
Abstract
Objective. Our previous studies indicated that CD40-mediated Fas-dependent apoptosis is important for the glandular destruction of Sjögren’s syndrome (SS), although other immune and nonimmune mechanisms are also involved in exocrine dysfunction. We investigated the roles of p38 mitogen-activated protein kinase (p38MAPK) and nuclear factor-κB (NF-κB) in salivary epithelial cell death in SS.
Methods. Expression of p38, phosphorylated p38 (pp38), and IκB-α was examined by Western blotting upon CD40 ligation. Activity of NF-κB induced by anti-CD40 monoclonal antibody (mAb) was examined by electrophoretic mobility shift assay (EMSA) and Western blotting. Expression of Fas was analyzed by flow cytometry and Western blotting with or without the p38-specific inhibitor SB203580 or the NF-κB-specific inhibitor caffeic acid phenethyl ester (CAPE). Induction of apoptosis in salivary epithelial cells was examined by DNA fragmentation and terminal deoxynucleotidyl transferase-mediated dUTP nick end-labeling assay. Expression of phosphorylated p38MAPK and NF-κB was measured by immunohistochemistry.
Results. pp38MAPK and NF-κB p65 were predominantly expressed in the ductal and acinar epithelium adjacent to lymphoid infiltrates of SS salivary gland by immunohistochemistry. CD40 ligation strongly enhanced p38MAPK and NF-κB activity by EMSA and Western blotting in cultured salivary epithelial cells. Treatment of cells with anti-CD40 mAb resulted in significantly upregulated Fas expression and induction of Fas-dependent apoptosis. Inhibition of p38MAPK and NF-κB activity by SB203580 and/or CAPE reduced Fas expression and apoptosis in salivary epithelial cells, establishing p38MAPK and NF-κB as proapoptotic factors in this context.
Conclusion. CD40 ligation plays an important role in activation of p38MAPK, NF-κB, and Fas molecules to initiate proapoptotic signaling. p38MAPK and NF-κB collaborate in regulation of proapoptotic signaling in CD40-mediated Fas-dependent apoptosis in salivary epithelial cells.
- SJÖGREN’S SYNDROME
- P38MAPK
- NUCLEAR FACTOR-κB
- APOPTOSIS
- CD40
Sjögren’s syndrome (SS) is an autoimmune disorder characterized by a progressive, immune-mediated destruction of exocrinopathy such as the lacrimal and salivary glands, leading to ocular and oral dryness1,2,3,4. The pathogenesis of the disease has not been fully elucidated, but accelerated apoptosis has been considered to have an important function in the pathogenesis of SS, although researchers have proposed that other immune and nonimmune-mediated salivary gland dysfunction may be involved in glandular destruction and atrophy5,6,7,8,9. Our previous studies have shown that salivary epithelial cells isolated from patients with SS undergo apoptosis through coactivation of Fas and CD40 pathways; CD40 ligation provides a potent proapoptotic signal10. However, the precise mechanism mediating CD40 ligation-induced apoptosis in SS salivary epithelial cells is unknown.
p38 mitogen-activated protein kinase (p38MAPK) has been implicated in both mitogenic and apoptotic responses, as overexpression of kinases that can activate p38MAPK resulted in the induction of apoptosis11,12. p38MAPK is phosphorylated (phosphorylated p38: pp38) in response to a variety of stress stimuli, such as heat, ultraviolet irradiation, osmotic shock, and proinflammatory cytokines including interleukin 1ß (IL-1ß), tumor necrosis factor-α (TNF-α), and CD4011,12,13,14. CD40 ligation leads to activation of p38MAPK, and p38MAPK is thought to be involved in the pathogenesis of SS15. This prompted us to examine whether salivary epithelial cell death induced by Fas and CD40 could use the distinct pathway of p38MAPK.
On the other hand, the mammalian Rel/NF-κB family plays a key role in the regulation of immune responses, inflammatory reactions, cell proliferation, and apoptosis16,17. NF-κB activation has been viewed as a cytoprotective event based on its ability to upregulate antiapoptotic proteins18,19. However, there have been reports that the NF-κB signal also plays a role in proapoptotic signaling. For example, NF-κB induces cell death following T cell receptor engagement or exposure to DNA-damaging agents, upregulation of p53 expression, and ceramide activation in osteoblasts20,21,22,23,24. These findings suggest that whether the function of NF-κB is proapoptotic or antiapoptotic depends on the cell type, extent of NF-κB activation, and nature of the apoptotic signals.
We observed that p38MAPK and NF-κB function as downstream molecules involved in CD40 ligation-induced apoptosis of salivary epithelial cell death. The results of our study may define novel target molecules for controlling salivary epithelial cell death in SS.
MATERIALS AND METHODS
Patients and controls
Twenty-eight patients (25 women, 3 men; mean ± SD age 64.2 ± 9.8 yrs, range 28–81 yrs) fulfilling the European classification criteria for primary SS25 were recruited from the First Affiliated Hospital, China Medical University, Shenyang. The control group consisted of 20 patients matched for age and sex (18 women, 2 men) who did not fulfill the criteria above. Biopsy of the minor salivary gland was performed as a routine part of the diagnostic evaluation of SS. All experimental protocols were approved by the Independent Ethics Committee of First Affiliated Hospital, China Medical University.
Reagents
Mouse monoclonal antibody (mAb) to human Fas (clone CH11) was purchased from Medical & Biological Laboratories (Nagoya, Japan). Purified mouse mAb to human CD40 (clone 5C3) and isotype controls (mouse IgG and IgM) were obtained from BD PharMingen (San Diego, CA, USA). Rabbit polyclonal antibodies against human pp38MAPK and “total” p38MAPK, and mouse mAb against human phosphorylated IκB-α (clone 5A5) were purchased from Cell Signaling Technology (Danvers, MA, USA). Mouse mAb to human NF-κB p65 (clone NF-12), the p38 inhibitor SB203580, the NF-κB inhibitor caffeic acid phenethyl ester (CAPE), and mouse mAb to human ß-actin (clone AC-15) were purchased from Sigma. Chimeric antagonistic mAb to human CD40 (ch5D12) was kindly provided by Tanox Pharma B.V. (Amsterdam, The Netherlands)26.
Primary cultures of salivary gland epithelial cells
Epithelial cells from salivary gland tissues were cultured according to modified methods as described10. Briefly, tissues were minced into fragments of ∼1 mm3. One fragment of each tissue sample was placed in a 24-well collagen type-1 coated plate (Iwaki, Tokyo, Japan) and cultured in defined keratinocyte-SFM with growth supplement, 0.4 μg/ml hydrocortisone, 100 U/ml penicillin, 100 μg/ml streptomycin, and 25 μg/ml bovine pituitary extract. Epithelial cell outgrowth from the explant was observed after 1 to 2 weeks. After reaching confluence, the cells in the monolayer were trypsinized and subcultured. Cultured epithelial cells were used for experiments when they reached 90% confluence. The epithelial origin of cultured salivary epithelial cells was verified routinely by morphology, as well as by the uniform and consistent expression of epithelial markers and the absence of markers indicative of lymphoid/monocytoid cells.
Flow cytometry
After stimulation, cultured salivary epithelial cells were subsequently incubated with 1:20 FITC-conjugated rabbit anti-mouse Ig mAb (Dako, Glostrup, Denmark) for 1 h at 4°C. The cells were washed twice with phosphate buffered saline (PBS) and analyzed by flow cytometry. Negative control cells were incubated with isotype-matched antibody. The expression of surface Fas was calculated from the mean fluorescence intensity using the CellQuest program (BD Biosciences, Franklin, NJ, USA).
Assessment of apoptosis
This assessment was done with 2 assays: (1) DNA fragmentation assay: all experimental procedures were performed according to a described protocol10; and (2) terminal deoxynucleotidyl transferase-mediated dUTP nick end-labeling assay.
The experiments were performed using a Mebstain Apoptosis Direct Kit (Medical & Biological Laboratories). Apoptotic cells were determined by flow cytometry.
Western blotting
After stimulation, 1 × 107 human salivary epithelial cells were lysed in Triton lysis buffer (20 mM Tris, pH 7.4, 137 mM NaCl, 2 mM EDTA, 10% glycerol, and 1% Triton X-100) containing protease inhibitor cocktail (Sigma) and phosphatase inhibitors (20 mM ß-glycerophosphate, 10 mM 4-nitrophenylphosphate, and 50 μM sodium vanadate; all from Sigma). Samples of 40 μg of nuclear protein (to detect NF-κB RelA p65) or cytoplasmic protein (to detect p38, pp38, phosphorylated IκB-α, and Fas) were subjected to 10% SDS-polyacrylamide gel electrophoresis (SDS-PAGE), and transferred onto polyvinylidene difluoride membranes (Hybond-P; Amersham-Pharmacia, Little Chalfont, UK). Membranes were incubated 1 h with 5% nonfat dried milk to block nonspecific binding sites, then Western blotting analyses were performed using rabbit polyclonal anti-human pp38MAPK and “total” p38MAPK (1:1000 dilution), 20 μg/ml of mAb to human Fas (clone CH11), mouse mAb to human NF-κB RelA (p65) (clone NF-12, 1:1000 dilution), and mouse mAb to human phosphorylated IκB-α (1:1000 dilution) for 1 h. Blots were subsequently stripped with 6.25 mM Tris, 2% SDS, and 100 mM 2-mercaptoethanol for 30 min at 50°C, washed 1 h in TBS-Tween, and reprobed with mAb to human ß-actin (clone AC-15, 1:10,000) for normalization. The blots were washed 3 times with PBS containing 0.05% Tween 20 and incubated with goat anti-mouse or donkey anti-rabbit antibody coupled to horseradish peroxidase (1:25,000; Amersham-Pharmacia). The enhanced chemiluminescence system (Amersham-Pharmacia) was used to develop the blots in accord with the manufacturer’s instructions. The expected sizes of the bands on Western blots were 38 kDa for p38 and pp38, 65 kDa for NF-κB RelA (p65), 36 kDa for phosphorylated IκB-α, 45 kDa for Fas, and 42 kDa for ß-actin.
Preparation of nuclear protein and electrophoretic mobility shift assay (EMSA)
Nuclear protein from 1 × 107 human salivary epithelial cells was prepared using NE-PER (Nuclear and Cytoplasmic Extraction Reagents; Pierce Biotechnology, Rockford, IL, USA). EMSA was performed using a commercial LightShift Chemiluminescent EMSA Kit (Pierce). NF-κB consensus oligonucleotide (5′-AGT TGA GGG GAC TTT CCC AGG C-3′; Santa Cruz Biotechnology, Santa Cruz, CA, USA) was labeled with biotin (Pierce). All procedures were performed in accord with the manufacturer’s instructions.
Immunohistochemistry
Deparaffinized sections of minor salivary glands 5 μm thick were processed for immunohistochemistry. After blocking with 3% dry milk and 5% bovine serum in PBS, sections were incubated with rabbit polyclonal antibodies against human pp38MAPK (1:200 dilution in PBS), and mouse mAb to human NF-κB p65 (1:75 dilution in PBS) for 12 h at 4°C. After washes in PBS, specimens were treated with peroxidase-conjugated antibodies against each immunoglobulin for 4 h at 4°C. After another wash in PBS, a peroxidase reaction was performed with 3,3′-diaminobenzidine. The sections were mounted in balsam and observed under light microscopy. Control slides were incubated with Tris buffered saline containing isotype-matched antibodies instead of the primary antibody. They were invariably negative (data not shown).
Statistical analysis
The data are expressed as mean ± SD. Statistical analysis was performed with Student’s t test or an ANOVA with a Bonferroni adjustment for multiple comparisons. P values < 0.05 were considered significant.
RESULTS
CD40 ligation activates p38MAPK in salivary epithelial cells
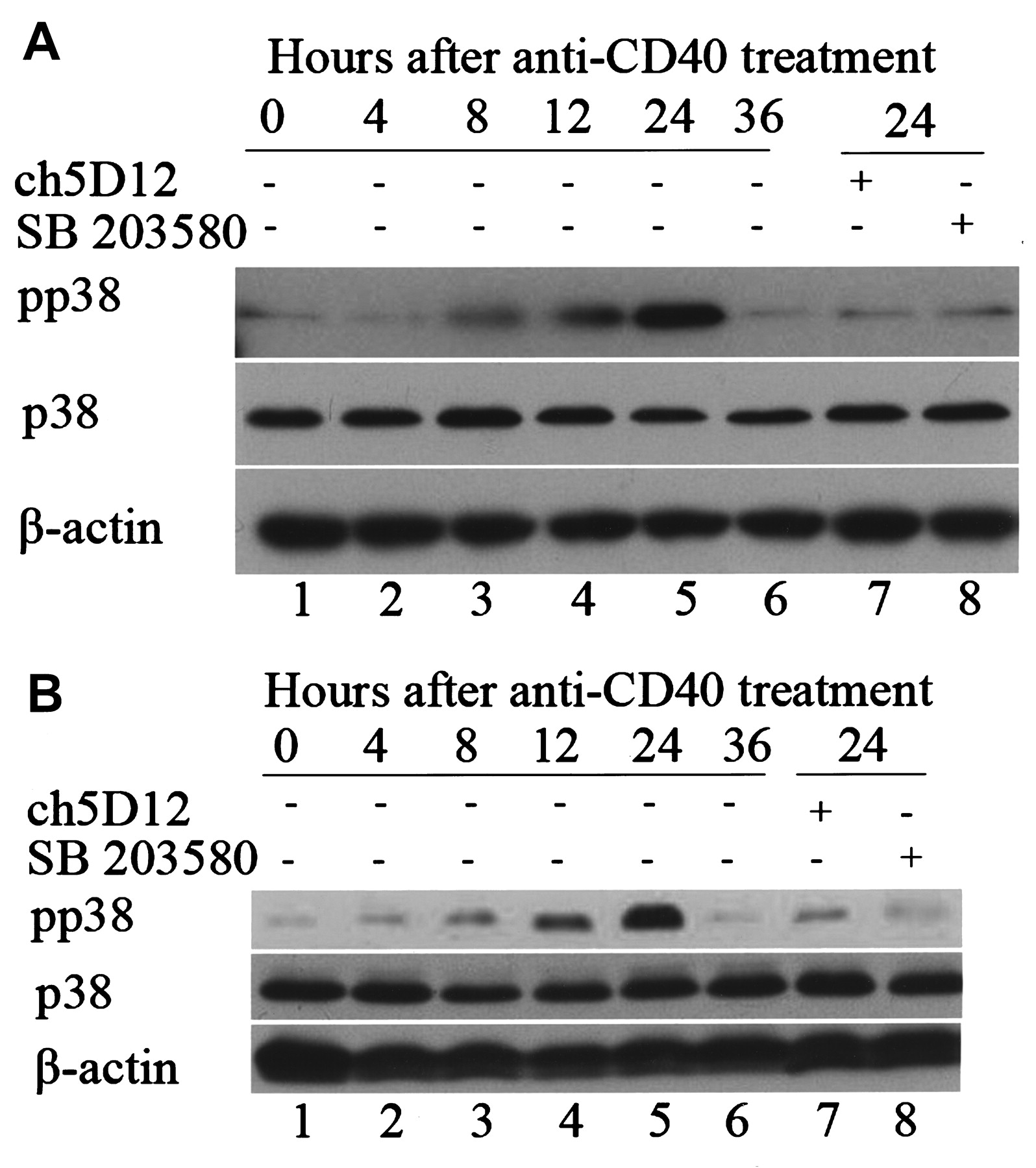
As shown in Figure 1A and 1B, CD40 ligation strongly increased p38MAPK phosphorylation in SS and normal control salivary epithelial cells. The phosphorylation of p38MAPK reached its maximal level after 24 h of CD40 treatment, which returned to baseline after 36 h. CD40 treatment up to 24 h had no effect on “total” p38MAPK expression in both SS and normal control salivary epithelial cells.

The effects of CD40 ligation on activation of the p38 mitogen-activated protein kinase (p38MAPK) pathway in cultured salivary epithelial cells. Sjögren’s syndrome (SS; A) and control salivary epithelial cells (B) were stimulated with anti-CD40 monoclonal antibody (mAb) or pretreated with antagonistic anti-CD40 mAb (ch5D12) or p38-specific inhibitor (SB203580) for the times indicated. Blots were probed for phosphorylated p38 (pp38, upper panel) and “total” p38 (p38, lower panel). Epithelial cells were incubated with medium alone (lane 1), or with anti-CD40 mAb (1 μg/ml) for 4 h (lane 2), 8 h (lane 3), 12 h (lane 4), 24 h (lane 5), or 36 h (lane 6). Cells were pretreated with antagonistic anti-CD40 mAb (ch5D12; 5 μg/ml) for 1 h before stimulation with anti-CD40 mAb for 24 h (lane 7). Cells were pretreated with SB203580 (10 μM) for 1 h before stimulation with anti-CD40 mAb for 24 h (lane 8). Results shown are representative of 12 independent experiments on cells derived from 6 different patients with SS and 6 different controls.
Further, CD40 ligation-induced p38MAPK phosphorylation was markedly inhibited by preincubation with antagonistic anti-CD40 mAb (ch5D12) or a p38MAPK-specific inhibitor, SB203580, in SS and control salivary epithelial cells (Figure 1A, 1B). Nevertheless, no difference was found in the expression of p38MAPK and pp38MAPK between SS and normal control cells (Figure 1A, 1B).
CD40 ligation induces NF-κB activation
As shown in Figure 2A, CD40 ligation induced a time-dependent increase in NF-κB activity in salivary epithelial cells. The maximal NF-κB activity induced by CD40 ligation was observed after 24 h treatment of cells with CD40 mAb, which returned to basal level at 36 h (Figure 2A, lanes 2–7). Binding of NF-κB was confirmed by supershifting of DNA-protein complexes with antibodies against NF-κB p65 (Figure 2A, lane 10). The specificity of the binding activity was demonstrated by coincubation with 100-fold molar excess of unlabeled NF-κB oligonucleotide probe (Figure 2A, lane 8), and a mutant sequence oligonucleotide probe (Figure 2A, lane 9). CD40 ligation-induced NF-κB activation was markedly inhibited in salivary epithelial cells when these cells were preincubated with antagonistic anti-CD40 mAb (ch5D12) or a specific inhibitor of NF-κB, CAPE (Figure 2A, lanes 11, 12).
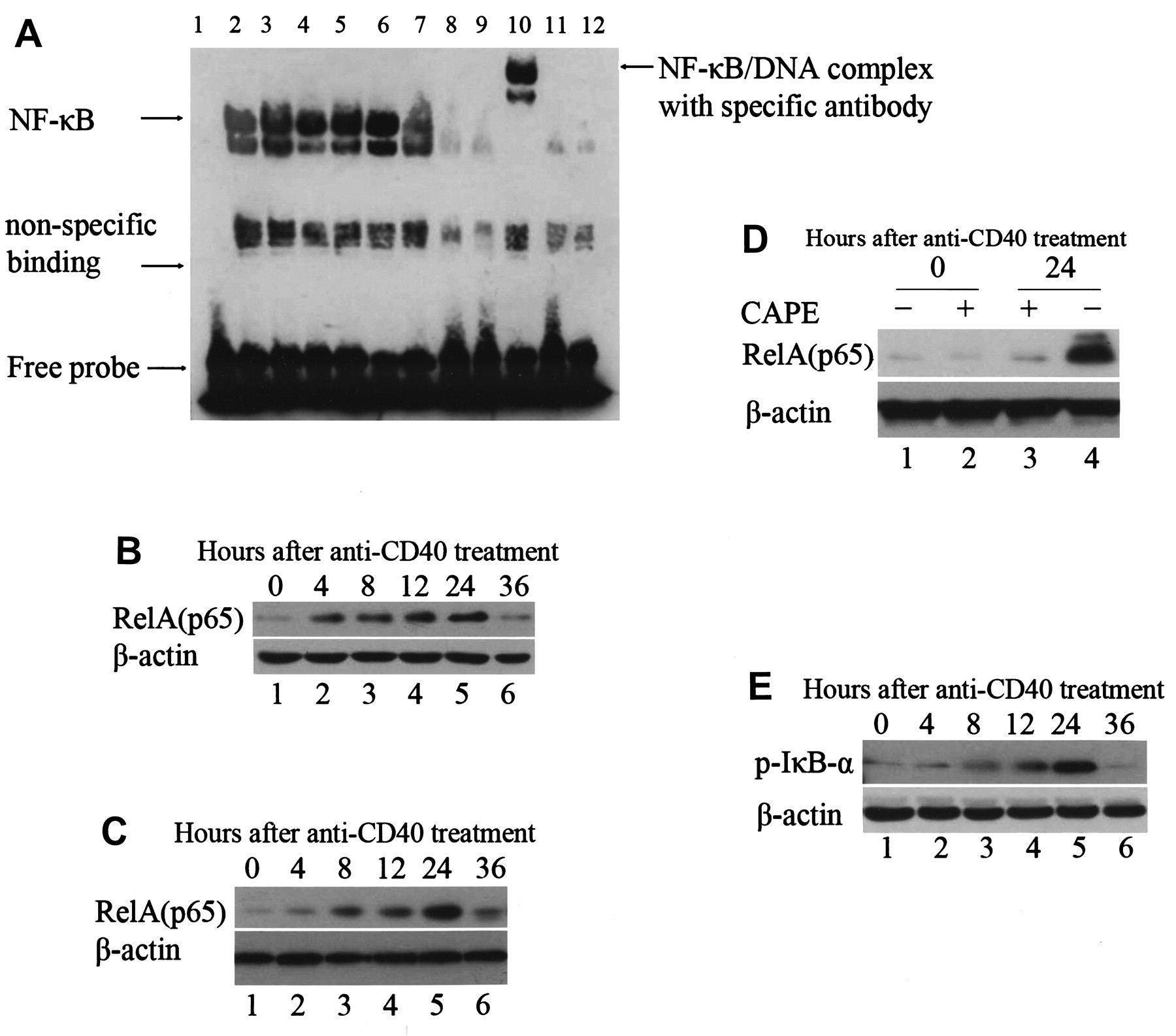
The effects of CD40 ligation on nuclear factor-κB (NF-κB) DNA-binding activity by electrophoretic mobility shift assay (EMSA) and Western blotting in salivary epithelial cells. (A) The effects of CD40 ligation on NF-κB DNA-binding activity by EMSA. Sjögren’s syndrome (SS) salivary epithelial cells were stimulated with anti-CD40 monoclonal antibody (mAb; 1 μg/ml) for the indicated times. Lane 1 shows biotin-labeled probe alone. Epithelial cells were incubated with medium alone (lane 2), anti-CD40 mAb for 4 h (lane 3), 8 h (lane 4), 12 h (lane 5), 24 h (lane 6), or 36 h (lane 7), cold competition with 100-fold excess of unlabeled probe (lane 8), a mutant NF-κB consensus probe (lane 9), supershift using anti-NF-κB p65 (lane 10), pretreated with NF-κB inhibitor caffeic acid phenethyl ester (CAPE; 25 μg/ml) for 1 h before stimulation with anti-CD40 mAb for 24 h (lane 11), or pretreated with antagonistic anti-CD40 mAb (ch5D12; 5 μg/ml) for 1 h before stimulation with anti-CD40 mAb for 24 h (lane 12). Results are representative of 6 independent experiments on cells derived from 6 different patients with SS. (B, C) The effects of CD40 ligation on NF-κB expression by Western blotting. SS (B) and control (C) salivary epithelial cells were stimulated with anti-CD40 (1 μg/ml) for the indicated times. Epithelial cells were incubated with medium alone (lane 1), anti-CD40 for 4 h (lane 2), 8 h (lane 3), 12 h (lane 4), 24 h (lane 5), or 36 h (lane 6). Results are representative of 12 independent experiments on cells derived from 6 different patients with SS and 6 different controls. (D) Specific inhibitory effects of CAPE on levels of NF-κB expression in SS salivary epithelial cells by Western blotting. Salivary epithelial cells were stimulated with or without CAPE (25 μg/ml) or anti-CD40 mAb (1 μg/ml) for the indicated times. Epithelial cells were incubated with medium alone (lane 1), CAPE for 1 h (lane 2), anti-CD40 mAb for 24 h, and then stimulated with CAPE for 1 h (lane 3) and anti-CD40 mAb for 24 h (lane 4). Results are representative of 6 independent experiments on cells derived from 6 different patients with SS. (E) Effects of CD40 ligation on levels of cytoplasmic phosphorylated IκB-α in SS salivary epithelial cells determined by Western blotting. Epithelial cells were incubated with medium alone (lane 1), anti-CD40 mAb (1 μg/ml) for 4 h (lane 2), 8 h (lane 3), 12 h (lane 4), 24 h (lane 5), or 36 h (lane 6). Results are representative of 6 independent experiments on cells derived from 6 different patients with SS.
In parallel with the EMSA results, CD40 ligation significantly increased RelA (p65) expression in the nuclear extracts of both SS and normal control salivary epithelial cells by Western blot (Figure 2B, 2C). These increases in RelA (p65) expression were detected after 4 h CD40 stimulation, which reached a maximal level at 24 h and returned to basal level after 36 h stimulation (Figure 2B, lanes 2–6). Further, CD40 ligation-induced expression of RelA (p65) did not differ between SS and normal cells (Figure 2B, 2C). This CD40 ligation-induced increase in nuclear RelA expression in salivary epithelial cells was markedly inhibited by CAPE (Figure 2D).
Similarly, we found that the expression level of cytoplasmic phosphorylated IκB-α in salivary epithelial cells increased at 4 h, reached a maximum at 24 h, and returned to basal levels at 36 h of CD40 stimulation (Figure 2E).
Increased susceptibility to Fas killing by CD40 is through p38MAPK and NF-κB
Flow cytometry and Western blot analyses were performed to examine the surface expression and total expression levels of Fas in salivary epithelial cells, respectively. As shown in Figure 3A and 3B, CD40 ligation induced similar levels of increases in surface expression of Fas in SS and control salivary epithelial cells, which was blocked by SB203580 and CAPE. As shown in Figure 3C and 3D, CD40 ligation-induced increase in total Fas expression in salivary epithelial cells was abolished in the presence of SB203580 and CAPE.
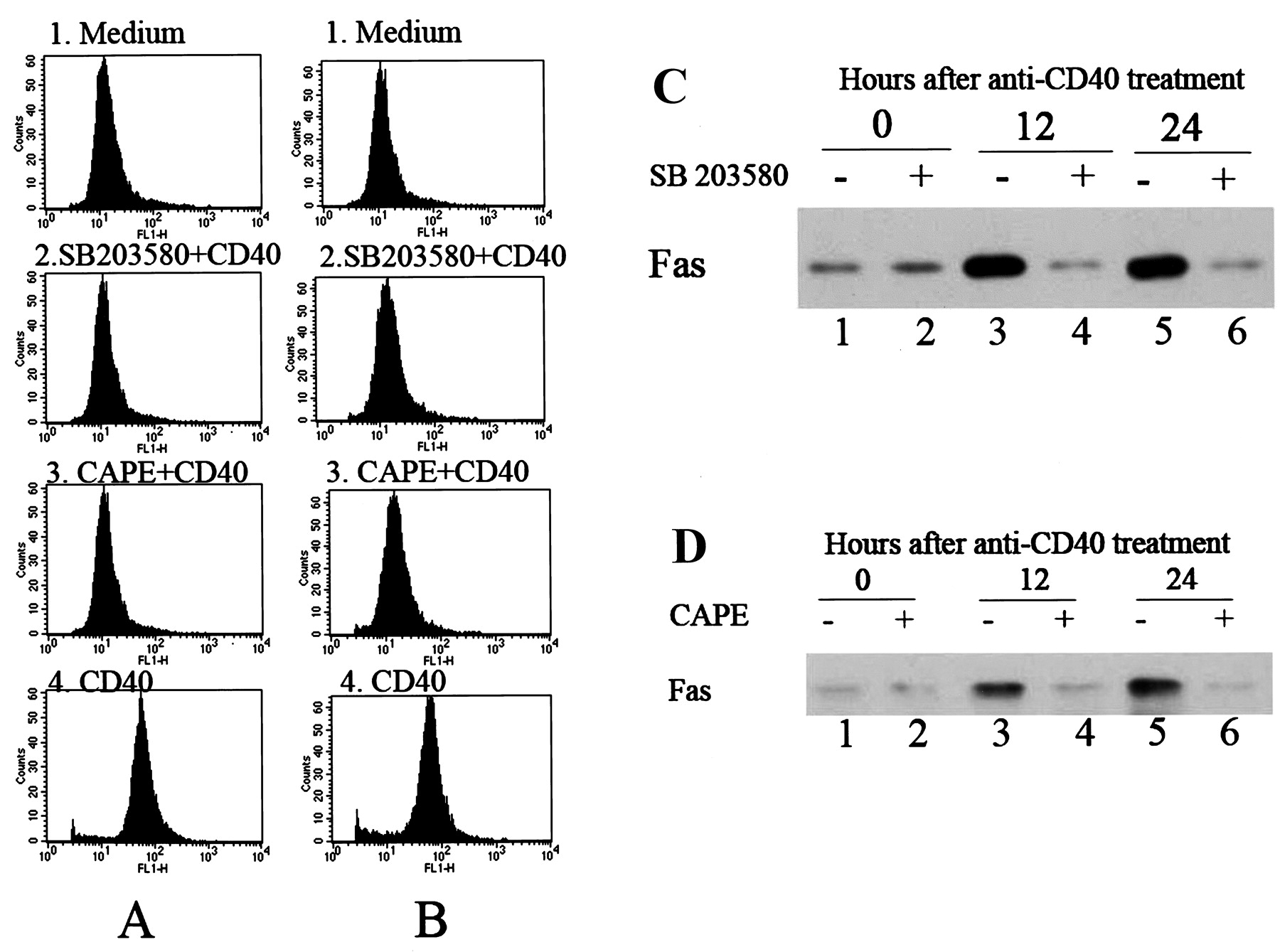
The inhibitory effects of the specific p38 mitogen-activated protein kinase (p38MAPK)-specific inhibitor SB203580 and the nuclear factor-κB (NF-κB) inhibitor caffeic acid phenethyl ester (CAPE) on CD40-induced Fas expression in salivary epithelial cells determined by flow cytometry and Western blotting. Sjögren’s syndrome (SS; A) and control (B) salivary epithelial cells were incubated with medium alone (panel 1), pretreated with SB203580 (10 μM) for 1 h before stimulation with anti-CD40 monoclonal antibody (mAb) for 24 h (panel 2), pretreated with CAPE (25 μg/ml) for 1 h before stimulation with anti-CD40 mAb for 24 h (panel 3), or stimulated with anti-CD40 mAb for 24 h (panel 4). Surface expression of Fas was analyzed by flow cytometry. Results are representative of 12 independent experiments on cells derived from 6 different patients with SS and 6 different controls. (C) SS salivary epithelial cells were incubated with medium alone (lane 1), treated with SB203580 for 1 h (lane 2), with anti-CD40 mAb for 12 h (lane 3), with SB203580 for 1 h before stimulation with anti-CD40 mAb for 12 h (lane 4), with anti-CD40 mAb for 24 h (lane 5), or with SB203580 for 1 h before stimulation with anti-CD40 mAb for 24 h (lane 6). Fas expression was determined by Western blotting. Results are representative of 6 independent experiments on cells derived from 6 different patients with SS. (D) SS salivary epithelial cells were incubated with medium alone (lane 1), treated with CAPE for 1 h (lane 2), with anti-CD40 mAb for 12 h (lane 3), with CAPE for 1 h before stimulation with anti-CD40 mAb for 12 h (lane 4), with anti-CD40 mAb for 24 h (lane 5), or with CAPE for 1 h before stimulation with anti-CD40 mAb for 24 h (lane 6). Fas expression was determined by Western blotting. Results are representative of 6 independent experiments on cells derived from 6 different patients with SS.
P38MAPK and NF-κB act synergistically to induce Fas-depen dent apoptosis in salivary epithelial cells
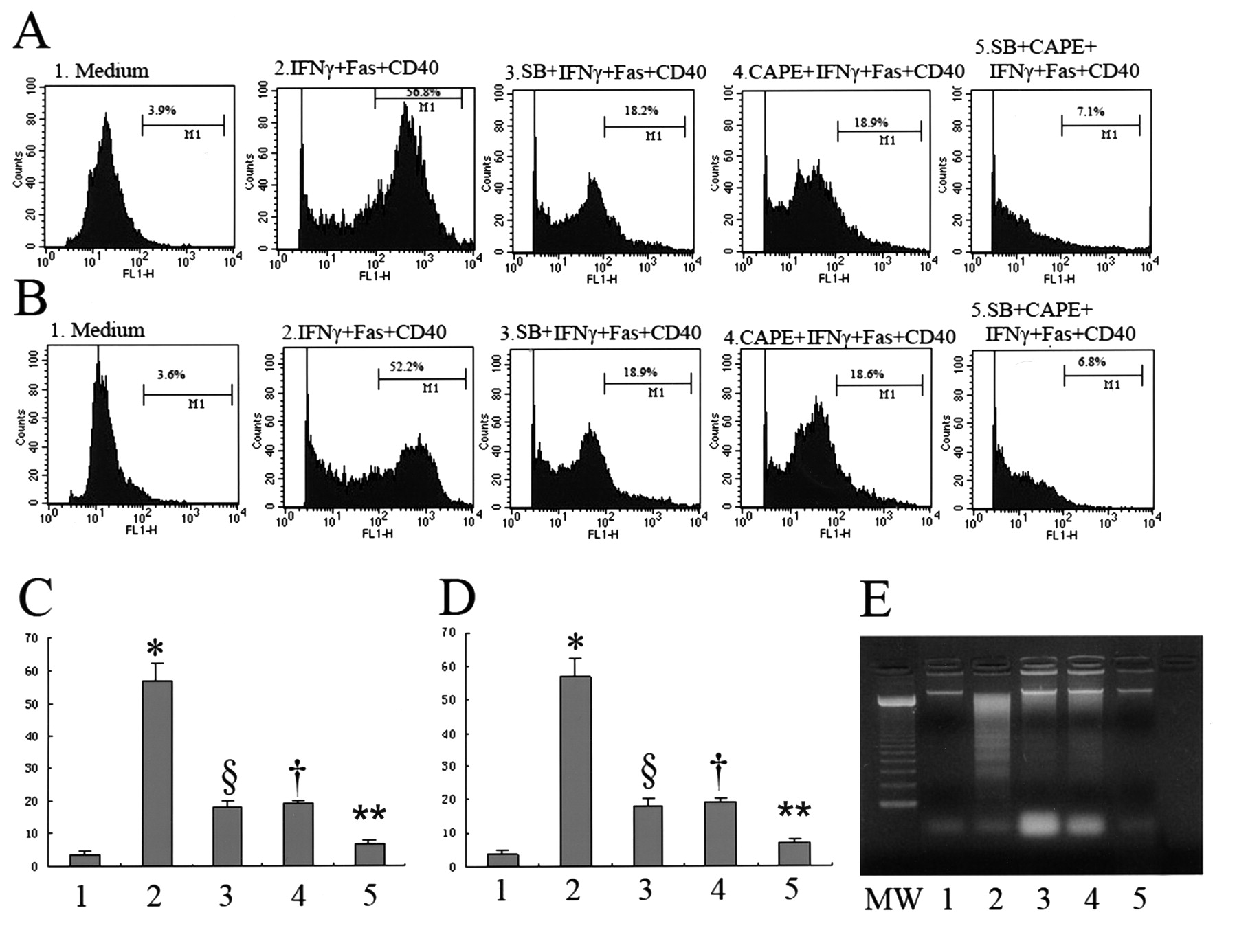
Preincubation of cells with 10 μM SB203580 or 25 μg/ml CAPE for 1 h before stimulation with interferon-γ (IFN-γ) and incubation with anti-CD40 mAb and anti-Fas mAb for 24 h resulted in a significant decrease in the number of apoptotic cells compared with cells treated with IFN-γ, anti-CD40 mAb, and anti-Fas mAb for 24 h (p < 0.01; Figure 4A–4E). In addition, preincubation of cells with 10 μM SB203580 and 25 μg/ml CAPE simultaneously for 1 h before stimulation with IFN-γ, and incubation with anti-CD40 mAb and anti-Fas mAb for 24 h resulted in a significant decrease in the number of apoptotic cells compared with cells pretreated with 10 μM SB203580 or 25 μg/ml CAPE, respectively (p < 0.01). These observations suggest that simultaneous activation of NF-κB and p38MAPK synergistically enhances the salivary epithelial cell apoptosis (Figure 4A–4E).

The inhibitory effects of p38 mitogen-activated protein kinase (p38MAPK) and the nuclear factor-κB (NF-κB)-specific inhibitors on apoptosis of salivary epithelial cells. Salivary epithelial cells were incubated with (1) medium alone, (2) interferon (IFN)-γ (1000 U/ml) for 72 h and anti-Fas monoclonal antibody (mAb; 1 μg/ml) and anti-CD40 mAb (1 μg/ml) for 24 h, (3) pretreated with p38-specific inhibitor (SB203580; 10 μM) for 1 h before incubation with IFN-γ (1000 U/ml) for 72 h, anti-Fas mAb (1 μg/ml) and anti-CD40 mAb (1 μg/ml) for 24 h, (4) pretreated with caffeic acid phenethyl ester (CAPE; 25 μg/ml) for 1 h before incubation with IFN-γ (1000 U/ml) for 72 h, anti-Fas mAb (1 μg/ml) and anti-CD40 mAb (1 μg/ml) for 24 h, (5) pretreated with SB203580 (10 μM) and CAPE (25 μg/ml) simultaneously for 1 h before incubation with IFN-γ (1000 U/ml) for 72 h, anti-Fas mAb (1 μg/ml) and anti-CD40 mAb (1 μg/ml) for 24 h. (A, B) Terminal deoxynucleotide transferase-mediated dUTP nick end-labeling (TUNEL) assay of apoptotic salivary epithelial cells using flow cytometry. Results are representative of 12 independent experiments on cells derived from 6 different patients with Sjögren’s syndrome (SS; A) and 6 different controls (B). Numbers in each box represent percentages of apoptotic cells. (C, D) Quantification of apoptotic cells in the TUNEL assay. Results are representative of 12 independent experiments on cells derived from 6 different patients with SS (C) and 6 different controls (D). Bars show mean ± SD. *p < 0.01 versus all other groups by ANOVA. §p < 0.01; †p < 0.01 versus group 5. **p < 0.01 versus group 2, 3, 4. (E) DNA fragmentation assay in SS salivary epithelial cells as assessed by agarose gel electrophoresis. Cells were lysed and fragmented DNA was isolated and electrophoresed. A 100-bp molecular weight marker is shown (MW). Results are representative of 6 independent experiments on cells derived from 6 different patients with SS.
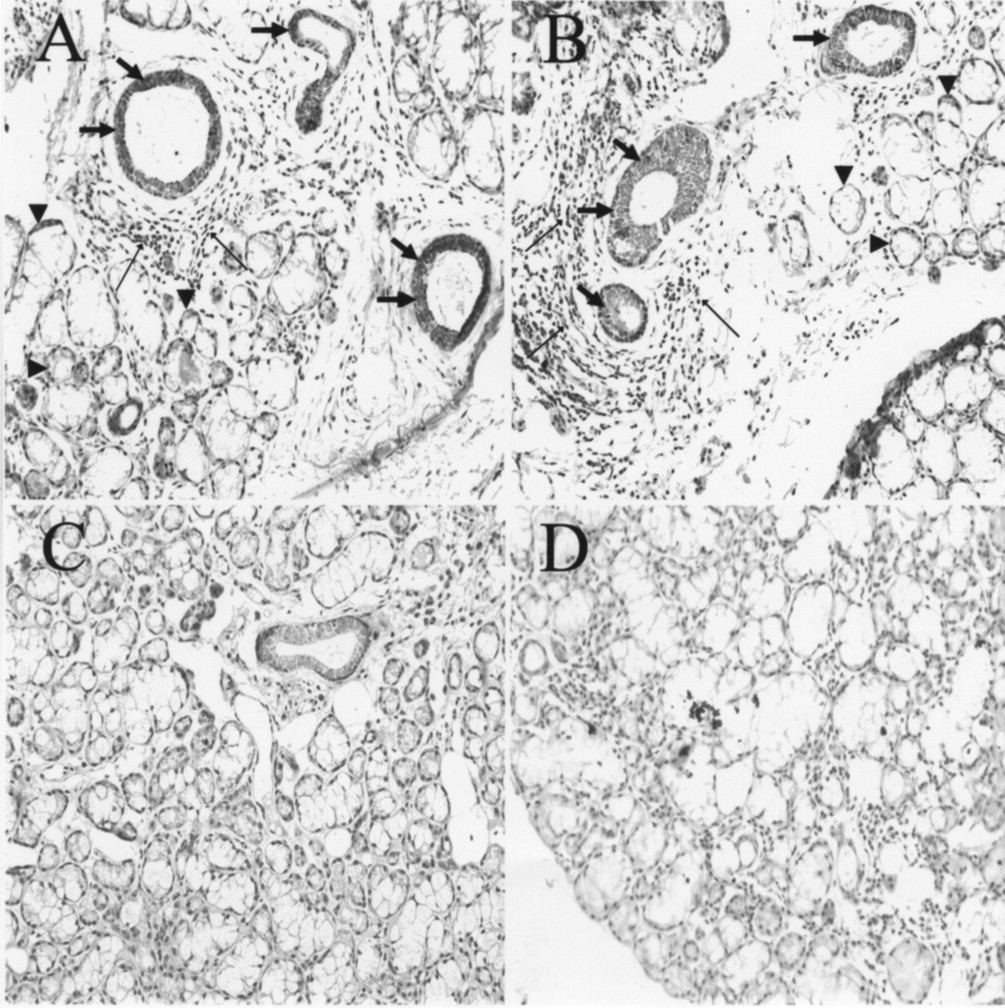
Immunohistochemical detection of p38MAPK and NF-κB in salivary gland tissues
As shown in Figure 5, immunohistochemical study demonstrated the enhancement of both pp38MAPK and NF-κB p65 expression in SS ductal cells and acinar cells located near infiltrated lymphocytes, where destruction of the acinar structure seems to occur. However, SS acinar cells distant from infiltrated lymphocytes demonstrated weak staining for both NF-κB p65 and pp38MAPK as compared with those adjacent to infiltrated lymphocytes and those in normal salivary glands.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Expression of nuclear factor-κB (NF-κB) p65 and p38 phosphorylated mitogen-activated protein kinase (pp38MAPK) in situ in minor salivary glands from patients with Sjögren’s syndrome (SS) and normal subjects. Sections of minor salivary glands were stained with specific antibodies to pp38MAPK (A and C) and NF-κB (B and D). Both ductal epithelium (large arrows) and acinar cells (arrowheads) adjacent to lymphoid infiltrates (small arrows) stained predominantly for pp38MAPK (A) and NF-κB (B) in SS salivary glands. In normal salivary glands, pp38 and NF-κB proteins were not detected (C and D; original magnification ×100). Results are representative of 6 independent experiments derived from 6 different patients with SS and 6 different controls.
DISCUSSION
Although previous reports indicated proapoptotic roles of CD40 signaling27,28, the nature of the intracellular signaling pathways and transcription factors activated by CD40 have not been studied extensively in human salivary epithelial cells. We have demonstrated for the first time that the mechanism underlying CD40-mediated salivary epithelial cell death may involve activation of p38MAPK and NF-κB signals, and upregulation of Fas expression, thereby rendering the cells more susceptible to Fas-induced apoptosis. Our study demonstrated that ductal epithelium and acinar cells adjacent to the lymphoid infiltrates of the SS salivary gland exhibited enhanced p38MAPK and NF-κB activity compared with cells distant from the infiltrated lymphocytes or with cells in normal salivary glands. Thus, these results may indicate that infiltrated lymphocytes affect the activity of NF-κB and p38MAPK in SS acinar and duct cells.
Engagement of TNF receptors, such as TNF-R2 and CD95/Fas, stimulates p38MAPK activities11,14 and CD40 is structurally homologous to the TNF-R family29. Therefore, we examined whether CD40-mediated signal transduction induces p38MAPK activation in salivary epithelial cells. Our results showed that p38MAPK phosphorylation occurred after 4 h of CD40 ligation in a time-dependent manner, and pretreatment with antagonistic anti-CD40 mAb (ch5D12) blocked p38MAPK phosphorylation (Figure 1A, 1B), suggesting that both activation of p38MAPK by CD40 and inhibition of p38MAPK by SB203580 are specific. SB203580, as a highly specific and cell-permeable inhibitor of p38MAPK, enabled us to study the role of the p38MAPK pathway in CD40-mediated salivary epithelial cell death. Pretreatment with P38MAPK inhibitor SB203580 partly blocked Fas and CD40-induced apoptosis (Figure 4A–4E). These findings suggest that the p38MAPK pathway may positively regulate apoptotic responses in a stimulus-dependent manner in salivary epithelial cells.
Many studies have shown that RelA, the p65 subunit of NF-κB, plays a key role in protecting cells from proapoptotic stimuli18,19. Conversely, a proapoptotic aspect of RelA activity has also been reported20,21,22,23,24. Thus, NF-κB can regulate both pro- and antiapoptotic signaling pathways. It is possible that this difference may be due to the different cell lines and the amounts of inducing agents used in the experiments. Our in vitro studies have shown that CD40 ligation rapidly and strongly activates NF-κB, as evidenced by prolonged NF-κB DNA-binding activity up to 24 h (Figure 2A–2C), suggesting that NF-κB is functional downstream of the CD40 signal in salivary epithelial cells. We show, for the first time, that inhibition of NF-κB activity partly abrogated Fas and CD40-mediated apoptosis (Figure 4A–4E), suggesting a possible mechanism by which NF-κB functions as a proapoptotic factor to regulate proapoptotic signaling cascades by upregulating Fas expression in salivary epithelial cells.
Our data demonstrate that CD40 ligation-induced expression of p38MAPK, pp38MAPK, NF-κB, and Fas are similar in SS salivary epithelial cells compared to that in normal cells. We also found no significant differences in the induction of apoptosis between SS cells and normal cells. One of the pathological characteristics of SS is multiple focal T lymphocyte aggregation in salivary glands1,2,3,4. Thus, these T lymphocytes constitute an important cellular source of CD40L30, which upregulates the expression levels of p38MAPK, pp38MAPK, NF-κB, and Fas in salivary epithelial cells, resulting in induction of apoptosis of these cells in salivary glands from patients with SS. In normal salivary glands, however, there are very few infiltrating T cells expressing CD40L, therefore, these T cells fail to induce expression of proapoptotic transcription factors, such as p38MAPK and NF-κB, and initiation of apoptotic signaling. In line with this view, we found that normal salivary epithelial cells can also be activated to express proapoptotic factors and undergo apoptosis when these cells were incubated with IFN-γ and anti-CD40 mAb to mimic the SS condition. Although the exact mechanism of CD40 ligation in vivo is not known, the results of our in vitro experiments with anti-CD40 mAb suggest that simultaneous activation of p38MAPK and NF-κB in response to CD40 ligation is essential for the induction of salivary epithelial cell death. In addition to our results, TNF and other proinflammatory cytokines such as IL-1ß, IL-6, and B cell-activating factor (BAFF) are also involved in the apoptosis of salivary epithelial cells31,32,33, and we will continue to investigate this. However, the fact that the degree of local destruction within exocrine glands does not always correlate with secretory dysfunction9,34 suggests the secretory function and apoptosis are both likely to be complex multifactorial processes of which CD40/CD40L and p38MAPK are among multiple agents.
Cooperation of p38MAPK and NF-κB signaling plays a critical role in salivary epithelial cell death. p38MAPK and NF-κB function as proapoptotic factors to upregulate Fas pathway and initiate cell apoptotic signaling, and this process can be controlled by CD40.
Acknowledgment
We thank Tanox Pharma B.V., Amsterdam, The Netherlands, for providing the chimeric antagonistic monoclonal antibody to human CD40 (ch5D12).
Footnotes
-
Supported by grant No. 30671948 (to Dr. Ping) from the National Natural Science Foundation of China.
- Accepted for publication February 15, 2012.
REFERENCES
- 1.
- 2.
- 3.
- 4.
- 5.
- 6.
- 7.
- 8.
- 9.
- 10.
- 11.
- 12.
- 13.
- 14.
- 15.
- 16.
- 17.
- 18.
- 19.
- 20.
- 21.
- 22.
- 23.
- 24.
- 25.
- 26.
- 27.
- 28.
- 29.
- 30.
- 31.
- 32.
- 33.
- 34.