases characterized pathologically by myonecrosis and chronic, acquired skeletal muscle inflammation resulting in symmetrical, proximal muscle weakness, decreased muscle endurance, and elevated muscle enzymes, including creatine kinase. They can be subclassified as polymyositis (PM), dermatomyositis (DM), and inclusion body myositis, according to differences in their clinical and immunohistopathologic features1. Although the initiating factors of inflammation are unknown, immune-mediated processes have a profound effect on the pathophysiology of IIM. Briefly, PM is a cell-mediated autoimmune disorder in which cytotoxic CD8+ lymphocytes and macrophages invade and destroy myofibers expressing antigens presented by MHC class 12. In DM, the cellular infiltrate consists of CD4+ T cells, B cells, and plasmacytoid dendritic cells2,3,4. The inflammatory component of inclusion body myositis is similar to that of PM1.
Studies have shown that specific B cell responses are ongoing in the skeletal muscle of patients with IIM. Cytokines and chemokines may influence antigen presentation by muscle-derived dendritic cells and primary muscle cells and may affect the endoplasmic reticulum stress response3,4,5,6. Moreover, reports of clinical studies7,8 on B cell-targeted therapy using anti-CD20 antibodies (i.e., rituximab) document an impressive symptomatic improvement in patients with refractory myositis, which has moved B cells onto the center stage of clinical and laboratory investigation of IIM. The effect of B cell depletion with rituximab in IIM implies the presence of still unknown roles for B cells in the pathogenesis of IIM. One of the most crucial factors for the maintenance of B cells is the BAFF (B cell-activating factor belonging to the tumor necrosis factor family)/APRIL (a proliferating-inducing ligand) system. BAFF and APRIL belong to the tumor necrosis factor (TNF) superfamily ligands9,10. Despite their similar structure and shared receptors, both ligands have distinct functions11. Whereas BAFF is essential to the survival of transitional and mature B cells, APRIL affects the activity of B-1 cells as well as humoral responses and immunoglobulin class switching; it also determines the size of the peripheral B cell pool12,13. Both BAFF and APRIL are homotrimeric type 2 transmembrane proteins that also exist as soluble proteins derived from the cleavage of transmembrane forms11,14. Whether the biologic functions controlled by membrane-bound BAFF differ from those triggered by soluble BAFF remains unclear. The noncleavable form ΔBAFF is localized to the cell surface and lacks the normal biologic activities of BAFF15; ΔBAFF-transgenic mice have suppressed BAFF activity, which indicates that ΔBAFF downregulates BAFF activity in vivo16. Unlike BAFF, APRIL is cleaved intracellularly rather than at the cell surface, which impedes detection of its integral form at the plasma membrane17. BAFF and APRIL are mainly expressed by monocytes, dendritic cells, and T cells18, whereas their receptors [BAFF-R, transmembrane activator and calcium modulator and cyclophilin ligand interactor (TACI), and B-cell maturation antigen (BCMA)] are expressed mainly by B cells11,19. Evidence indicates that dysregulation of the BAFF/APRIL system is involved in the pathogenesis of B cell-related autoimmune diseases, with increased serum levels of BAFF reported in some conditions20,21,22,23,24. Moreover, blockade of BAFF activity induces clinical improvement in patients with autoimmune diseases, such as systemic lupus erythematosus (SLE) or rheumatoid arthritis25,26, and may represent an effective strategy to treat B cell-related autoimmune diseases. Although increased serum levels of BAFF have also been documented in patients with IIM24,27, the degree of elevation varies widely and may not be a direct reflection of disease activity. However, serum BAFF protein levels do not measure membrane-bound BAFF, so messenger RNA (mRNA) levels of BAFF might be a more sensitive and accurate measure of peripheral activity. Thus, we examined mRNA-expression levels of these TNF B cell survival cytokines and their receptors, and their correlations with disease activity, medication use, and autoantibodies.
MATERIALS AND METHODS
Patients and clinical assessment
Our study was approved by the Mayo Clinic Institutional Review Board, and informed consent was obtained from each participant. A total of 98 adult and pediatric patients were studied. They had varying degrees of disease activity and disease duration, ranging from new onset (n = 23) to several years after diagnosis. All patients had a definitive diagnosis of IIM and were being followed in the Division of Rheumatology at the Mayo Clinic, Rochester, Minnesota, USA. Associated connective tissue disease occurred in 3 patients (2 had Sjögren syndrome and 1 had antisynthetase syndrome).
Demographic and clinical data were abstracted from the medical records of all participants. For some analyses, healthy individuals with no history of inflammatory or malignant disease served as controls. To determine whether the BAFF profile in patients with IIM differs from that in patients with SLE (a prototype autoimmune disease), we also included a disease control group for some analyses, comprising 38 patients who fulfilled the American College of Rheumatology criteria for SLE28 (35 females and 3 males; mean age 45.5 years, range 16–69 years). The patients with SLE had varying degrees of disease activity (evaluated using the SLE activity measure index) and disease duration (ranging from untreated onset to 36 years after diagnosis).
In patients with IIM (63 females and 35 males) with a mean age of 46.7 years (range 2–79 years), disease activity was assessed by scores on the International Myositis Assessment and Clinical Studies (IMACS) core set tool, including the physician’s score of extramuscular, muscular, and overall disease activity; muscle enzyme evaluation; and manual muscle testing29. Muscle strength was evaluated using the manual muscle testing score, as described30,31. Extramuscular activity was determined using the Myositis Disease Activity Assessment Visual Analog Scale and the extra-skeletal visual analog scale (VAS)31,32. A Luminex bead-based immunoassay (Luminex Corp.) was used to detect anti-Sm, anti-RNP, anti-SSA/Ro (subtypes 52 kDa and 60 kDa), anti-SSB/La, antitopoisomerase 1 (anti-Scl-70), anti-hystidyl-tRNA synthetase (anti-Jo-1), antiribosomal P, and antichromatin antibodies in serum samples collected at the timepoint corresponding with BAFF analysis. Antinuclear antibodies and anti-dsDNA were measured using commercial enzyme-linked immunoassay kits. The antibody assays were performed in the Antibody Immunology Laboratory at the Mayo Clinic.
Total RNA isolation and quantitative real-time polymerase chain reaction (PCR) experiments
A 10-ml peripheral blood sample was collected in a tube containing sodium heparin. Peripheral blood mononuclear cells (PBMC) were isolated by density gradient centrifugation. Total RNA was extracted from mononuclear cells using the RNeasy Mini Kit (Qiagen Inc.), and RNA concentration was estimated using the ND-1000 Spectrophotometer (NanoDrop Products; Thermo Fisher Scientific Inc.). About 100 ng of total RNA was reverse-transcribed using an oligo primer (Mayo Core Facilities) and Moloney murine leukemia virus reverse transcriptase (Life Technologies Corp.). Absolute quantification standards were prepared by PCR amplification of complementary DNA using the cell line human acute myelocytic leukemia. Primers and probes were specifically designed to amplify mRNA of interest with sequence-specific primers designed to span introns (Life Technologies Corp.). Gene expression of full-length BAFF (FL BAFF), ΔBAFF, APRIL, BCMA, TACI, BAFF-R, and CD20 was measured by quantitative real-time PCR using primers uniquely designed to amplify mRNA of interest with SYBR Green PCR Master Mix (Life Technologies Corp). The sequences of pairs of primers and fluorogenic probes used for amplification are listed in Table 1. The mean of triplicate PCR determined the mRNA copy number. Specificity of PCR amplicons was validated by pyrosequencing. Data were represented as absolute mRNA copy number and was normalized to 2 referent samples run on all plates.
Primers and probe sequences used for quantitative real-time polymerase chain reaction.
Statistical analysis
Descriptive statistics [median, percentage (range)] were used to summarize the data. The log-transformed quantitative real-time PCR gene expression data from multiple plates were normalized by dividing each patient’s values by the values of a common referent patient measured on the same plate, and then multiplying each value by the overall mean of the common referent across all plates. The Spearman rank correlation coefficient was calculated to assess the association between measures of interest and IIM disease features. A ratio was computed to evaluate BAFF, APRIL, and BCMA mRNA expression compared to CD20 mRNA expression. Partial correlation methods were used to adjust for global disease activity. Group comparisons were performed using the rank-sum test.
RESULTS
The study included 98 persons with IIM (63 females and 35 males) with a mean age of 46.7 years (range 2–79 years). Sixty-five were diagnosed with DM (20 juvenile DM and 45 adult DM), 26 with PM defined by the Bohan and Peter criteria33, and 7 with inclusion body myositis34. Most (96%) were white; of the rest, 1% was black, 1% was Asian, and 2% were multiracial. Table 2 summarizes the characteristics of the 98 patients with IIM, the 38 patients with SLE, and the 21 controls. Altogether, 28% of the patients with IIM had autoantibodies, with the most common being anti-SSA/Ro (subtypes 52 kDa and 60 kDa), anti-SSB/La, and anti-Jo-1. Anti-SSA 52-kDa and anti-SSA 60-kDa antibodies were detected in 54% and 19% of patients with PM, and in 16% and 8% of patients with DM, respectively. Anti-Jo-1 was observed in all IIM subgroups (27% PM, 14% inclusion body myositis, and 5% DM). Patients were either untreated (n = 22), or treated with prednisone, either alone or in combination with disease-modifying antirheumatic drugs (DMARD) such as methotrexate, azathioprine, mycophenolate mofetil, and hydroxychloroquine. Fifty-six patients (57%) were taking corticosteroids at the time of the blood draw.
Demographic characteristics of 98 patients with idiopathic inflammatory myopathies (IIM) compared to 38 patients with systemic lupus erythematosus (SLE) and 21 healthy controls. Values are number (%) unless specified otherwise.
Altered expression of BAFF survival pathway-related genes in patients with IIM
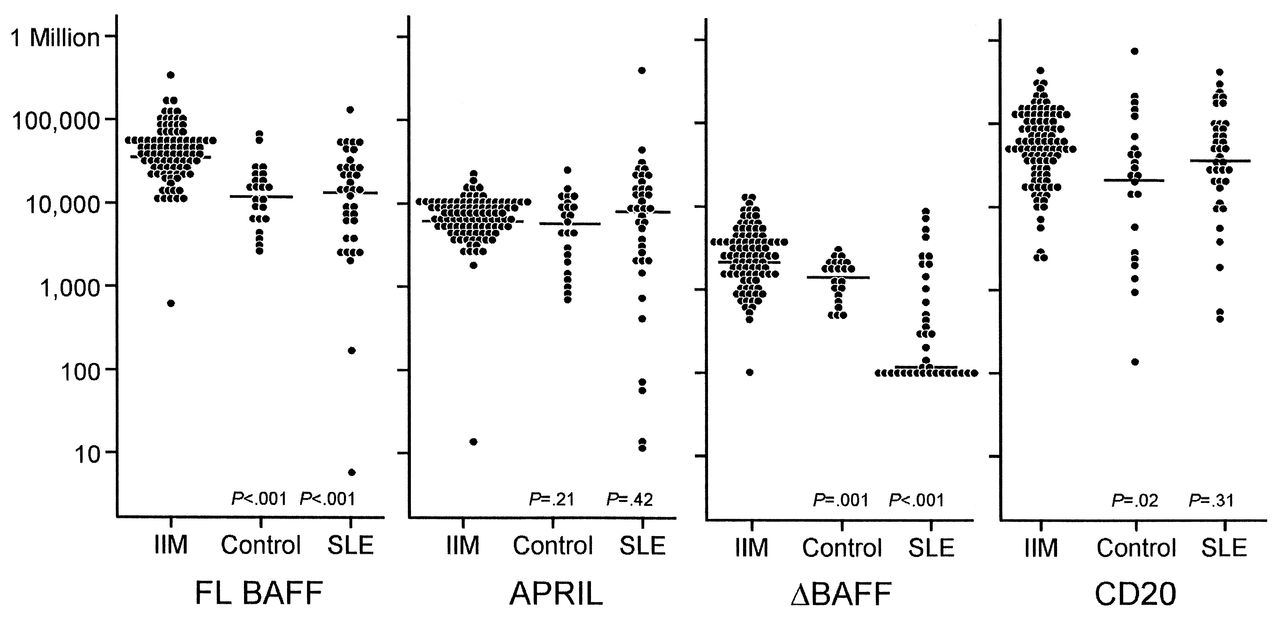
We investigated mRNA expression of FL BAFF, ΔBAFF, and APRIL and their receptors (BAFF-R, TACI, and BCMA) in PBMC from the 98 patients with IIM, and compared it with mRNA expression from 38 patients with SLE and from 21 healthy controls. The median level of BAFF mRNA in PBMC from patients with IIM was significantly increased compared to that in PBMC from patients with SLE and healthy controls (p < 0.001 for each; Figure 1). No significant differences were observed in BAFF expression in patients with SLE compared to that in healthy controls (p = 0.70). CD20 mRNA levels in patients with IIM were greater when compared to controls (p = 0.02). No significant differences were observed between the CD20 mRNA levels in patients with SLE and those in healthy controls (p = 0.19). No significant differences in BAFF mRNA levels were found between the subgroups of IIM subjects (p = 0.56). In addition to increased expression of FL BAFF in patients with IIM, the level of ΔBAFF mRNA expression was increased in subjects with IIM compared to patients with SLE (p < 0.001) and in healthy controls (p = 0.001). Notably, mRNA copy number levels of ΔBAFF were dramatically decreased in patients with SLE (median 134.5, range 100.0–7851.7) compared to those in healthy controls (median 1584, range 454.1–3090.4; p < 0.001; Figure 1). Although some IIM subjects exhibited high APRIL copy numbers, no significant differences were detected in average APRIL mRNA levels compared to patients with SLE (p = 0.42) or in healthy controls (p = 0.21). Patients with SLE had APRIL levels similar to those in healthy controls (p = 0.24).

Plots comparing absolute full-length B cell-activating factor belonging to the TNF family (FL BAFF), a proliferation-inducing ligand (APRIL), ΔBAFF, and CD20 messenger RNA (mRNA). FL BAFF, APRIL, and ΔBAFF mRNA expression were measured in PBMC from patients with idiopathic inflammatory myopathies (IIM), from patients with systemic lupus erythematosus (SLE), and from healthy controls using quantitative real-time PCR. X axis represents FL BAFF, APRIL, ΔBAFF, and CD20 mRNA after log-transformation. Horizontal bars indicate medians. Y axis represents absolute copy number gene expression. Median FL BAFF and median ΔBAFF mRNA levels in patients with IIM, compared to controls or patients with SLE, were significantly increased. CD20 mRNA levels in patients with IIM were greater compared to controls. No significant differences were found in APRIL mRNA expression between patients with IIM and controls.
We next sought to evaluate the expression of receptors for BAFF and APRIL (BAFF-R, BCMA, and TACI). Compared to TACI mRNA levels in patients with SLE, those in patients with IIM were increased (p = 0.001). However, neither BAFF-R (p = 0.47; p = 0.13) nor BCMA (p = 0.13; p = 0.61) expression in IIM subjects differed significantly compared to those in patients with SLE and healthy controls, respectively.
Correlation of BAFF pathway expression with disease activity
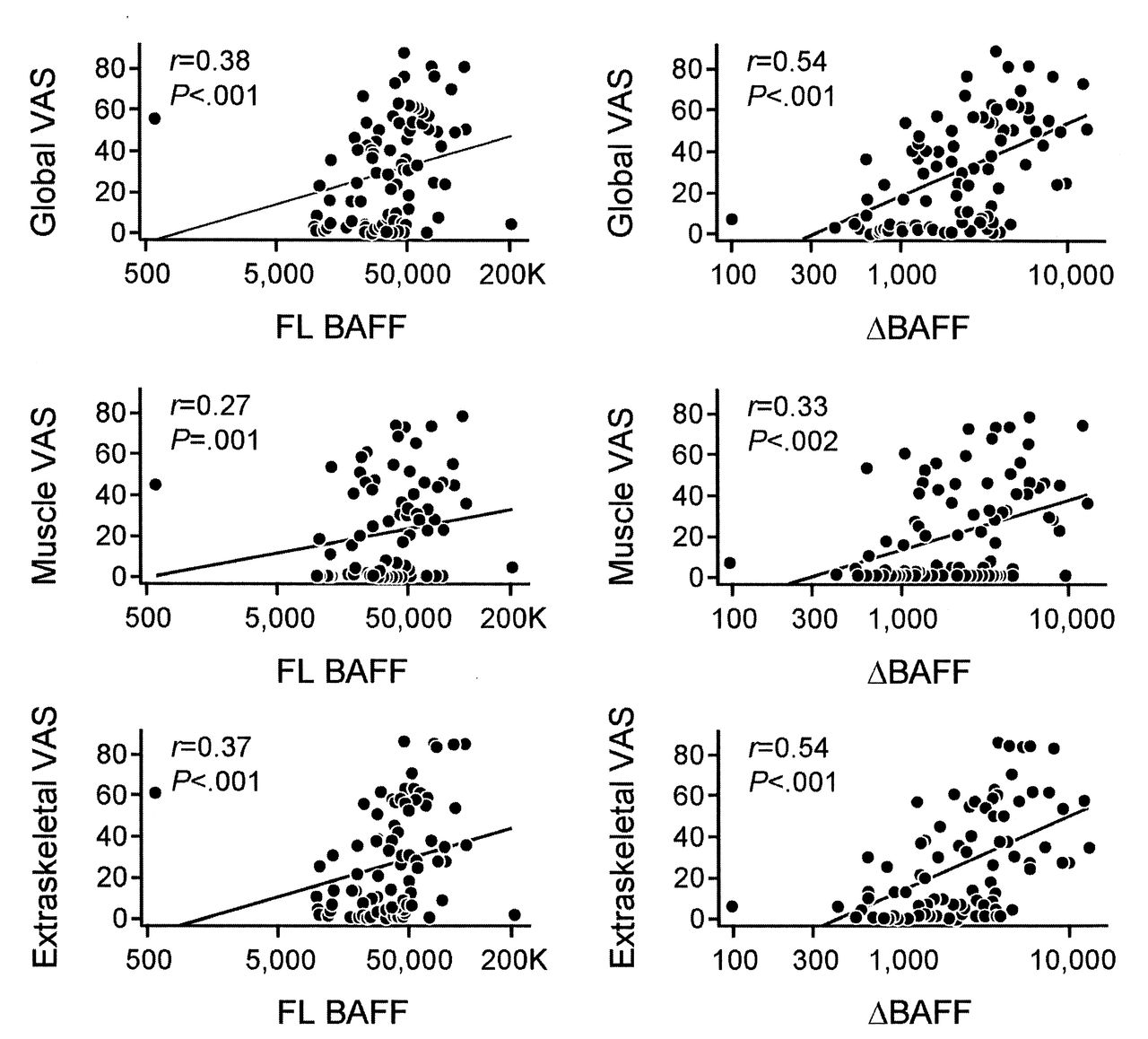
We found a positive correlation between BAFF and ΔBAFF expression and all 3 disease activity measures, including the extraskeletal disease activity score (r = 0.37, p < 0.001; and r = 0.54, p < 0.001, respectively); the VAS for muscle disease activity (r = 0.27, p = 0.01; and r = 0.33, p = 0.002, respectively); and the global VAS for disease activity (r = 0.38, p < 0.001; and r = 0.54, p < 0.001, respectively; Figure 2). Notably, ΔBAFF levels correlated more strongly than BAFF levels with disease activity measures. These data suggest that ΔBAFF might be a stronger predictor of disease activity in patients with IIM than in BAFF alone.
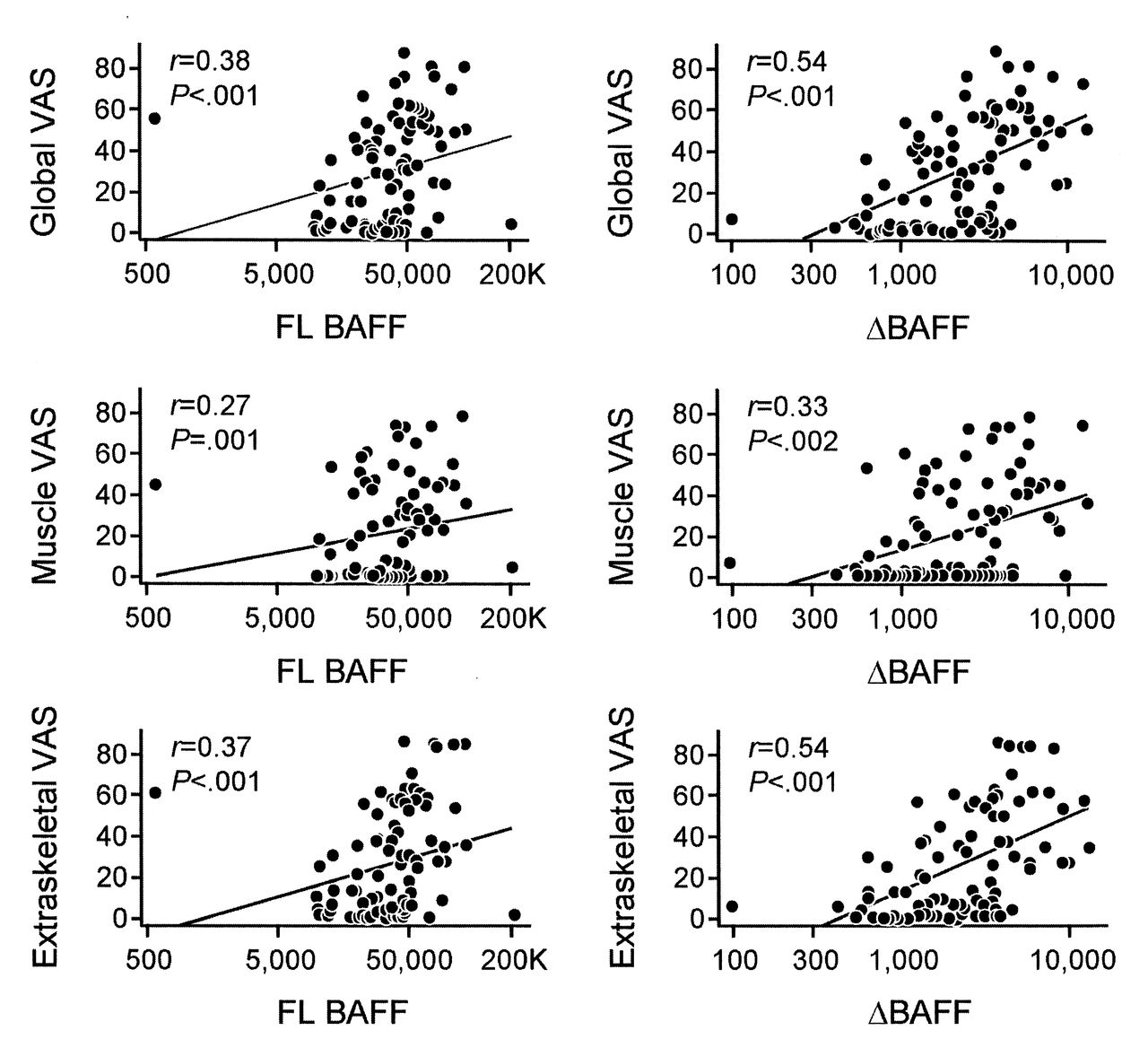
Correlation scatterplots of full-length B cell-activating factor belonging to the TNF family (FL BAFF) and ΔBAFF messenger (mRNA) expression in patients with idiopathic inflammatory myopathies by disease activity measure. FL BAFF and ΔBAFF mRNA expression in patients with the myositis disease activity core set measures established by International Myositis Assessment and Clinical Studies included global activity visual analog scale (VAS; upper panels), muscle VAS (middle panels), and extraskeletal VAS (lower panels). Y axes represent a VAS (0–100), X axes represent absolute copy numbers. A positive correlation was found between FL BAFF and ΔBAFF mRNA expression and all 3 disease activity measures.
To estimate the potential inhibition of BAFF by ΔBAFF, we calculated their relative ratios. The relative ratio of BAFF/ΔBAFF mRNA expression increased both in patients with IIM and in patients with SLE compared to that in healthy controls. Notwithstanding, the median mRNA copy number of BAFF/ΔBAFF was higher in patients with SLE (median 52, range 1.7–30,165) than in healthy controls (median 8.4, range 2.5–58.6; p < 0.001). In contrast, patients with IIM had a median BAFF/ΔBAFF mRNA expression of 15.2 (range 0.1–1457.9; p < 0.001), which suggests that in patients with SLE, decreased ΔBAFF may lead, in part, to BAFF overexpression. In patients with IIM, the relative ratio of BAFF/ΔBAFF was inversely correlated with the extramuscular disease activity score (r = −0.23, p = 0.03) and with the global disease activity score (r = −0.22, p = 0.04). In patients with SLE, BAFF/ΔBAFF was not correlated with disease activity, but it was marginally correlated with disease duration (p = 0.05). The BAFF/APRIL ratio was also increased in patients with IIM compared to that in patients with SLE (p < 0.001) and in healthy controls (p < 0.001). Patients with IIM had a median mRNA copy number of 5.2 (range 0.1–3562.1), compared to 1.6 (range 0.0–4895.9) in patients with SLE and 2.6 (range 0.8–5.8) in healthy controls. A significant correlation was observed between the BAFF/APRIL ratio and the extramuscular disease activity score (r = 0.33, p = 0.002), the muscle disease activity VAS score (r = 0.27, p = 0.01), and the global disease activity score (r = 0.31, p = 0.003).
Correlation of BAFF pathway expression with autoantibodies
In patients with IIM, BAFF and ΔBAFF levels and the relative ratio of BAFF/APRIL were significantly higher in persons with at least 1 anti-RNA binding protein (RBP) autoantibody (Table 3) than in persons without any of these autoantibodies. In addition, IIM patients with at least 1 RBP autoantibody had a significantly lower BAFF/ΔBAFF relative ratio (p = 0.02). Further, most of the IIM patients with at least 1 RBP antibody had anti-SSA/Ro 52-kDa present, which was associated with significantly higher relative ratios of mRNA levels of BAFF (p = 0.01), ΔBAFF (p < 0.001), BAFF/APRIL (p = 0.001), and BCMA/BAFF-R (p = 0.008). The presence of the anti-SSA/Ro 60-kDa autoantibody was associated with significantly higher mRNA levels of ΔBAFF (p < 0.001), whereas the presence of anti-Jo-1 autoantibody was associated with higher levels of the relative ratio of BAFF/APRIL (p = 0.006) and was marginally correlated with APRIL (p = 0.06).
Differences in BAFF pathway expression in patients with idiopathic inflammatory myopathies, with or without at least 1 anti-RNA binding protein (RBP) autoantibody. Values are median (range) unless otherwise indicated.
Correlation of BAFF pathway expression with medication use in patients with IIM
Some studies have shown that treatment with corticosteroids induces a marked decrease in BAFF in patients with SLE35 and in patients with immune thrombocytopenia36. We have analyzed the association between steroid treatment and the expression of these ligands/receptors. Patients with IIM who were not receiving steroids when BAFF was measured (n = 42) had significantly higher mRNA levels of BAFF-R (median mRNA copy number 2051.8, range 251.2–7811.7), BCMA (median mRNA copy number 475.7, range 3.0–21,373.0), and CD20 (median mRNA copy number 65,224, range 9680.4–448,402.0) compared to patients treated with steroids (BAFF-R median mRNA copy number 1115.2, range 26.2–13,579.0; BCMA median mRNA copy number 296.7, range 0.9–10,273.0; and CD20 median mRNA copy number 48,019, range 2270.7–229,960.0; p < 0.001, p = 0.03, and p = 0.01, respectively, adjusted for disease activity).
TACI mRNA expression was also higher in patients not treated with steroids (p = 0.04, adjusted for disease activity). However, BAFF, ΔBAFF, and APRIL mRNA levels showed no significant differences by use or nonuse of steroids (p = 0.81, p = 0.16, and p = 0.20, respectively, adjusted for disease activity). The BAFF/ΔBAFF relative ratio was higher in patients treated with steroids than in those not on steroids (median mRNA copy number 13.3, range 0.1–1457.9, vs median mRNA copy number 17.2, range 3.3–582.4; p = 0.02, adjusted for disease activity). Similarly, patients with IIM who were taking any DMARD at the time of blood draw had lower levels of BAFF, ΔBAFF, BCMA, and CD20 expression (p = 0.01, p = 0.02, p = 0.005, and p = 0.045, respectively, adjusted for disease activity). These data suggest that steroids may play a role in the BAFF receptor-signaling pathway; however, decreased BAFF and ΔBAFF expression in patients treated with DMARD suggests that steroids are not the only factor involved in reducing BAFF levels in patients with IIM.
DISCUSSION
BAFF and APRIL are TNF superfamily members known to be important for B cell development, maturation, and survival. There is growing evidence that BAFF and APRIL are involved somehow in the establishment and/or maintenance of systemic autoimmune diseases. In general, BAFF and APRIL may promote autoimmunity by increasing the survival of autoreactive B cells and plasma cells. BAFF-transgenic mice have been found to develop B cell hyperplasia and the deposition in tissue of immunoglobulin G and A autoantibodies, much as occurs in SLE in humans37. However, when autoimmune mice were treated with the soluble decoy receptor that blocks BAFF and APRIL (atacicept, formerly TACI-Ig), they had measurable depletion of peripheral B cells, with resultant alleviation of the signs and symptoms of autoimmune disease38.
The presence of elevated levels of BAFF and APRIL in patients with autoimmune diseases such as SLE, rheumatoid arthritis, and Sjögren syndrome suggests that these cytokines may play a role in human autoimmunity22,23,24. In contrast, little is known about the involvement of BAFF and APRIL in the pathogenesis of IIM. Previous studies have documented elevated serum levels of BAFF in patients with IIM25,28, which suggests that BAFF may affect the development of autoimmunity. However, whether a ligand is secreted or is membrane-bound has profound biologic ramifications. For instance, TNF-α has different (and antagonistic) biologic effects when it is a membrane-bound versus a soluble ligand39. Moreover, the anti-TNF agents infliximab, adalimumab, and etanercept bind to and neutralize soluble TNF-α but produce different effects on transmembrane TNF-α-producing cells40. In contrast to the action of TNF-α, the functional differences between transmembrane and soluble BAFF are unclear. Further, the aggregation of soluble BAFF into larger oligomeric states can also alter ligand activity41. Nonetheless, the correlation between serum BAFF protein levels and disease activity in human autoimmune diseases seems modest at best. Collins, et al42 has shown that serum BAFF correlates significantly with disease activity in patients with SLE, whereas BAFF mRNA is more closely associated with serum immunoglobulin levels and disease activity scores than with BAFF protein levels. We found that BAFF mRNA expression in PBMC was significantly elevated in patients with IIM and that this elevation correlated with clinical disease activity as assessed by the IMACS core set measures.
An important finding of our study is a significant increase in mRNA transcripts of inhibitory ΔBAFF, similar to those in FL BAFF, in patients with IIM compared to patients with SLE and healthy controls. In patients with SLE, ΔBAFF mRNA transcripts were significantly lower than in controls, resulting in a significantly increased BAFF/ΔBAFF ratio, particularly in SLE patients with active disease. Functionally, our findings may be important, because ΔBAFF has been reported to form heterotrimers with BAFF15, which might restrain the effects of BAFF and thereby regulate B lymphocyte homeostasis. Importantly, ΔBAFF transgenic mice have a reduced B cell pool, a suboptimal antibody response to T cell-dependent antigens and more stringent selection of their B cell repertoire16. We hypothesized that the activation of innate immunity and type 1 interferon may increase ΔBAFF, but to a lesser extent than BAFF. In that situation, ΔBAFF upregulation may be insufficient to inhibit excess BAFF (as illustrated by increased relative ratios of BAFF/ΔBAFF), which may perpetuate autoimmunity. Hence, the regulation of BAFF alternative splicing might be a complementary approach to anti-BAFF in developing strategies to combat autoimmune diseases. Notably, ΔBAFF correlated more strongly with disease activity in both patients with IIM and patients with SLE, which suggests that ΔBAFF may be a stronger predictor of disease activity than BAFF alone; however, few data are available on the cellular origin of ΔBAFF and its physiologic importance. Because our PCR experiments amplified BAFF and ΔBAFF from PBMC, our results do not fully exclude the possibility of variability in ΔBAFF expression in monocyte and lymphocyte subsets.
In line with these results, we also found an increase in B cells (as assessed by CD20 expression) in patients with IIM compared to healthy controls. The mechanism controlling the homeostatic level of BAFF is unknown, but it may have a direct effect on the number and composition of the B cell pool. In mouse models, transgenic overexpression of BAFF results in an increased number of mature B cells and subsequent development of autoimmune disorders20. These data suggest that alterations in BAFF levels in patients with IIM may significantly affect B cells, which may contribute to the development of autoimmunity.
The mechanism of action of BAFF and APRIL remains poorly understood, in part because of the complexity introduced by multiple receptors. We found TACI (but not BAFF-R) to be overexpressed in patients with IIM compared to patients with SLE and healthy controls; TACI mRNA expression was particularly higher in patients with IIM who were not treated with steroids. As a receptor for BAFF, TACI is mainly involved in B cell responses to T cell-independent antigens, and it is quickly induced upon in vitro B cell activation43. Studies have revealed an interesting relationship between TACI expression and Toll-like receptor (TLR) function. Groom, et al44 reported that in BAFF-transgenic mice, BAFF promoted the expression in B cells of the TLR 7 and 9. This activation strongly upregulated the expression of TACI, which in turn controlled the production of proinflammatory autoantibodies. TLR 7 and 9 proteins have been found in inflamed muscle tissue from patients with IIM, particularly on plasma cell-rich infiltrates45. In our cohort of patients with IIM, the high levels of TACI expression might reliably indicate a BAFF-binding receptor. Thus, the upregulation of TACI in IIM patients with elevated levels of BAFF could play a pathogenic role in the TLR-mediated production of autoantibodies.
Interestingly, BAFF and APRIL have been implicated in antibody production. By binding to their receptors and signaling for the activation of nuclear factor-κB(NF-κB), they regulate multiple aspects of T cell-dependent and T cell-independent antibody production, such as class switching, somatic hypermutation, plasmacytoid differentiation, and tolerance46. We found that BAFF and ΔBAFF transcript levels correlated with the presence of autoantibodies to SSA/Ro, SSB/La, and any RNA-binding protein in patients with IIM, whereas anti-Jo-1 correlated only with the BAFF/APRIL relative ratio. Notably, our data showed a clear correlation of BAFF and ΔBAFF expression with anti-SSA/Ro 52-kDa and/or anti-SSA/Ro 60-kDa-positive antibodies compared to that in healthy controls or in IIM patients without antibodies against RNA-binding protein. The relative ratios of BAFF/ΔBAFF were unexpectedly decreased significantly in patients with IIM who had RBP autoantibodies. In contrast, we found an inverse correlation of APRIL and antibody production as evidenced by APRIL expression alone and by APRIL in relation to BAFF in patients with anti-SSA and anti-Jo-1 antibodies. Overall, our results are consistent with those in other reports47 and they further support an emerging consensus for BAFF as a modulator, either primarily or secondarily, of the level of antibody production in autoimmune diseases.
Our analysis presents the mRNA levels of B cell activation ligands/receptors in the largest cohort of patients with IIM prior to any drug treatment described to date. Our results are consistent with those of other authors, in that we found that patients with IIM who were not treated with corticosteroids or DMARD had significant increases in BAFF and ΔBAFF. Importantly, BAFF-R and BCMA were significantly higher in IIM patients without any prior steroid treatment than in steroid-treated patients with IIM. Similarly, we found an increased number of B cells in patients not treated with steroids. BAFF and APRIL receptors are expressed mainly on mature B cells, but also on T cells18,42. The BAFF/APRIL-system receptors trigger activation of NF-κB, signaling BAFF-system receptors that affect the survival and basal proliferation of B cells41. Our findings therefore suggest that corticosteroids may play a role in downregulating BAFF-R and BCMA expression in patients with IIM, possibly through inhibition of the NF-κB pathway. Thus, it will be of interest to examine whether BAFF-R reduction by NF-κB blocking might be another choice for controlling B cells in patients with autoimmune diseases such as IIM.
Acknowledgment
We thank the Biospecimens Accessioning and Processing Core and the Gene Expression Core of the Advanced Genomics Technology Center at the Mayo Clinic, Rochester, Minnesota, USA, for assistance with sample collection and quantitative real-time polymerase chain reaction, respectively.
Footnotes
-
Supported by grants from the Arthritis Foundation and the State of Minnesota Partnership for Biotechnology and Medical Genomics.
- Accepted for publication November 8, 2012.








{kind=link}
{kind=link}