

Abstract
Objective. To examine whether IgG glycosylation changes and MBL2 genotypes are associated with systemic inflammation and joint destruction in rheumatoid arthritis (RA).
Methods. IgG N-glycan content was determined from serum in 118 patients with RA by high-throughput glycan analysis using normal-phase high-pressure liquid chromatography. MBL2 extended genotypes (YA/YA, YA/XA, XA/XA, YA/YO, XA/YO, YO/YO) were determined. Systemic inflammation was assessed by serum levels of C-reactive protein (CRP), interleukin 6 (IL-6), and tumor necrosis factor-α (TNF-α). Joint destruction was assessed by total Sharp score (TSS) and alloplastic surgery records.
Results. IgG hypogalactosylation was significantly correlated to IL-6 (Spearman’s rho = 0.32, p < 0.001), CRP (Spearman’s rho = 0.31, p < 0.001), TSS (Spearman’s rho = 0.25, p = 0.01), and alloplastic replacement of joints (Spearman’s rho = 0.18, p = 0.05). In multivariate analysis including age, CRP, anticitrullinated protein antibodies (ACPA), and other confounders, IgG hypogalactosylation was significantly associated with TSS (p = 0.014) and alloplastic joint replacement (OR 76.5, p = 0.041) in patients homozygous for the high expression MBL2 genotype YA/YA, but not in carriers of lower expression genotypes.
Conclusion. Decreased galactosylation of IgG correlated to markers of inflammation, i.e., IL-6 and CRP. Only in patients homozygous for high expression of the MBL2 genotype YA/YA was IgG hypogalactosylation associated with markers of joint destruction. Our results suggest that inflammation-associated decreased galactosylation of IgG combined with high expression MBL2 genotypes are involved in the pathophysiology of RA.
Rheumatoid arthritis (RA) is a chronic inflammatory disease of unknown etiology affecting about 1% of the adult population. RA is characterized by synovial inflammation and joint destruction leading to impaired joint function. The course of RA is variable and different markers of prognostic value have been discussed. Acute-phase reactants, especially C-reactive protein (CRP), have been shown to be highly reliable markers of disease activity and in the long term are associated with radiographic outcomes1,2 and high titers of rheumatoid factor (RF), and antibodies against citrullinated protein (ACPA) are associated with progression of radiographic signs of joint destruction3,4,5.
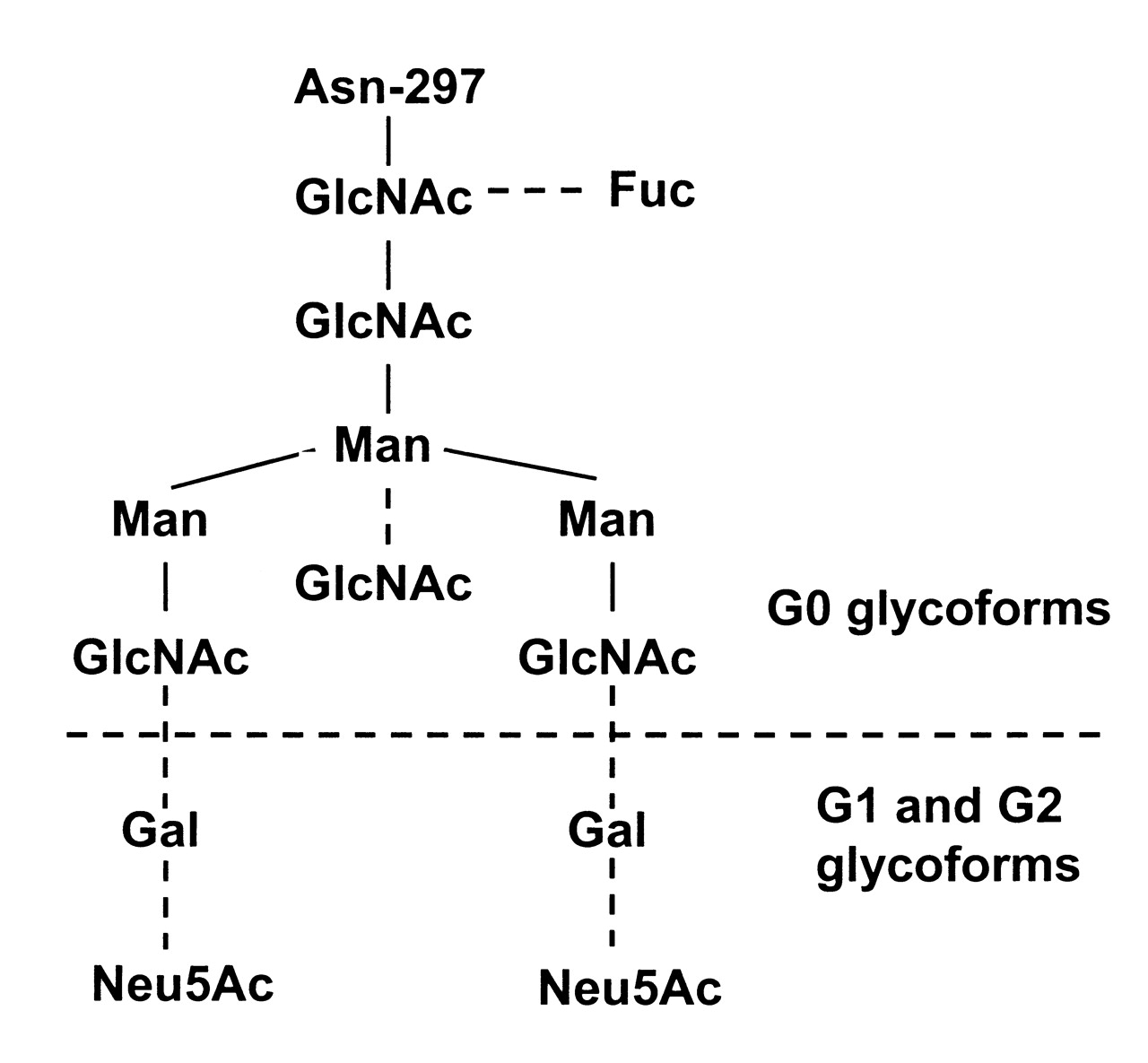
Human IgG is a glycoprotein with 2 conserved N-linked glycosylation sites. One of the glycans is present at Asn-297 on each heavy chain in the CH2 domain (Fc portion). The other N-glycan is attached to the hypervariable region of the Fab arm in about 20% of circulating IgG, and is heavily sialylated. The N-glycans at Asn-297 are of complex biantennary type with a heptasaccharide core that terminates in 2 N-acetylglucosamine (GlcNac) residues onto which may be added core fucose, bisecting GlcNAc, terminating galactose, and sialic acid, giving rise to the presence of multiple glycoforms (Figure 1). Based on the presence or absence of terminal galactose on Asn-297 glycans, IgG have been divided into 3 categories: nongalactosylated (G0), monogalactosylated (G1), and digalactosylated (G2) IgG. In healthy individuals the IgG G0 glycoform accounts for about 20%–35% of the total IgG glycan pool, but in patients with RA serum levels of IgG G0 may rise 3-fold6. Serum levels of hypogalactosylated IgG also increase with age7,8. Studies indicate that decreased galactosylation of IgG may be of diagnostic and prognostic value in RA9,10,11,12,13.

Structure of the complex biantennary oligosaccharide at Asn-297. Solid lines indicate core heptasaccharide; broken lines: additional sugars that may be added. Fuc: fucose; GlcNAc: N-acetylglucosamine; Gal: galactose; Man: mannose; Neu5Ac: sialic acid.
Mannose-binding lectin (MBL) is a liver-derived component of the innate immune system that may bind to various sugar motifs on microorganisms and apoptotic cells and thereby activate complement through MBL-associated serine proteases. Serum concentrations of MBL mainly reflect the expression of the MBL2 gene in the liver. Subjects homozygous or compound-heterozygous for defective structural MBL2 alleles produce dysfunctional low molecular weight MBL with little or no apparent biological function, whereas subjects with the extended YA/YA genotype have the highest MBL-producing potential14.
MBL may play differentiated roles in the pathogenesis of RA. In some studies MBL deficiency has been associated with erosive disease15,16. Other studies have been inconclusive in this regard17,18. Conversely, the high-expressing MBL2 YA/YA genotype has a synovitis-enhancing effect in ACPA-positive patients with early RA19 and high serum levels of MBL are associated with increased risk of developing RA20 and with increased overall and cardiovascular mortality in RA21.
In vitro MBL interacts with the IgG G0 glycoform through the exposed GlcNac and thereby activates complement22. In one study we observed that high serum levels of the IgG G0 glycoform in combination with genetically determined high serum levels of MBL were associated with development of ischemic heart disease in patients with RA23. Moreover, a previous study suggested that MBL/IgG G0 complexes may increase inflammation in late-onset and rapidly progressive RA24.
We used a recently developed high-throughput method for IgG glycan quantification25 in this study. We examined if IgG hypogalactosylation and MBL2 genotypes were associated with markers of systemic inflammation and joint destruction in patients with RA. We also investigated possible interaction between hypogalactosylated IgG and the high-expressing MBL2 YA/YA genotype with respect to joint damage.
MATERIALS AND METHODS
Patients
One hundred eighteen consecutive unrelated white Danish patients with RA attending the Department of Rheumatology, Rigshospitalet, Copenhagen University Hospital, were included from October 2006 to March 2007 as described26. The median disease duration at inclusion was 12 years (range 1–41 yrs). All patients fulfilled the American College of Rheumatology 1987 classification criteria for RA27. At inclusion, all patients underwent a detailed interview and a clinical examination. Posteroanterior (PA) and PA oblique radiographs of hands and feet were taken. Serum samples and whole EDTA blood for DNA extraction were collected and stored at −80°C.
Our study was approved by the local scientific ethical committee (reference no. KF 01286984). Written informed consent was obtained from each patient.
Clinical and laboratory assessment
Demographics, disease descriptors, and comorbidities recorded were as follows: (1) sex and age at inclusion; (2) numbers of tender and swollen joints; (3) presence of serum ACPA and RF; (4) current treatment with glucocorticoids, methotrexate (MTX), and tumor necrosis factor-α (anti-TNF-α) drugs; (5) history of ischemic heart disease; (6) history of diabetes; and (7) smoking status calculated as “pack-years” [number of packs (20 cigarettes) per day times number of years]. The following outcome variables were assessed: (1) systemic inflammation measured by serum levels of interleukin 6 (IL-6), CRP, and TNF-α; and (2) joint destruction, evaluated according to the principles of the Sharp/van der Heijde scoring method [total Sharp score (TSS), possible range 0–448]28 and alloplastic surgery records. All patients were interviewed and examined by the same physician, and the same radiologist evaluated all radiographs of hands and feet without knowledge of other study data. In 10 cases it was not possible to calculate TSS because of previous articular surgery and no available radiographs to which the last-observation-carry-forward method could be applied. Five of these belonged to the group of patients with alloplastic surgery (n = 27) and 5 belonged to the group without alloplastic surgery (n = 91). All the alloplastic surgeries were performed because of joint destruction caused by chronic inflammation, e.g., destruction of joint surface and deformity. Clinical charts were used to verify information regarding alloplastic surgery and treatment with antirheumatic drugs.
Quantification of IgG G0 glycoforms
IgG N-glycan analysis was performed using a 96-well plate in-gel block high-throughput glycan analysis method as described25. N-glycans were released from the serum samples by N-glycosidase F (PNGase F; Prozyme, San Leandro, CA, USA) and fluorescent labeled with 2-aminobenzamide (2AB; LudgerTag 2AB labeling kit; Ludger, Abingdon, UK) and analyzed by normal phase high-pressure liquid chromatography (NP-HPLC) with a 60-min run time. Peak retention times were expressed as glucose unit values relative to a 2AB–dextran ladder standard (glucose homopolymer) using Empower GPC software (Waters Corp., Milford, MA, USA). Structural assignments of the NP-HPLC glycan profile data were performed by database matching using Glycobase29. The content of IgG G0 glycoforms in serum was expressed as the ratio of core fucosylated nongalactosylated (FG0) to core fucosylated monogalactosylated (FG1).
MBL2 genotypes
Detection of MBL2 genotypes was as described30. All patients were genotyped for 3 MBL2 structural polymorphisms and for a promoter polymorphism in position −221 (X/X, X/Y, Y/Y). Patients were stratified into those with the extended YA/YA genotype (high-producing) and those with low and intermediate MBL-producing genotypes (non-YA/YA genotypes: YA/XA, XA/XA, YA/YO, XA/YO, and YO/YO).
Statistical analysis
Nonparametric analyses (Spearman’s rank correlation, Mann-Whitney test) were initially used to examine the relation between FG0/FG1 ratio, MBL2 genotypes, and markers of inflammation and joint destruction. Logarithmic transformation of FG0/FG1 ratio allowed the relations between FG0/FG1 ratio, CRP, IL-6, and TSS to be further determined by simple and multivariate linear regression analyses. The relations between FG0/FG1 ratio and alloplastic surgery were analyzed by univariate and multivariate logistic regression analyses. Adjustment variables in the multivariate analyses of CRP and IL-6 were sex, age, ACPA, and treatment with antirheumatic drugs; in the multivariate analysis of TSS and alloplastic surgery, CRP was included in addition to the variable mentioned above. All regression analyses were repeated among patients with the extended YA/YA genotype and patients with the non-YA/YA genotypes. Equality of regression coefficients was tested by calculating the Z score. Statistical processing was performed with SPSS 15. Statistical significance was defined as p value < 0.05.
RESULTS
Clinical and laboratory variables at inclusion for all patients, patients with the extended YA/YA genotype (n = 43; 36%), and patients with the non-YA/YA genotype (n = 75; 64%) are shown in Table 1. The mean FG0/FG1 ratio was 1.18 ± 0.41 and the median FG0/FG1 ratio was 1.12 (range 0.6–3.3). The mean FG0/FG1 ratio was 1.21 ± 0.47 among patients with the YA/YA genotype and 1.17 ± 0.38 among patients with the non-YA/YA genotype. Using Levene’s test for equality of variance we found no difference in the FG0/FG1 ratio between patients with the YA/YA genotype and those with the non-YA/YA genotype (p = 0.57). We found no significant correlations between the FG0/FG1 ratio and disease descriptors, i.e., tender and swollen joint counts, RF, ACPA, and treatment with antirheumatic drugs.
Clinical and laboratory variables assessed in the cohort of 118 patients with rheumatoid arthritis (RA).
Markers of systemic inflammation
Results of Spearman’s rank correlation analyses between FG0/FG1 ratio and markers of systemic inflammation are shown in Table 2. FG0/FG1 ratio was significantly correlated to IL-6 (correlation coefficient 0.32, p < 0.001) and CRP (correlation coefficient 0.31, p = 0.001). There were no significant correlations between FG0/FG1 ratio and TNF-α (correlation coefficient −0.09, p = 0.32). Using the Mann-Whitney test we found no differences in IL-6, CRP, or TNF-α between patients with the extended YA/YA genotype and patients with the non-YA/YA genotypes.
Results of Spearman rank correlation analysis between FG0/FG1 ratio and outcome variables.
Table 3 shows that the FG0/FG1 ratio was significantly associated with IL-6 and CRP in both simple and multivariate linear regression analyses including all patients. However, these associations did not differ between patients with the extended YA/YA genotype and patients with the non-YA/YA genotypes.
Results of linear regression analyses examining the association between FG0/FG1 ratio* and C-reactive protein (CRP), interleukin 6 (IL-6), and total Sharp score (TSS).
Joint destruction
Using Spearman’s correlation analysis we found that the FG0/FG1 ratio was significantly correlated to TSS (correlation coefficient 0.25, p = 0.01) and to alloplastic replacement of joints (correlation coefficient 0.18, p = 0.05; Table 2). Using the Mann-Whitney test we found no significant difference in TSS or alloplastic replacement of joints between patients with the extended YA/YA genotype and patients with the non-YA/YA genotypes.
Simple regression analysis showed a significant linear association between FG0/FG1 ratio and TSS for all patients (regression coefficient 110, p = 0.041). However, upon dividing the patients into 2 groups based on the MBL2 genotypes we found that this linear association was significant in patients with the extended YA/YA genotype (regression coefficient 243, p = 0.003) and not in patients with the non-YA/YA genotypes (regression coefficient 20.7, p = 0.772; Table 3). In multivariate linear regression analysis allowing adjustment for confounders we found that the linear association between FG0/FG1 ratio and TSS remained significant in patients with the extended YA/YA genotype (regression coefficient 272, p = 0.014; Table 3).
Table 4 shows that the FG0/FG1 ratio was significantly associated with alloplastic replacement of joints in univariate logistic regression analysis including all patients and patients with the extended YA/YA genotype (OR 3.27, p = 0.022; OR 13.9, p = 0.017) but not in patients with the non-YA/YA genotypes (OR 1.49, p = 0.57). In the multivariate analysis FG0/FG1 ratio only remained significantly associated with alloplastic replacement of joints in patients with the extended YA/YA genotype (OR 76.5, p = 0.041).
Results of logistic regression analysis examining the association between FG0/FG1 ratio and alloplastic surgery.
Further, we found that the FG0/FG1 ratio association with TSS and alloplastic replacement of joints differed significantly between patients with the extended YA/YA genotype and patients with the non-YA/YA genotypes, being highest in the first group (Tables 3 and 4).
DISCUSSION
In our study of 118 patients with RA we used a validated high-throughput method for IgG glycan quantification9. Our results showed that the FG0/FG1 ratio was associated with IL-6 and CRP. Moreover, in patients homozygous for the high-expression MBL2 YA/YA genotype, the FG0/FG1 ratio was significantly associated with markers of joint destruction independent of age, CRP, ACPA status, and other potential confounders.
In a recent study by Ercan, et al9 the mean FG0/FG1 ratio was 1.36 ± 0.43. In our study the mean FG0/FG1 ratio was 1.18 ± 0.41, which is relatively low given the age of the patients in our cohort. However, the low FG0/FG1 ratio in our study could be explained by the fact that our RA population generally had low disease activity, with a mean Disease Activity Score 28-CRP (DAS28-CRP) of 3.09 ± 1.35 (mean DAS28-CRP in the study by Ercan, et al9 was 4.15 ± 1.74) due to effective medication of most patients with MTX and/or TNF-α inhibitors at the time of inclusion. The low disease activity could also explain the lack of significant relations between hypogalactosylated IgG and number of tender and swollen joints in our study compared to the study by Ercan, et al9. Despite these differences both studies showed highly significant associations between the FG0/FG1 ratio and markers of systemic inflammation, suggesting that IgG hypogalactosylation may reflect the inflammatory process in RA. As the overall regulation of IgG glycosylation in plasma cells is not fully understood, future investigations may include studies on how various cytokines and immune factors influence the glycosyltransferases activities that determine the composition of the arms of the N-glycans.
We found that the FG0/FG1 ratio was significantly associated with radiographic scores of joint destruction and alloplastic joint replacement in patients homozygous for the high-expression MBL2 YA/YA genotype but not in carriers of lower-expression genotypes. These results were consistent in both univariate and multivariate analysis with adjustment for sex, age, CRP, ACPA status, and immunomodulatory treatment. Further, the differences observed in the associations were statistically significant, lending validity to the concept that the coexistence of hypogalactosylated IgG and MBL may have a negative influence on joint integrity in RA.
In a previous study24, a weak correlation was observed between erosion score and hypogalactosylated IgG in patients with RA who are homozygous for the normal MBL2 genotype. Moreover, we have recently shown that high serum levels of MBL in combination with high serum levels of IgG G0 glycoforms confer a significant risk of developing ischemic heart disease including acute myocardial infarction in patients with RA23. Our observation that hypogalactosylated IgG associates only with markers of joint damage in patients with the high-expressing MBL2 YA/YA genotype parallels these previous findings, and is in accord with the hypothesis that MBL and IgG G0 complexes are harmful in RA22. However, it remains to be proven experimentally that complexes of IgG G0 and MBL participate in the development of destructive joint pathology in RA. Thus we cannot exclude that our observation reflects coexisting, but independent pathophysiological change events indirectly associated with the clinical events studied.
Glycosylation changes could also be linked to inflammation and to the pathogenesis of RA in other ways. A previous study showed that absence of sialic acid on IgG results in proinflammatory responses through engagement of specific Fc receptors on effector cells31,32. As IgG-Fc degalactosylation inevitably leads to decreased amounts of sialic acid attached to IgG-Fc it could be suggested that desialylation also plays an important role in the inflammatory processes in RA.
Fucose on IgG heavy chains is increased in patients with RA compared to healthy controls33 and increased IgG fucosylation was found to be a diagnostic marker in juvenile chronic arthritis34. As fucose may alter antibody-dependent cellular cytotoxicity by affecting the binding of IgG to FcγRIIIa35, fucosylation changes on IgG heavy chains may also play a role in the pathogenesis of RA. In this respect the FG0/FG1 ratio, which is based on fucosylated G0 and fucosylated G1, may be a more relevant measurement compared to the classical %G0, which includes fucosylated as well as nonfucosylated G0, G1, and G2 Fc-glycans.
We did not find a significant difference between the high-expressing MBL2 YA/YA genotype and the non-YA/YA genotypes with respect to markers of disease activity or joint destruction, which is in line with findings in 2 studies17,18. However, the results concerning the role of MBL in RA are divergent and it is likely that differences in study design, differences in medication, age and duration of disease, and confounders such as IgG G0 may have influenced the observed associations. Several pathophysiological mechanisms have been discussed in relation to the role of MBL in RA. It has been hypothesized that MBL deficiency may result in defective clearance of pathogens and circulating immune complexes as well as dying host cells and cellular debris, thereby aggravating ongoing joint inflammation leading to joint damage in RA. In contrast, the high-expressing MBL2 genotypes could be associated with excess joint inflammation and increased joint destruction.
We show that decreased IgG galactosylation is associated with systemic inflammation in RA and that the presence of decreased IgG galactosylation and high-expression MBL2 genotypes may contribute to the pathogenesis of RA. These findings need to be corroborated by experimental studies examining the molecular details of the putative MBL/IgG G0 interaction in vivo. As IgG hypogalactosylation is also increased in other inflammatory diseases36,37,38, future studies may address whether our findings are applicable beyond RA.
Acknowledgment
Matthew Campbell from Dublin-Oxford Glycobiology Laboratory is acknowledged for his critical reading of the manuscript.
Footnotes
-
Supported by grants from the Danish Rheumatism Association, The Novo Nordisk Research Foundation, The Capital Region of Denmark, and Rigshospitalet, Copenhagen University Hospital.
- Accepted for publication November 18, 2011.








{kind=link}