

Abstract
Objective. Membrane-bound receptor for advanced glycation endproducts (mRAGE) is overexpressed in response to increasing concentrations of its ligand (e.g., S100A12) and triggers an inflammatory immune response. In contrast, soluble RAGE (sRAGE) acts as a decoy receptor and downmodulates inflammation. Decreased sRAGE levels are associated with autoimmune diseases; however, limited data are available in juvenile idiopathic arthritis (JIA). We studied sRAGE levels in patients with JIA [enthesitis-related arthritis (ERA) category].
Methods. sRAGE levels were estimated in the serum of patients with ERA JIA (n = 101), systemic-onset JIA and polyarticular JIA (n = 10 each), and healthy controls (n = 45). Synovial fluid (SF) sRAGE was measured in patients with ERA, rheumatoid arthritis, reactive arthritis, and osteoarthritis (n = 10). Levels of S100A12 were also measured. Twenty-four patients with ERA were followed for 4 months. Disease activity was assessed by swollen joint count (SJC), tender joint count (TJC), and erythrocyte sedimentation rate (ESR). All levels are expressed as median (range).
Results. The serum sRAGE (pg/ml) level was significantly lower in patients compared to healthy controls [515 (64–1887) vs 1542 (627–3159); p < 0.0001]. In paired samples, SF had lower levels compared to corresponding plasma level [102 (51–799) vs 481 (134–1006); p < 0.0001]. The level of S100A12 (ng/ml) was higher in SF (1042; 573–1415) than serum (638; 208–779). Serum sRAGE correlated negatively with S100A12 levels (r = −0.474; p < 0.01.), ESR (r = −0.306; p < 0.01), and SJC (r = −0.237; p < 0.05), but not with TJC (r = −0.134; p = NS). The levels of sRAGE remained stable over time in patients with stable disease.
Conclusion. Levels of sRAGE are reduced in patients with ERA and correlate negatively with disease activity and S100A12 levels. sRAGE may be a modulator of inflammation in these patients.
The receptor for advanced glycation endproducts (RAGE) is a cell-surface molecule belonging to the immunoglobulin super-family and is expressed by several cells, including immune cells such as neutrophils, macrophages, and T cells1. It is encoded in the Class III region of the major histocompatibility complex. It recognizes a wide repertory of ligands2 — the advanced glycation endproducts, the S100/calgranulin family of proinflammatory cytokine-like proteins, and the high mobility group box protein-1. Receptor-ligand binding results in transcription of proinflammatory factors and in overexpression of the receptor on the cell surface. Increasing evidence suggests that membrane-bound RAGE (mRAGE) acts like a non-canonical pattern recognition receptor and its dysregulation leads to persistent inflammation in diseases such as diabetes, atherosclerosis, arthritis, and Alzheimer’s disease3.
Soluble RAGE (sRAGE) lacks the short cytoplasmic domain that not only anchors the full receptor to a cell, but also transmits downstream signals. sRAGE functions as a “decoy receptor,” competing with its membrane-bound form for the same ligand and causing a reduction in overall signaling4. sRAGE may also act as a direct inhibitor of leukocyte recruitment5. Thus, it is capable of reducing the inflammation caused by mRAGE. sRAGE interacts with the integrin Mac-1 and triggers a proinflammatory cascade6, suggesting a dual function for sRAGE.
Calgranulin C, also called S100A12 or EN-RAGE (extra-cellular newly identified RAGE-binding protein), is a member of a multigene family involved in calcium-dependent regulation of several cellular activities such as protein phosphorylation, cellular homeostasis, and even inflammation7. It is secreted mainly by neutrophils and activated macrophages8 and is now recognized as a damage-associated molecular pattern molecule (DAMP), a group that represents endogenous ligands of pattern recognition receptors9. It binds to mRAGE and induces inflammatory cascade10.
There are several studies in which dysregulation in levels of sRAGE and S100A12 have been associated with inflammatory diseases. Studies done in different arthritides found serum levels of sRAGE to be lower in rheumatoid arthritis (RA)11, osteoarthritis (OA)12, and Sjögren’s syndrome13. sRAGE levels also correlated negatively with S100A12 levels in Kawasaki disease14. In addition, serum sRAGE levels in patients with RA were found to be negatively associated with current smoking, family history of cardiovascular disease, history of vasculitis, blood pressure, presence of rheumatoid factor, and serum levels of C-reactive protein (CRP) and S100A12, again suggesting that sRAGE acts as an antiinflammatory molecule and S100A12 as a proinflammatory one15.
Juvenile idiopathic arthritis (JIA) is another disease characterized by inflammation and articular damage. Among all the categories of JIA16, enthesitis-related arthritis (ERA) is most prevalent in India. Studies have shown elevated levels of S100A12 in serum of patients with JIA8,14,17. Data on sRAGE levels in JIA are limited. Only a single study on Kawasaki disease had used patients with JIA as disease controls and shown that sRAGE levels were lower in the serum of children with systemic-onset JIA and were similar to healthy controls in oligoarticular and polyarticular JIA (polyJIA)14. That study had not included children with ERA. In our study we measured sRAGE and S100A12 levels in serum, synovial fluid (SF), and paired plasma of patients with ERA and studied its correlation with disease activity. Further, we studied the proportion of these patients longitudinally to see whether sRAGE levels change over time and whether they correlate with changes in disease activity.
MATERIALS AND METHODS
Patients and controls
Patients with ERA who satisfied International League of Associations for Rheumatology criteria16 and who gave or whose parents gave written informed consent were enrolled in the study. Nonrelated healthy controls matched for age and sex were enrolled from blood bank donors. Stored samples of patients with systemic-onset (SoJIA) and polyJIA were used as disease controls. The study was approved by the institutional ethics committee. We also measured sRAGE levels in stored SF of patients with RA, OA, and reactive arthritis (ReA) as disease controls.
Separation of serum, plasma, and SF
Blood samples and SF were centrifuged at 1000 g for 10 min within 30 min of withdrawal. Samples for plasma were collected in lithium heparin. The serum/plasma/cell-free SF thus obtained was stored at −80°C.
ELISA
sRAGE (DuoSet ELISA development kit; R&D Systems, Minneapolis, MN, USA) and S100A12 (Abnova Corp., Taipei, Taiwan) levels were measured in serum/plasma/cell-free SF and determined using sandwich ELISA as per manufacturer’s instructions.
Statistical analysis
Data were analyzed using GraphPad Prism (GraphPad Software Inc., La Jolla, CA, USA) and SPSS 13.0 software (SPSS Inc., Rainbow Technologies, Somers, NY, USA). Because we found no difference between serum and plasma levels in ERA, we analyzed both these samples together and henceforth they have been referred to as serum. The Mann-Whitney U test was used for comparing differences in serum analyte concentrations of patients and controls, while analysis of paired plasma and SF samples was done by Wilcoxon signed-rank test. The Kruskal-Wallis test was used for multiple group comparison. The relationship between sRAGE concentration and disease activity was measured by Spearman’s rank correlation. All results are expressed as median (range).
RESULTS
The study involved 101 patients with ERA (median age at sample collection 15 years and median disease duration 3 years; 98 males, 84 HLA-B27-positive; Table 1), 10 patients with SoJIA (median age at sample collection 13 years, 8 males, median disease duration 4.8 years, 9 patients taking methotrexate and 8 taking steroids; Table 1), 10 patients with polyJIA (median age at sample collection 13.5 years, 6 males, median disease duration 4 years, 9 patients taking methotrexate and 6 taking steroids; Table 1), and 45 healthy controls (median age 24 years; all males).
Clinical details of patients with juvenile idiopathic arthritis (JIA).
Median sRAGE levels were significantly lower (p < 0.0001) in patients with ERA (515; 64–1887 pg/ml) than in healthy controls (1542; 627–3159 pg/ml). There was no difference in levels between patients with ERA, patients with SoJIA (300; 165–400 pg/ml), and patients with polyJIA (380; 180–1000 pg/ml; Figure 1A).
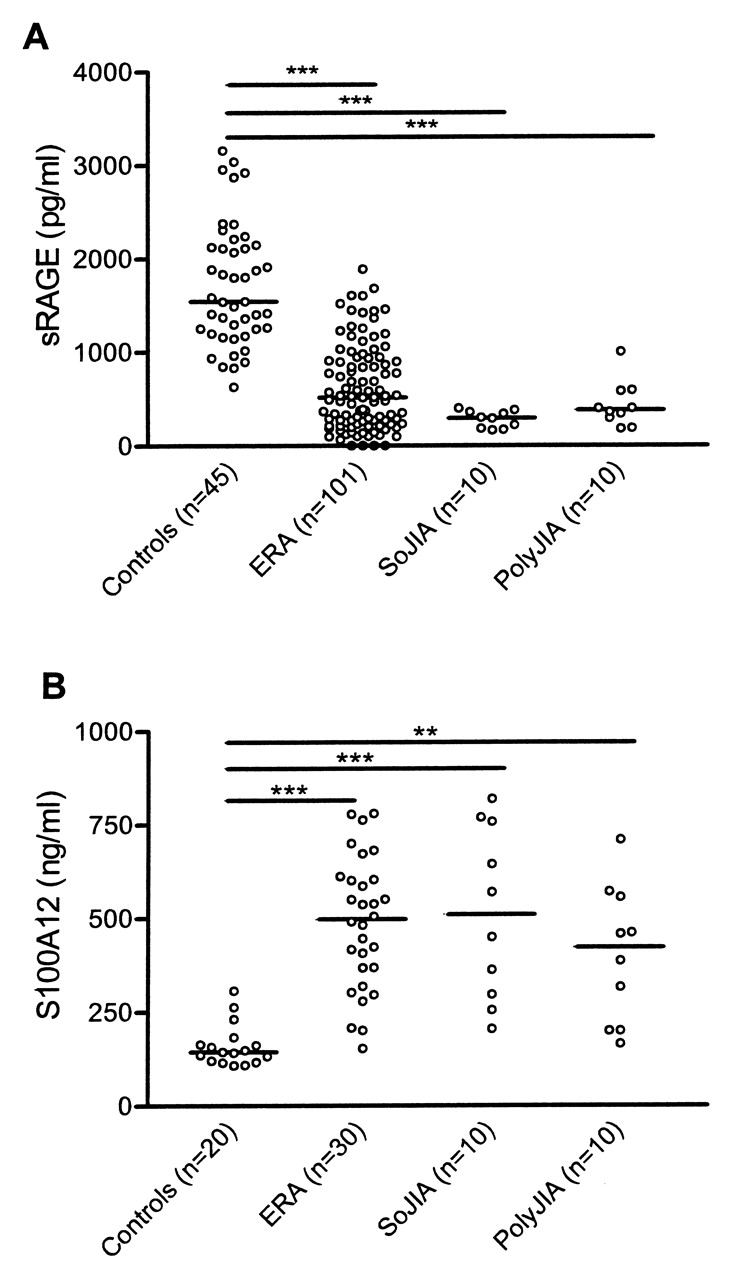
Levels of soluble receptor for advanced glycation endproducts (sRAGE; A) and S100A12 (B) in serum of healthy controls and patients with enthesitis-related arthritis (ERA), systemic-onset juvenile idiopathic arthritis (SoJIA), and polyarticular JIA (polyJIA). ***p < 0.0001; **p < 0.01. No difference was seen between sRAGE and S100A12 levels in ERA, SoJIA, and polyJIA sera.
Median S100A12 levels were significantly higher (p < 0.01) in patients with ERA (498; 153–783 pg/ml) than in healthy controls (144; 108–307 pg/ml). There was no difference in levels between patients with ERA, patients with SoJIA (510; 205–819.6 pg/ml), and patients with polyJIA (423; 165–710 pg/ml; Figure 1B).
SF and paired plasma were available from 10 patients with ERA (median age at onset 10.5 years and median disease duration 8 years; all males). Eight were HLA-B27-positive, 1 was negative, and data were not available for 1 patient. Five patients were taking nonsteroidal antiinflammatory drugs and 3 were taking methotrexate. Median sRAGE levels (pg/ml) in SF (102.5; 51–799) were lower than those in plasma (481.6; 134–1006; p < 0.0001; Figure 2A). Median SF S100A12 levels (1042; 573–1415) were higher (p < 0.01) than in plasma (638; 208–779; Figure 2B).

Soluble receptor for advanced glycation endproducts (sRAGE) and S100A12 levels in paired synovial fluid and serum of ERA patients (n = 10). Each dot represents an individual patient. Median is marked on the plot. p value calculated by Wilcoxon signed-rank test. A. sRAGE levels. B. S100A12 levels.
Median sRAGE levels in ERA SF did not differ significantly from those in RA (180; 64–430), OA (180; 64–580), and ReA SF (170; 140–290).
The serum sRAGE levels in ERA correlated negatively with S100A12 levels (r = −0.474; p < 0.01; Figure 3). The levels also showed inverse correlation with ESR (r = −0.306; p < 0.01), swollen joint count (SJC; r = −0.237; p < 0.05), but not with TJC (r = −0.134; p = nonsignificant), which were taken as markers of disease activity (Figure 4). The sRAGE and S100A12 levels did not have any association with the age of the patients.

Correlation between levels of soluble receptor for advanced glycation endproducts (sRAGE; pg/ml) and S100A12 (ng/ml) in serum of patients with enthesitis-related arthritis (n = 30). r = −0.474.
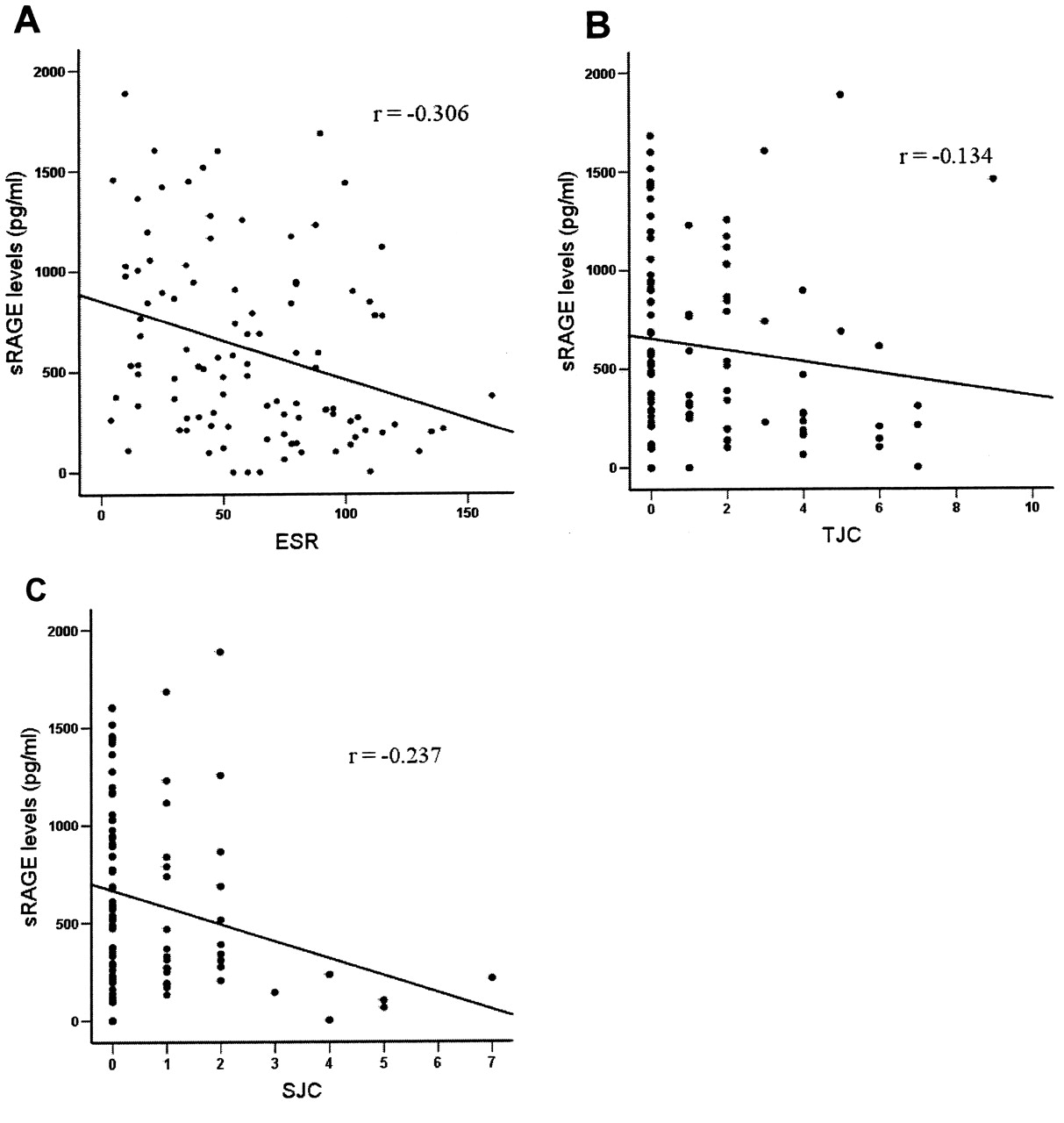
Correlation between (A) levels of soluble receptor for advanced glycation endproducts (sRAGE) and erythrocyte sedimentation rate (ESR); r = −0.306; p < 0.05. B. sRAGE levels and tender joint count; r = −0.134; p = nonsignificant. C. sRAGE levels and swollen joint count; r = −0.237; p < 0.05. Each dot represents serum from individual patient with enthesitis-related arthritis (ERA; n = 101).
In 24 paired serum samples obtained at a median difference of 4 months, there was no significant difference between SJC, TJC, ESR, and sRAGE at baseline and followup.
DISCUSSION
Our study revealed that levels of sRAGE in the serum/plasma of patients with JIA-ERA were lower and levels of S100A12 higher compared to healthy controls, but were similar to those of patients with SoJIA and polyJIA. Further, in paired samples, the SF sRAGE level was lower and S100A12 levels were higher than the corresponding plasma level. Serum sRAGE levels had inverse correlation with disease activity measures such as ESR and SJC, and also with S100A12 concentration. However, in patients with stable disease sRAGE levels did not change significantly.
Our observation of reduced sRAGE and elevated S100A12 levels in children with ERA-JIA is similar to the data reported in children with SoJIA14. We found lower levels of S100A12 in SoJIA compared to the study by Wittkowski, et al14. The likely reason could be that their study enrolled drug-naive patients at the start of illness, while most of our patients were taking disease-modifying antirheumatic drugs and steroids and had a longer disease duration. Similar reduction in sRAGE and elevation in S100A12 levels is seen in RA15 and OA12. On the other hand, while 1 study showed that patients with oligoarticular and polyJIA have sRAGE levels similar to those of healthy controls14, we found lower sRAGE levels in patients with polyJIA compared to healthy controls. S100A12 is known to stimulate production of proinflammatory cytokines upon binding to mRAGE10; it is likely that an increase in its levels contributes significantly to JIA pathology. This effect is further worsened by decrease in sRAGE concentration, which, under normal conditions, mops up mRAGE ligand, thereby modulating its signaling.
SF sRAGE levels were even lower compared to the plasma levels. This is reflective of the potent regulatory role played by sRAGE at the local site of inflammation. Our data are similar to previous reports in RA11 and OA12. Lower sRAGE levels in SF may result in continued activation of RAGE pathways, leading to inflammation. Further, S100A12 levels were markedly higher in SF than in plasma. This observation is again in tandem with findings in RA11, polyJIA, and oligoarticular JIA17. More recently, Baillet, et al18 showed that S100A12 was one of the most upregulated proteins in RA SF and could distinguish RA from other joint diseases with great accuracy.
Synovial tissue samples from both RA and OA show expression of mRAGE; however, the levels were higher in RA19. It is mainly expressed by macrophages, T cells, and B cells. Cultured human synovial fibroblasts also constitutively express RAGE and on stimulation with cognate ligands produce monocyte chemoattractant proteins-120. S100A12, after binding to RAGE, produces proinflammatory cytokines and matrix metalloprotease21. Thus, modulation of RAGE expression in the synovial compartment might be a therapeutic avenue. Indeed, Pullerits, et al22 measured and demonstrated autoantibodies against RAGE in the SF of patients with RA. The autoantibodies’ presence correlated with less erosive disease, suggesting that blocking the RAGE pathway can reduce severity of disease.
In our study, negative correlation was seen between sRAGE levels and measures of disease activity such as SJC and ESR. In RA, serum sRAGE levels correlated with CRP levels, presence of rheumatoid factor, and vasculitis, but not with age, duration of disease, or erosions15. In another study, lower urinary sRAGE level had an association with increased risk of atherothrombosis in RA23. In SoJIA and Kawasaki disease, sRAGE levels correlated negatively with CRP levels14. In our longitudinal study, we did not find any difference in sRAGE levels, suggesting that sRAGE remains stable over time, when there is no change in disease activity. Thus it can be used as a biomarker. In Kawasaki disease, sRAGE levels normalize once the inflammation subsides after intravenous immunoglobulin treatment14. In RA followup, study levels at 1 year were significantly different from those at baseline, again suggesting inflammation-mediated regulation of sRAGE levels23. Indeed, RAGE mRNA increases in adherent macrophages on exposure to proinflammatory cytokines, especially interleukin 1ß19.
The etiology and pathogenesis of JIA-ERA are unknown, but innate immunity is postulated to play a role in disease perpetuation. These triggers can be exogenous (microbial products)24 or endogenous damage-associated molecular patterns (e.g., S100A12). They stimulate immune responses through pattern recognition receptors, and the mRAGE is one of them. sRAGE probably absorbs the ligands of the mRAGE and thus downmodulates the proinflammatory response. sRAGE also affects survival of monocytes/neutrophils and macrophage development25. In other words, a decrease in sRAGE levels probably tips the balance in favor of inflammation. However, we were unable to establish a cause-and-effect relationship between decreased sRAGE and increased S100A12 levels.
Our study suggests that patients with ERA have low serum and SF sRAGE levels and that these levels correlate negatively with disease activity as well as concentration of S100A12 protein. Thus sRAGE may be involved with downmodulation of inflammation in patients with ERA.
Footnotes
-
Supported by a grant from the Department of Biotechnology, Government of India, to Dr. Aggarwal. Ms. Myles is supported by the Council of Scientific and Industrial Research-Junior Research Fellowship.
- Accepted for publication April 21, 2011.








{kind=link}
{kind=link}
{kind=link}
{kind=link}