

Abstract
Objective. Genetic factors account for more than 90% of overall susceptibility to ankylosing spondylitis (AS), and recent studies have focused on non-major histocompatibility complex genes. Vitamin D binding protein (DBP) is a highly polymorphic protein that transports vitamin D and its metabolites. In addition to its sterol binding capacity, DBP has many other roles in the inflammatory and immune systems, and has been reported to be associated with autoimmune diseases. We investigated the association between DBP polymorphisms and susceptibility to AS.
Methods. This case-control study was conducted in 223 patients with AS and 239 ethnically matched controls who were genotyped for 8 single-nucleotide polymorphisms (SNP) in the DBP and its promoter. Genomic DNA was isolated from peripheral blood leukocytes using the standard phenolchloroform method, and the GoldenGate assay was used for genotyping.
Results. No significant association was found between the susceptibility to AS and DBP polymorphisms. In a subgroup analysis of patients with AS, G alleles at rs222016 and rs222020 (OR 0.63, 95% CI 0.42–0.95, p = 0.03; OR 0.63, 95% CI 0.42–0.95, p = 0.03, respectively) and A allele at rs3733359 (OR 0.59, 95% CI 0.39–0.90, p = 0.01) showed the decreased risk of peripheral arthritis. G allele at rs4752 showed increased risk of uveitis (OR 2.04, 95% CI 1.12–3.72, p = 0.02). On the haplotype analyses, haplotype 2 (AGGA) protected against the development of peripheral arthritis (p = 0.01) and haplotype 3 (GAAG) was associated with an increased likelihood of uveitis (p = 0.02).
Conclusion. DBP gene polymorphisms are associated with the development of peripheral arthritis and uveitis in Korean patients with AS. Given the influence of different DBP variants on the immune system, larger-scale studies are warranted to elucidate the role of DBP in the pathogenesis of AS.
Ankylosing spondylitis (AS) is a chronic inflammatory disease that affects the axial spine and sacroiliac joints, and leads to new bone formation and ankylosis. AS is known to have a strong genetic factor that determines disease susceptibility and severity. Considerable research effort has been focused on HLA-B27 in AS, but this risk allele contributes only partly to genetic susceptibility to AS1,2,3. Several reports have revealed that an association exists between non-major histocompatibility complex (MHC) genes and AS susceptibility. And non-MHC genes have been estimated to account for at least half of the genetic variance in AS susceptibility4,5.
Studies of the roles of vitamin D in autoimmune diseases have established an association between vitamin D deficiency and many autoimmune diseases6,7,8. Specifically, evidence indicates that relationships exist between osteoporosis, elevated disease activity, and increased bone resorption in AS and vitamin D metabolism9,10. The many known effects of vitamin D on the inflammatory and immune systems encouraged us to investigate a possible role of vitamin D binding protein (DBP) on the development of AS. DBP is also called DBP macrophage-activating factor (DBP-MAF), which mediates bone resorption by osteoclast differentiation and direct activation of osteoclasts11. In addition, it showed many other important biological roles, including actin scavenging, macrophage activation, and neutrophil chemotaxis11,12,13. The DBP gene is located on chromosome 4 (4q12–q13) and contains 13 exons and 12 introns, and many DBP polymorphisms have been reported in Graves’ disease, chronic obstructive pulmonary disease, alcoholic liver cirrhosis, acquired immunodeficiency syndrome, rheumatic fever, and osteoporosis13. To our knowledge DBP polymorphisms have not been studied previously in AS. We investigated the possible association between DBP polymorphisms and AS susceptibility.
MATERIALS AND METHODS
Study population and clinical evaluation
A total of 462 unrelated Korean subjects including 223 patients with AS and 239 healthy controls were recruited from Hanyang University Hospital and Eulji University Hospital. All patients showed clinical evidence of primary AS and fulfilled the modified New York criteria. Sacroiliitis was confirmed by qualified radiologists, and AS was diagnosed by qualified rheumatologists. All patients were > 16 years old. All cases of uveitis were diagnosed by ophthalmologists, and peripheral arthritis was defined as the presence of swelling and/or limitation of motion, and/or previous swelling in at least 1 peripheral joint. Genomic DNA was prepared from peripheral venous blood using standard methods.
The study protocol was approved by the institutional review boards of Hanyang University Hospital and Eulji University Hospital. All participants provided informed consent prior to enrollment.
Genotyping
Eight single-nucleotide polymorphisms (SNP; rs705117, rs2282679, rs4588, rs4752, rs222016, rs222020, rs3733359, and rs16847024) were selected based on the location of the SNP in DBP genes and minor allele frequency (MAF; > 0.05). Two SNP (rs4752, rs3733359) were selected because they are located in the exon, and rs16847024 was selected because it is located in the regulatory region. Three SNP (rs705117, rs222016, rs222020) located in the intron were selected by MAF (> 0.05). The other 2 SNP (rs2282679, rs4588) were selected because they had been reported as significant SNP in other diseases14,15. One SNP (rs7041) was excluded because it was not in Hardy-Weinberg equilibrium (p < 0.05).
Genomic DNA was prepared from EDTA whole blood with a commercial DNA extraction kit (Gentra Systems Inc., Minneapolis, MN, USA). Genotyping of the DBP SNP was done using the Illumina GoldenGate chemistry (Illumina, San Diego, CA, USA) and Sentrix Array Matrix technology on the BeadStation 500GX, according to the protocol supplied. Genotype calling was done with the Illumina BeadStudio software, and each call was confirmed manually.
Immobilization of genomic DNA on streptavidin-coated magnetic beads
Genomic DNA (5 μl at 500 ng/μl) was mixed with 5 μl of photobiotin (MSI; Illumina) and 15 μl of mineral oil, and incubated at 95°C for 30 min.
Oligo extension and ligation
Nonhybridized oligonucleotides were removed by washing, and allele-specific extension plates were made. Extension was carried out at 37°C overnight. After washing, 37 μl of master mix for extension/ligation (MEL; Illumina) was added to the extension products, and incubated for 15 min at 45°C.
Polymerase chain reaction (PCR) amplification
After extension and ligation, beads were washed with universal buffer 1 (Illumina), resuspended in 35 μl of elution buffer (Illumina), and heated at 95°C for 2 min to release the ligated products. Resulting supernatant was then used in 60 μl PCR preparations using the following cycle sequence: 10 min at 37°C, 3 min at 98°C, 34 amplification cycles (95°C for 35 s, 56°C for 35 s, and 72°C for 2 min), then 10 min at 72°C and cooling to 4°C for 5 min. The 3 universal PCR primers (P1, P2, and P3) were labeled with Cy3, Cy5, and biotin, respectively.
Preparation of PCR products
Double-stranded PCR products were immobilized on paramagnetic particles by adding 20 μl of Paramagnetic Particle B Reagent (Illumina) to 60 μl of each PCR mixture, and incubating at room temperature for a minimum of 60 min. The bound PCR products were washed with universal buffer 2 (UB2; Illumina) and then denatured by adding 30 μl of 0.1 N NaOH. After 1 minute at room temperature, 25 μl of the released ssDNA was neutralized with 25 μl of hybridization reagent (Illumina) and hybridized to arrays.
Array hybridization and imaging
Arrays were hydrated in UB2 for 3 min at room temperature, preconditioned in 0.1 N NaOH for 30 s, and returned to UB2 reagent for at least a minute to neutralize the NaOH. The pretreated arrays were exposed to the labeled single-stranded DNA samples. Hybridization was conducted using a temperature gradient from 60°C to 45°C over ∼ 12 h. The hybridized arrays were kept at 45°C until they were processed, which involved rinsing twice in UB2 and once with IS1 (Illumina) at room temperature under mild agitation. They were then imaged at a resolution of 0.8 microns using a BeadArray Reader (Illumina). PMT settings were optimized for dynamic range, channel balance, and signal-to-noise ratio. Cy3 and Cy5 dyes were excited using lasers emitting at 532 nm and 635 nm, respectively.
Statistical analysis
The genotype frequencies of each polymorphism were calculated and compliances with Hardy-Weinberg equilibrium were evaluated to check data quality and to assess genotyping errors. Linkage disequilibrium (LD) analyses by pairwise comparison of biallelic loci and haplotypes and frequencies were constructed using an EM algorithm and genotyped SNP. Case-control haplotype analyses were performed using permutation tests. The chi-square test (with a chi-square distribution and 1° of freedom) was used to compare observed numbers of subjects with each genotype with those expected. The haplotypes and their frequencies were calculated by the expectation-maximization algorithm using SNPStats, SNPAnalyzer (ISTECH Inc., Goyang, Korea).
Associations between AS susceptibility and phenotype of AS and individual SNP were quantified using OR and corresponding 95% CI using unconditional logistic regression analysis (adjusting for sex as a covariate). All analyses were performed assuming dominant, recessive, and codominant allelic effects for each polymorphism. In the dominant model, the heterozygous variant and the corresponding rare homozygote were combined. In the recessive model, the variant was defined as the rare homozygote only. In the codominant model each genotypic variant had the same effect, and in the allele model only the rare allele was responsible for the effect. The likelihood ratio test was used to assess the effect of each SNP at the 5% significance level. To address the multiple testing problem, we controlled false discovery rate and used the Benjamini and Hochberg method in R software (http://www.r-project.org).
RESULTS
In total, 462 individuals (223 patients with AS and 239 controls) were genotyped. The basic clinical characteristics of patients with AS and controls are summarized in Table 1. The 2 groups exhibited similar age and sex distributions. Of the patients with AS, 95.3% were HLA-B27-positive, 24% had uveitis, and 41% had peripheral arthritis. Figure 1 shows the LD pattern for the DBP gene. Two haplotype blocks were defined in our analyses: block 1 (10 kb, SNP 1–3) and block 2 (15 kb, SNP 5–8).
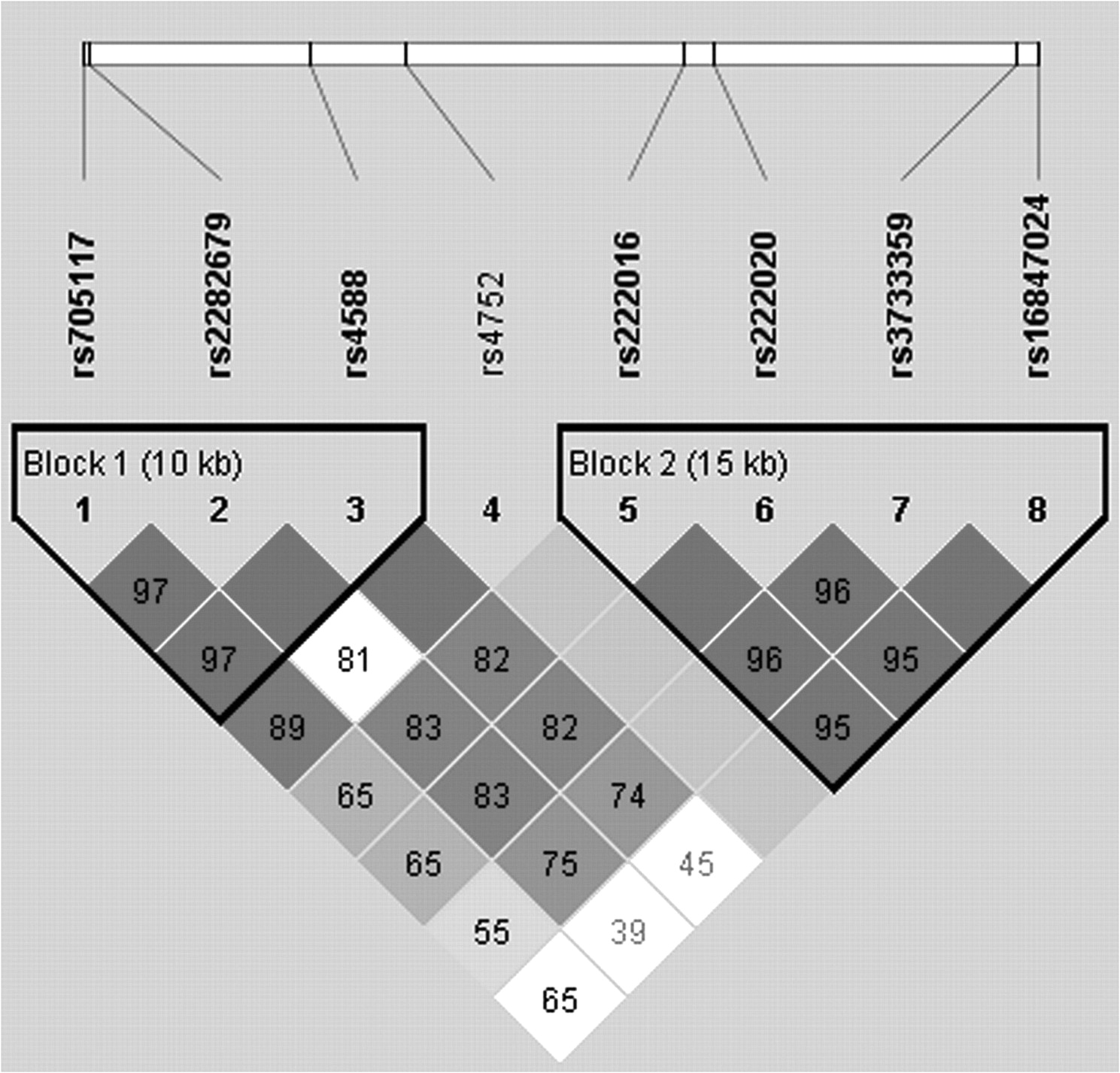
Linkage disequilibrium pattern in the vitamin D binding protein gene.
Clinical characteristics of the study subjects. Data are means ± SD; clinical features are number (%).
First, we compared the genotype frequencies of patients with AS and controls, but we found no significant association between the 8 DBP polymorphisms and AS susceptibility (Table 2).
Genotype and allele frequencies of the DBP gene SNP in patients with AS and controls.
Next, we conducted subgroup analysis in patients with AS to determine whether DBP polymorphisms were associated with the development of peripheral arthritis. Interestingly, G alleles at rs222016 and rs222020 (OR 0.63, 95% CI 0.42–0.95, p = 0.03; OR 0.63, 95% CI 0.42–0.95, p = 0.03, respectively) and A allele at rs3733359 (OR 0.59, 95% CI 0.39–0.90, p = 0.01) showed the decreased risk of peripheral arthritis (Table 3). In addition, we investigated the association between DBP polymorphisms and the development of uveitis in AS (Table 4). The rs4752 polymorphisms were associated with a development of uveitis in patients with AS. G allele at rs4752 showed the increased risk of uveitis (OR 2.04, 95% CI 1.12–3.72, p = 0.02).
Logistic analysis of DBP polymorphisms and the risk of peripheral arthritis among patients with AS.
Logistic analysis of the risk of uveitis among patients with AS for different DBP polymorphisms.
Haplotype analyses were also performed using 4 SNP (rs4752, rs222016, rs222020, rs3733359, sequentially). The analyses showed an association with the development of peripheral arthritis or uveitis. The frequencies of 4 possible haplotypes are shown in Table 5. The results showed that haplotype 2 (AGGA) protected against the development of peripheral arthritis in patients with AS (p = 0.01) and that haplotype 3 (GAAG) was associated with an increased likelihood of uveitis (p = 0.02; Table 5).
The haplotype frequencies of the DBP SNP for AS patients with or without peripheral arthritis/uveitis.
DISCUSSION
The etiology of AS is unclear, but the roles that genetic factors play in susceptibility to AS and in disease severity are well established. However, only 1%–5% of HLA-B27-positive individuals develop AS1,2,3, and thus the genetic influences of non-MHC genes have been emphasized4,5. Recently, immune modulatory actions of vitamin D and an association between vitamin D deficiency and several autoimmune diseases have been reported6,7,8. Since vitamin D has immunosuppressant properties, supplementation with this vitamin has been suggested as a treatment for Th1-mediated autoimmune diseases6. Further, it has also been suggested that vitamin D plays an important role in the pathogenesis of AS. Lange, et al found that high disease activity is associated with an alteration in vitamin D metabolism that increases bone resorption9, and another study provided evidence that vitamin D deficiency leads to osteoporosis in AS by increasing inflammatory activity10. In addition, Lauridsen, et al detected relationships between the DBP phenotype and plasma concentrations of 25(OH) vitamin D3 and 1,25(OH)2 vitamin D316, and Ahn, et al in a metaanalysis of 5 genome-wide association studies among Europeans showed that SNP in the DBP gene are associated with different 25(OH)D concentrations14. It seems, therefore, that the activity of vitamin D may depend on the genetic makeup of the DBP gene. DBP is a highly polymorphic protein, and associations have been reported between DBP polymorphisms and susceptibility to several diseases13. A few studies have also examined the association between DBP polymorphisms and rheumatic diseases, but no similar investigation has been conducted in AS.
Even though we found no significant relationship between the 8 DBP polymorphisms and AS susceptibility, we did uncover associations between the DBP polymorphisms and the development of peripheral arthritis and uveitis in AS. Among extraarticular manifestations in AS, inflammatory bowel disease (IBD) and psoriasis were extremely rare in our cohort. So we did not analyze associations between these manifestations and DBP gene polymorphisms. According to a recent report, there is a high incidence of peripheral arthritis among Korean patients with AS, and patients with AS who have peripheral arthritis are younger and more frequently have enthesitis17. Interestingly, Singh, et al suggested that the etiology of peripheral arthritis in AS involves different factors and that AS with peripheral arthritis should be considered a distinct form of AS18. Peripheral arthritis is known to be more prevalent in patients with uveitis18,19, and Zeboulon, et al reported that uveitis and arthritis have common genetic predisposing factors and similar pathophysiologies20. However, the actual causative mechanism has not been identified. Increased expressions of adhesion molecules have been reported in patients with spondyloarthropathies, especially in patients with peripheral arthritis21. The adhesion molecules — intercellular adhesion molecule-1 (ICAM-1) and leukocyte functional antigen-1 — play critical roles in the immune mechanisms of uveitis in AS22. Martinesi, et al reported that vitamin D derivatives significantly downregulate ICAM-1 levels in healthy controls and patients with IBD, and suggested that vitamin D derivatives could be considered therapeutic in IBD23.
Ahn, et al reported that 3 SNP in DBP (rs2282679, rs7041, rs1155563) are significantly associated with differences in 25(OH)D levels14. A systematic review of the association between DBP SNP and 25(OH)D showed that rs4588 and rs7041 were significantly associated with 25(OH)D concentrations15. We analyzed rs2282679 and rs4588, but failed to observe any association with susceptibility to AS.
It is a limitation of our study that we did not investigate possible associations between serum 25(OH)D levels and DBP polymorphisms in AS. Further study regarding the associations between vitamin D deficiency in patients with peripheral arthritis or uveitis and DBP polymorphisms would be helpful for determining the role played by vitamin D in the pathogenesis of AS. However, variations in 25(OH)D measurement among laboratories, and several variables that affect 25(OH)D status such as season at time of blood collection, leisure time physical activity, total vitamin D intake, total calcium intake, and smoking made us hesitate to take a serum 25(OH)D level measurement24. We anticipate a well organized study regarding the association of serum 25(OH)D level with DBP polymorphisms in patients with AS in the near future.
Rs4752, which showed association with development of uveitis, is a synonymous SNP. According to a recent review, synonymous SNP can also change messenger RNA splicing, stability, and protein folding. Therefore these changes may have an effect on cellular response to therapeutic targets25. Chen, et al reported that synonymous SNP and nonsynonymous SNP have similar likelihood and effect size for disease association26.
Sample sizes in subgroup analyses are often small and subgroup analyses therefore usually lack statistical power. Our results showed an association between DBP polymorphisms and the development of peripheral arthritis and uveitis in spite of an insufficient sample size. The modest p values may make conclusions difficult, so additional larger-scale studies need to be performed. Additionally, DBP has a regulatory role in bone formation and remodeling11, and DBP modulates inflammatory and immune mechanisms. Taking these roles into consideration, it is necessary to perform genetic and functional studies to reveal association between DBP and AS in large cohorts.
Haplotype analyses revealed that haplotype 2 (AGGA) was protective from the development of peripheral arthritis and haplotype 3 (GAAG) increased the risk of uveitis. Since the p values were modest, further fine mapping is warranted to validate whether the association was due to linkage disequilibrium or a true association with functionally relevant polymorphisms.
Our study provides supportive evidence of associations between DBP polymorphisms and development of peripheral arthritis or uveitis in Korean patients with AS.
Footnotes
-
Supported by the Korean Health 21 R&D Project of the Ministry of Health and Welfare (#01-PJ3-PG6-01GN09-003).
- Accepted for publication June 30, 2011.








{kind=link}