

Abstract
Objective. To determine the role of natural killer (NK)-22 cells in the pathogenesis of rheumatoid arthritis (RA).
Methods. Using flow cytometry, the proportions of NK-22 cells and intracellular contents of perforin, granzyme B, and interferon-γ (IFN-γ) were determined in the peripheral blood (PB) and synovial fluid (SF) of patients with RA and healthy individuals. The levels of interleukin 22 (IL-22) and tumor necrosis factor-α (TNF-α) in the NK-22 supernatant and gene expressions were measured using ELISA and QuantiGene Plex assay, respectively. The effect of NK-22 supernatant on the proliferation of fibroblast-like synoviocytes (FLS) and recombinant human IL-22 (rhIL-22) on the production of monocyte chemoattractant protein 1 (MCP-1) by RA FLS was detected using the yellow tetrazolium salt method and ELISA, respectively. The relationship between the proportions of NK-22 cells and disease activity was analyzed.
Results. NKp44 and CCR6 were expressed in a larger population of SF NK cells than in the PB NK cells of patients with RA. NK-22 cells produce low content of perforin, granzyme B, and IFN-γ. NK-22 cells in vitro can secrete IL-22 and TNF-α and there was increased messenger RNA coding for IL-22 and TNF-α. NK-22 supernatant can induce the proliferation of RA FLS. Addition of IL-22 antibody plus TNF-α antibody inhibited the proliferation of FLS induced by the NK-22 supernatant. Both rhIL-22 1 ng/ml and rhIL-22 10 ng/ml induced the production of MCP-1 by RA FLS. The NK-22 proportions were positively correlated with disease activity.
Conclusion. NK-22 cells are increased in patients with RA and might play a role in the pathogenesis of RA through the production of IL-22 and TNF-α. The proportion of NK-22 cells and disease activity were highly correlated.
Rheumatoid arthritis (RA) is an autoimmune disease in which a chronic inflammatory response develops, particularly in synovial joints. There is compelling evidence that the activation of T-cells is central to the development of the inflammation1, but the mechanisms leading to the longstanding T-cell activation are poorly understood. In several animal models of autoimmunity, natural killer (NK) cells have been shown to control T-cell activation2,3. NK cells are important components of the innate immune system and account for 10%–15% of peripheral blood lymphocytes (PBL). NK cells are defined by the expression of CD56 and the lack of expression of CD34. NK cells have a number of effector functions, including recognition and lysis of virus-infected or tumor cells and production of immunoregulatory cytokines. Their activity is regulated by integration of both activatory and inhibitory signals from a wide range of cell-surface receptors. Better understanding of the biology of NK receptors has led to greater knowledge about their physiological role, and consequently their role in disease can now be investigated.
The receptors involved in the regulation of NK cell activity fall into 3 categories5. Natural cytotoxicity receptors, including NKp30, NKp44, and NKp46, bind mostly undefined ligands on potential target cells. The resulting activating signal is essential for NK cell-mediated cytotoxicity. In RA, several studies have investigated the potential involvement of NK cells. Two subsets of NK cells in peripheral blood have been recognized6. The majority (CD56dim NK cells) express moderate levels of CD56 and high levels of CD16. The minor subset of NK cells (CD56bright NK cells) accounts for only 10% of circulating NK cells. These cells express high levels of CD56, CD94, and NKG2A. The 2 subsets of NK cells show important functional differences; the CD56dim subset has superior cytotoxic capacity, while the CD56bright subset has greater ability to produce proinflammatory cytokines including TNF-α. Researchers have demonstrated that the CD56bright subset of NK cells is greatly expanded in the synovial fluid (SF) of patients with inflammatory arthritis7. These NK cells are capable of interacting in a reciprocal fashion with the CD14 mononuclear cell population, thereby promoting the development or maintenance of inflammation.
The phenotype and function of NK cells in the SF of patients with chronic joint inflammation were also investigated8. Compared to the PB NK subset, the synovial NK cells expressed a higher degree of activation markers including CD69 and NKp44; synovial NK cells were also upregulated on CD56bright NK cells in the PB of patients. Recently, a human NK cell subset called NK-22 cells was reported in patients with inflammatory bowel disease (IBD)9. Its phenotype is NKp44+CCR6+NK cell. NK-22 cells can secrete many cytokines including interleukin 22 (IL-22), which has been shown to activate signal transducer and activator of transcription 3 (STAT3)10, a known inducer of cell survival and proliferation, as well as STAT1. NK-22 cells play a crucial role in the development of IBD through the production of IL-22. It has been proved that high levels of IL-22 were expressed in RA synovial tissues11. IL-22 can induce the proliferation and chemokine production of synovial fibroblasts.
In our study, we examined whether NK-22 cells exist in patients with RA and whether they play a role in the pathogenesis of RA. Determining the function of NK-22 cells in RA and any correlation between NK-22 cells and disease activity is important to further understanding of RA pathogenesis and future treatment.
MATERIALS AND METHODS
Patients, controls, and cell separation
Twenty patients and 20 healthy controls matched for sex ratio and mean age were included in our study. The demographic and key clinical information of the patients with RA is summarized in Table 1. All the patients had fulfilled the revised American College of Rheumatology criteria for the disease. Ten patients had undergone therapeutic aspiration of SF at the time of analysis. Informed consent was obtained from all subjects before sample collection. All SF samples were compared to PB samples from the same patient. For all patients, SF was obtained from 1 knee joint. All samples were collected in EDTA tubes. Mononuclear cells were isolated within 2 hours of sample collection by density gradient centrifugation.
Patient demographic details.
Monoclonal antibodies and flow cytometry
Anti-CD56-FITC antibodies, anti-CD3-PE Cy5 antibodies, and isotype-matched controls (mouse IgG1-FITC antibodies and mouse IgG1-PE Cy5 antibodies) were purchased from BD Biosciences (San Jose, CA, USA). Anti CCR6-PE antibodies, anti NKp44-APC antibodies, and isotype-matched controls (mouse IgG1– PE antibodies and mouse IgG1– APC antibodies) were purchased from R&D Systems (Minneapolis, MN, USA). Anti-perforin-PE antibodies, anti-granzyme B-PE antibodies, anti-IFN-γ-PE antibodies, and isotype-matched controls were purchased from BD Biosciences. NK cells were defined as CD56+ CD3– cells within the lymphocyte gate. NK-22 cells were NKp44+CCR6+NK cells. Data in the text are expressed as median and ranges. Expression levels are given as the geometric mean of fluorescence intensity. NK-22 cells and normal NK cells were sorted with a BD FACSAria 1™ flow cytometer; 2 × 104 cells were collected.
Cell culture and identification
NK-22 cell cultures were maintained in complete medium consisting of RPMI-1640 (Invitrogen) supplemented with 10% heat-inactivated fetal bovine serum, 2 mM L-glutamine, 100 U/ml penicillin, and 100 μg/ml streptomycin (Sigma-Aldrich, St. Louis, MO, USA). The purity of the NK-22 cells was measured using flow cytometry. RA fibro blast-like synoviocytes (FLS) were isolated and cultured in vitro. Briefly, synovial tissues were minced into 2–3 mm pieces and treated for 2 hours with 2 mg/ml type I collagenase in Dulbecco’s modified Eagle’s medium (DMEM; Sigma) at 37°C with continuous shaking. After removal of the tissue debris through a nylon mesh, the cells were washed with Hanks’ balanced salt solution and then harvested in 25 cm2 flasks in DMEM supplemented with 10% heat-inactivated fetal bovine serum (FBS; Sigma). All in vitro experiments were conducted using primary RA FLS cultures between passages 3 and 5. RA FLS were stained with vimentin antibodies for identification.
ELISA and cell proliferation assay
After 2 weeks in in vitro culture, the NK-22 cells were stimulated with 20 ng/ml phorbol 12-myristate 13-acetate (PMA) and 0.5 μM ionomycin (Sigma) for 4 h. The concentrations of IL-22 and TNF-α in the PB and SF of patients and in NK-22 cell culture supernatants were determined using an ELISA from ExCell Biology Inc. (Shanghai, China).
RA FLS were incubated with variable concentrations of rhIL-22 for 72 h. The concentration of MCP-1 in cell culture supernatants was determined by an ELISA (ExCell Biology). Cell proliferation was assessed with a yellow tetrazolium salt (MTT) assay. RA FLS were seeded at 1 × 105 cells/well in 96-well plates. After 1, 2, 3, and 4 days of incubation with NK-22 cell culture supernatants, MTT (5 mg/ml) was added to each well. After 4 h, the optical density of each well was determined using a microplate reader set at 490 nm. The supernatant of NK-22 cells was diluted with RPMI-1640. After incubation of NK-22 cell culture supernatants with IL-22 antibody (50 μg/ml) and TNF-α antibody (1.2 μg/ml) for 24 h, cell proliferation was assessed with the MTT assay.
Detection of gene expression of IL-22 and TNF-α with a QuantiGene Plex (QGP) 2.0 assay
Gene expressions of IL-22 and TNF-α in NK-22 cells were detected with a QGP assay (Panomics Inc., Santa Clara, CA, USA). Targeted genes and housekeeping genes were synthesized by Panomics Inc. A working Plex Set was dispensed into a hybridization plate. A volume of 80 μl of NK-22 cell suspension (105/ml) was added to each assay well. Simultaneously, 80 μl of the diluted lysis mixture was added to 3 of the wells as assay background control. The hybridization plate was incubated for 18–22 h in a Vortemp shaking incubator at 54° ± 1°C and 600 rpm. The hybridization mixture was transferred to a magnetic separation plate. The plate was then washed 3 times and 100 μl of the preamplifier reagents were added to each assay well. The magnetic separation plate was incubated for 1 h at 50°C ± 1°C and 600 rpm. After washing the plate, 100 μl of amplifier reagents was added. The magnetic separation plate was incubated for 1 h at 50°C ± 1°C and 600 rpm. The plate was washed and then 100 μl of label probe working reagents was added. The magnetic separation plate was incubated for 1 h at 50°C ± 1°C and 600 rpm. The plate was then washed and 100 μl of SAPE working reagents was added to each assay well. The plate was shaken at 800 rpm for 1 min followed by 600 rpm for 30 min. The plate was washed and 130 μl of SAPE wash buffer was added. The magnetic separation plate was sealed and placed on a shaking platform at 800 rpm for 2–3 min, and read immediately.
Statistics
All results are expressed as a mean ± SEM or median ± interquartile range. The Wilcoxon signed-rank test was used to compare the paired data (patient PB and SF), while the Mann-Whitney U test was used to compare the data derived from the PB of patients and healthy controls. Statistical significance in cell proliferation was determined using repeated measures or a 1-way analysis of variance followed by a Bonferroni test. Spearman rank correlations were used to determine the relationship between variables. The reported p values are 2-tailed; significance was set at p < 0.05.
RESULTS
NK-22 cells exist in the PB and SF of patients with RA
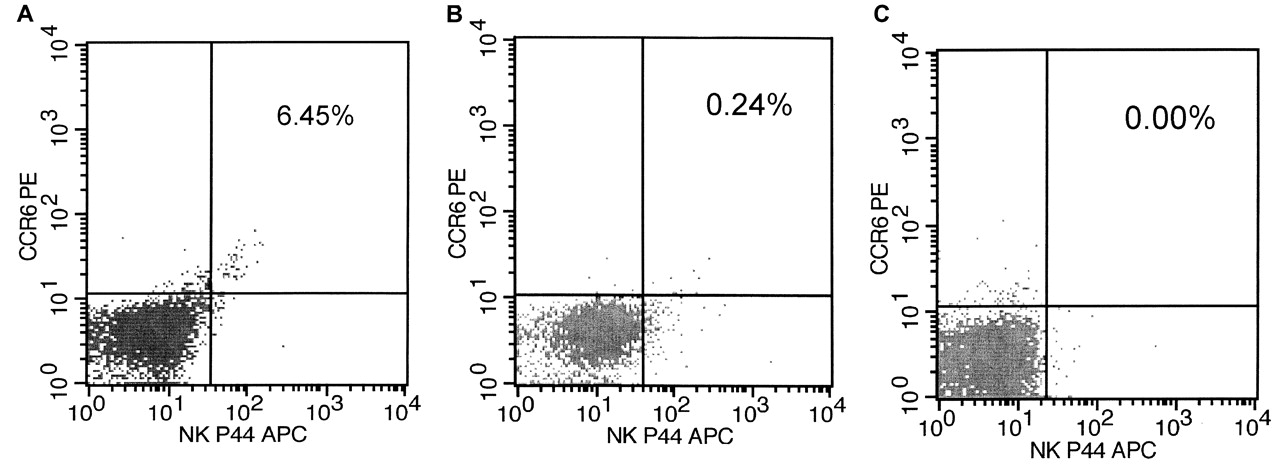
The phenotype of NK-22 cells was analyzed using 4-color flow cytometric analysis (Figure 1). The results presented below were comparisons of SF NK-22 cells and PB NK-22 cells from patients with RA, as well as comparisons with blood NK cells from healthy controls. The frequencies of total NK-22 cells differed between the SF and PB of matched patients (z = −3.781, p < 0.01). In PB, the frequency of NK-22 cells ranged from 0.42% to 2.44% (median 1.195%) of the total lymphocytes, while SF ranged from 3.57% to 6.45% (median 4.165%). Similarly, the percentages of PB NK-22 cells from all patients and controls were significantly different (z = −5.674, p < 0.01). The frequency of NK-22 cells in patients ranged from 0.05% to 2.44% (median = 0.690%), and in the controls from 0% to 0.03% (median = 0.000%).

Expression of natural killer (NK)p44 cells and CCR6 receptors on mononuclear cells isolated from peripheral blood (A) and synovial fluid (B) of patients with RA and healthy controls (C). Cells were stained for CD3, CD56, NKp44, and CCR6 receptors. NK cells were defined by gating for CD3– CD56+ lymphocytes. The data show the proportion of NK-22 cells within the NK gate from 1 representative patient and 1 control.
Lytic mediators and IFN-γ produced by NK-22 cells
NK-22 cells have low intracellular content of perforin and granzyme B. After stimulation with PMA and ionomycin, NK-22 cells produced low levels of IFN-γ (Figure 2).

Natural killer (NK)p44+NK cells produce low amounts of perforin, granzyme B, and IFN-γ. Peripheral blood mononuclear cells were stained for cell-surface expression of CD3, CD56, and NKp44, and counterstained for intracellular perforin, granzyme B, and IFN-γ. A gate was applied on CD3–CD56+ NK cells.
Level of IL-22 and TNF-α in the supernatant of NK-22 cells and gene expressions of IL-22 and TNF-α in NK-22 cells
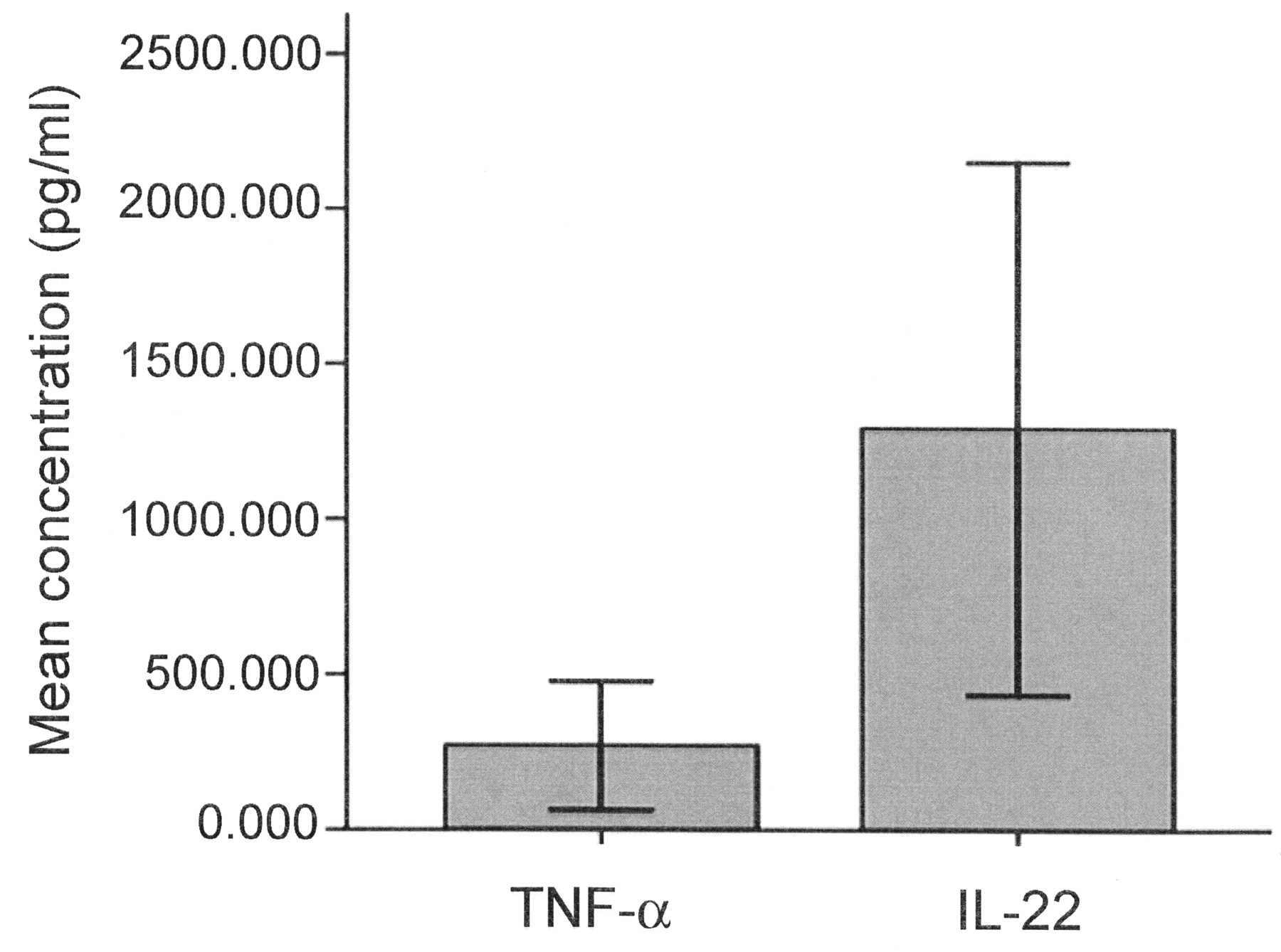
The purity of NK-22 cells sorted by flow cytometry was greater than 90% (Figure 3). The levels of IL-22 and TNF-α in the supernatant of NK-22 culture solution were 1293.971 ± 345.048 pg/ml and 272.238 ± 83.471 pg/ml, respectively (Figure 4). Gene expressions of IL-22 and TNF-α in NK-22 cells were significantly higher than in normal NK cells (Table 2).
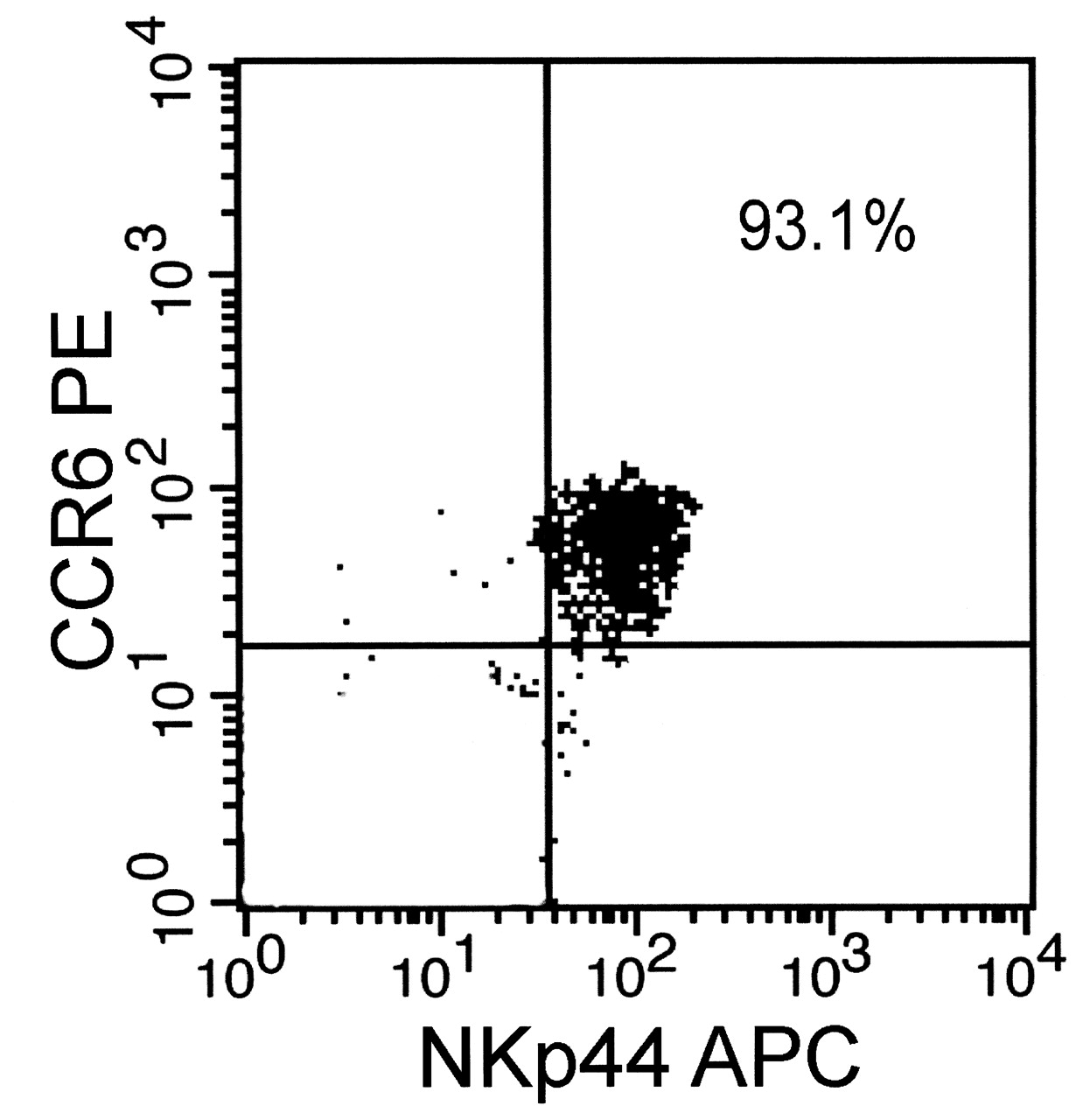
The purity of natural killer (NK)-22 cells by flow cytometry.

The concentration of IL-22 and TNF-α in the supernatant of culture solution after stimulation with phorbol 12-myristate 13-acetate and ionomycin.
Levels of gene expression in interleukin 22 (IL-22) and tumor necrosis factor-α (TNF-α).
Effect of NK-22 supernatant on proliferation of FLS
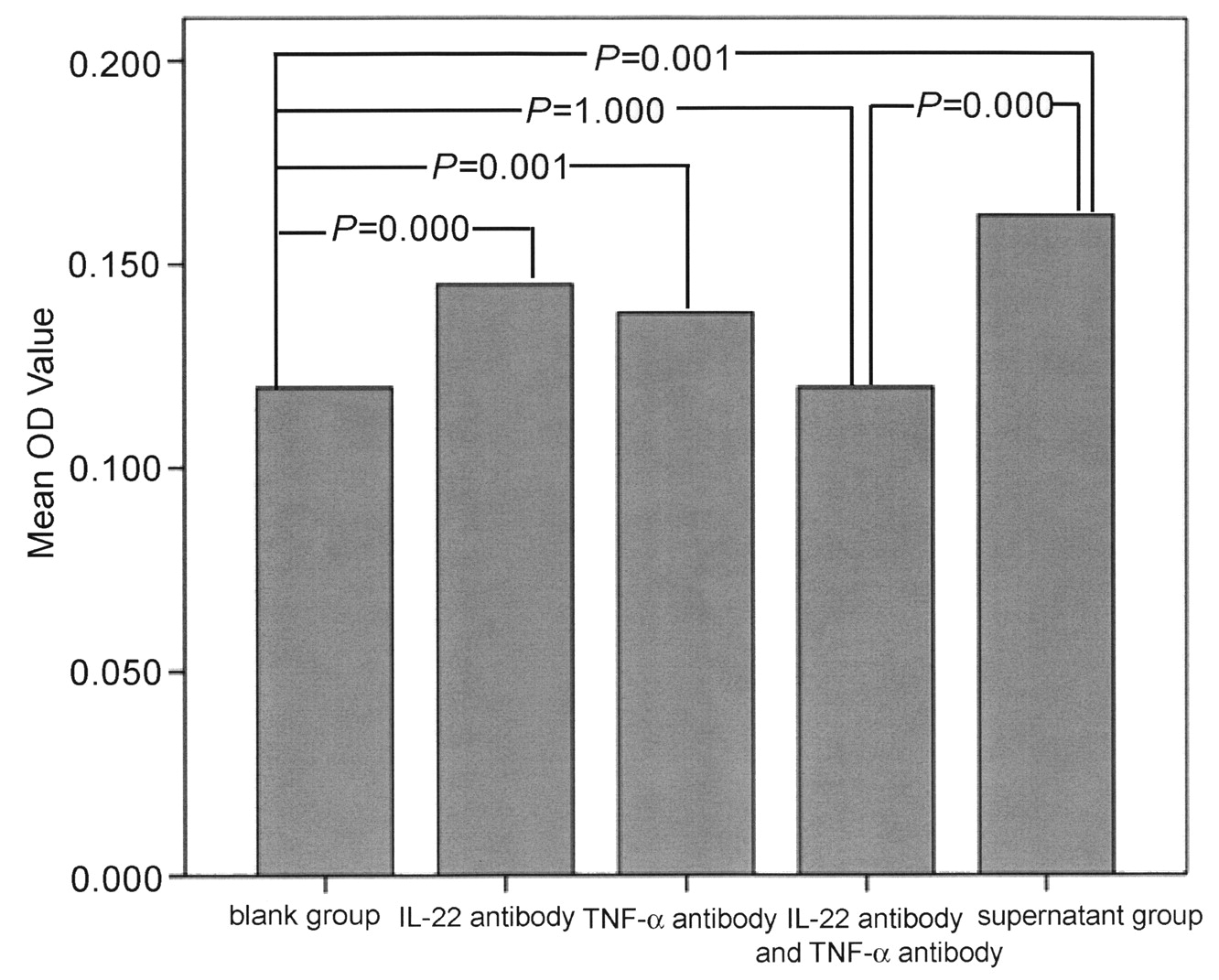
The supernatant of NK-22 cells contained 941.82 pg/ml of IL-22 and 368.1 pg/ml of TNF-α. The purity of RA FLS was nearly 100%, according to vimentin staining. Twenty-four hours after the addition of supernatant, FLS proliferated significantly. At 72 h and 96 h, groups of FLS at 3 different concentrations of supernatant proliferated quickly compared with the blank group (Figure 5). Cell proliferation was obviously inhibited after the addition of IL-22 antibody and TNF-α antibody for 24 h, compared with the NK-22 supernatant group (p < 0.05). There was no significant difference between IL-22 antibody plus TNF-α antibody group and blank group (Figure 6).
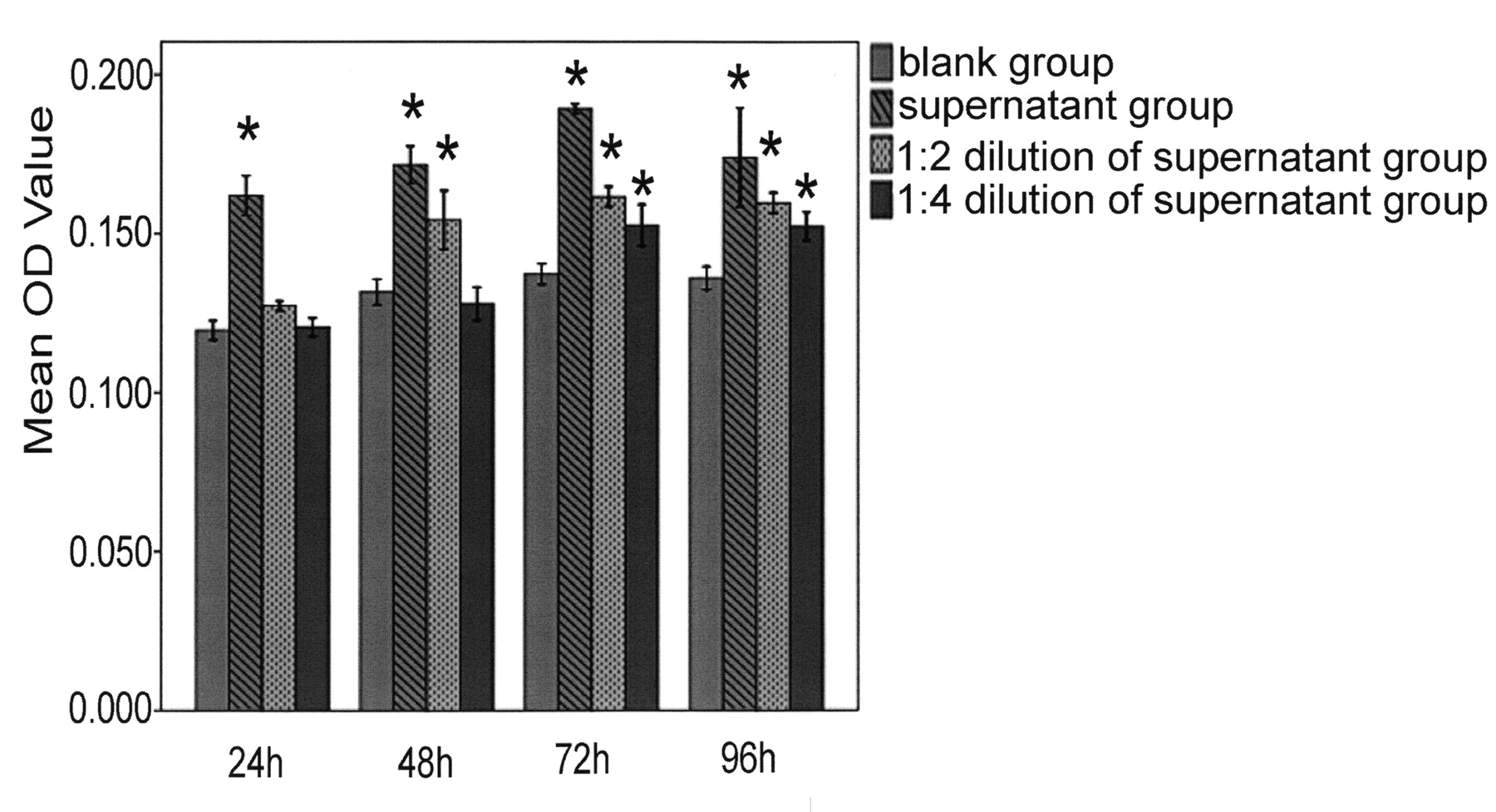
The effect of natural killer-22 supernatant on the proliferation of fibroblast-like synoviocytes. The final concentrations of IL-22 and TNF-α in the 1:2 dilution group were 470.91 pg/ml and 184.05 pg/ml, respectively. *p < 0.01 compared with the blank group. Error bars show ± 1 SD.
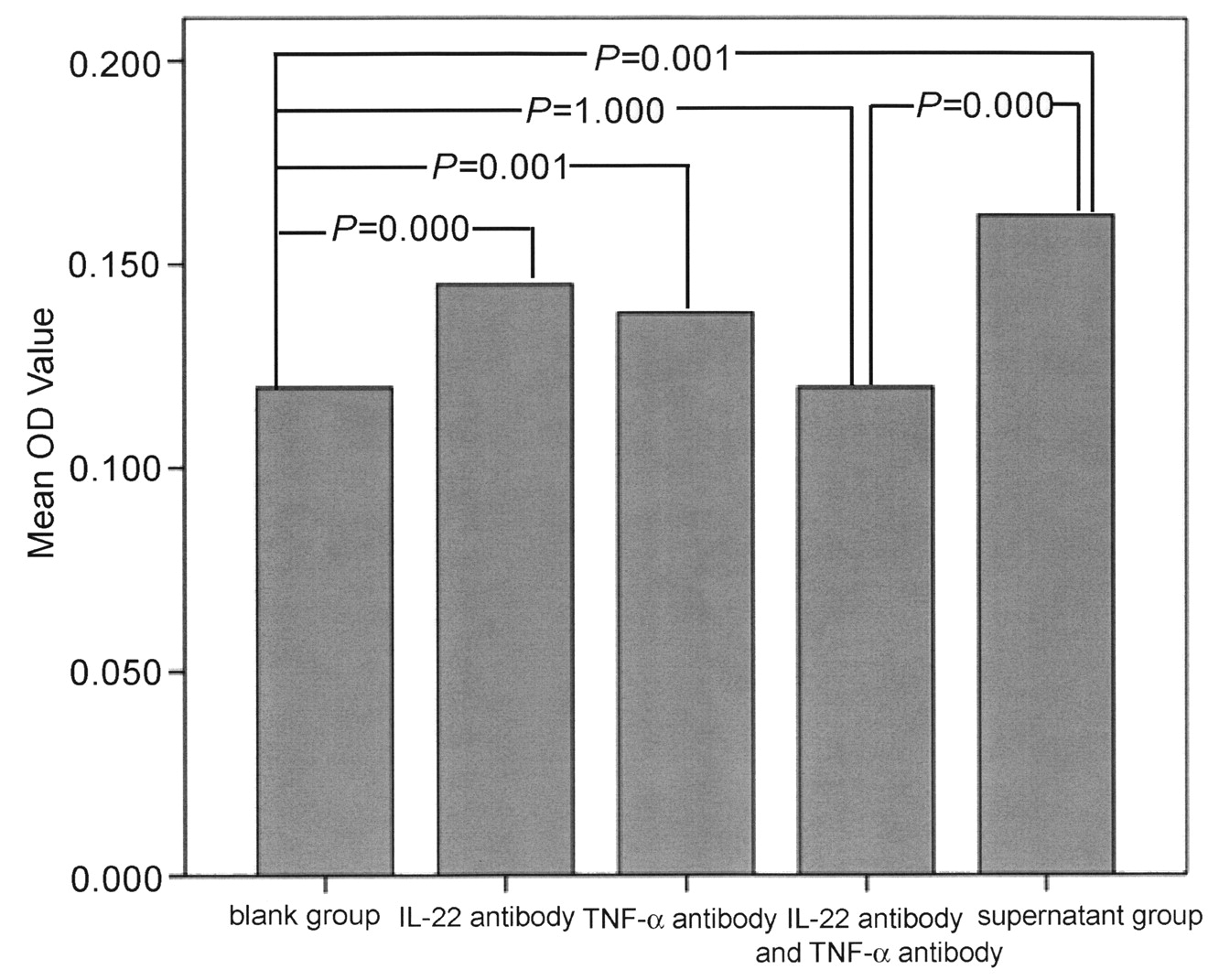
The effect of IL-22 antibody and TNF-α antibody on the proliferation of RA fibroblast-like synoviocytes induced by the natural killer-22 supernatant after 24 h.
Effects of rhIL-22 on RA FLS by production of chemokines
Figure 7 shows that treatment with rhIL-22 increased the production of monocyte chemoattractant proteins (MCP-1) by RA FLS. Both rhIL-22 1 ng/ml and rhIL-22 10 ng/ml induced the production of MCP-1 by RA FLS compared with media controls (p < 0.01). Moreover, 10 ng/ml rhIL-22 induced higher production of MCP-1 than 1 ng/ml rhIL-22 (p < 0.01).

Increased RA fibroblast-like synoviocyte production of MCP-1 by IL-22. MCP-1 synthesis in the supernatant was determined by ELISA after 72 h. *p < 0.05 vs media control. #p < 0.05 vs 10 ng/ml rhIL-22.
Correlation between proportion of NK-22 cells and levels of IL-22 and TNF-α in PB and SF of patients with RA
Levels of TNF-α and IL-22 in the PB of 20 patients with RA were 313.059 ± 90.513 pg/ml and 5.34 ± 1.614 ng/ml, respectively, while levels of IL-22 and TNF-α in the SF of 10 patients with RA were 681.245 ± 65.996 pg/ml and 7.22 ± 1.249 ng/ml. There was positive correlation between NK-22 cells in PB and the levels of IL-22 and TNF-α in PB (r = 0.869, p = 0.000; r = 0.957, p = 0.000). There was also positive correlation between NK-22 cells in SF and the levels of IL-22 and TNF-α in SF (r = 0.930, p = 0.000; r = 0.967, p = 0.000; Figure 8).

Correlation between NK-22 cells and levels of IL-22 and TNF-α in synovial fluid. X axes refer to the proportion of NK-22 cells (percentage).
Correlation between NK-22 cells and disease activity
The proportions of NK-22 in PB were positively correlated with Disease Activity Score 28 (DAS28; r = 0.975, p = 0.000) and the levels of C-reactive protein (CRP; r = 0.484, p = 0.031), while the proportions of NK-22 in SF were positively related to DAS28 (r = 0.918, p = 0.000) and the levels of rheumatoid factor (r = 0.784, p = 0.007).
DISCUSSION
Strong evidence indicates that the innate immune system, and in particular NK cells, influences subsequent adaptive immune responses. Apart from their cytotoxic activity, NK cells secrete a variety of cytokines and modulate T-cell responses, both in vitro and in autoimmunity animal models2,3. Clearly, NK cell functions must be carefully regulated to prevent damage to normal tissues or the indiscriminate release of cytokines resulting in inappropriate activation of the adaptive immune system. Since the early 1980s, it has been known that there were decreased NK cell numbers and impairment of NK cell function in the PB of patients with autoimmune diseases, including RA12. However, rather than consisting of a homogeneous population, NK cell subsets with distinct functional characteristics have been defined in both mice and humans. Experiments in vitro have shown that NK cells in the SF of patients with RA exhibit a CD56bright, CD94bright, CD158negative phenotype13. Moreover, it has been found that the synovial NK cells of patients with arthritis express a higher degree of activation markers including CD69 and NKp448.
We report the presence of a new subtype of NK cells, called NK-22 cells, in the SF and PB of patients with RA, whereas it was absent from healthy individuals. Our results are consistent with the findings of previous studies of NK cells at inflammatory sites. Our data demonstrate that the number of NK-22 cells in SF is obviously greater than that in the PB of patients with RA. NK-22 cells can coexpress the NKp44 receptor and CCR6 molecules. NKp44 is a member of the natural cytotoxicity receptor family, which is expressed following NK cell activation14. The overall expression of NKp44 was elevated in synovial NK cells compared to PB NK cells, indicating the presence of activated NK cells in the joint8. The CCR6 molecule is a G-protein-linked transmembrane receptor15 and is a specific receptor for macrophage proinflammatory human chemokine-3α (MIP-3α), which participates in directing inflammatory cell migration and in modulating angiogenesis. So we postulated that NK-22 cells have the ability to migrate to inflammatory sites.
Although we found there are low cellular contents of lytic mediators and IFN-γ, suggesting NK-22 cells have lower cytotoxicity, NK-22 cells in vitro can secrete IL-22 and TNF-α. It has become increasingly clear that dysregulation of cytokine networks within the joints of patients with arthritis plays an important role in the maintenance of the disease16. These networks are complex, with many different cell types secreting, and expressing receptors for, a wide range of cytokines. Careful analysis has shown that TNF-α plays a central role within the networks, and that expression of other proinflammatory cytokines and growth factors, such as IL-1, IL-6, and granulocyte-macrophage colony-stimulating factor, is dependent on TNF-α17,18. It has been shown that high levels of IL-22 are expressed in both the lining and the sublining layers of RA synovial tissues11. IL-22 can induce the proliferation and chemokine production of synovial fibroblasts.
IL-22 and TNF-α are 2 important proinflammatory cytokines that are involved in the pathogenesis of RA. Because of their ability to produce cytokines, NK-22 cells may participate in the development of RA. To further investigate the role of NK-22 cells in the pathogenesis of RA, we also observed the effect of cytokines produced by NK-22 cells on the proliferation of synovial fibroblasts. The supernatant from NK-22 cell culture solution can significantly promote the proliferation of synovial fibroblasts. To evaluate the contributions of IL-22 and TNF-α or other cytokines in the NK-22 supernatant to FLS proliferation, we investigated the effect of IL-22 antibody and TNF-α antibody on the proliferation of RA FLS induced by NK-22 supernatant. When IL-22 and TNF-α were inhibited, NK-22 supernatant-induced proliferation of RA FLS was almost completely inhibited. Our results indicated IL-22 and TNF-α were the main contributors in NK-22 supernatant to the proliferation of RA FLS. MCP-1 likely plays a pivotal role in macrophage infiltration. Previous studies have shown that MCP-1 could be secreted by RA FLS and contribute to joint destruction in RA19.
It is plausible that in our study IL-22 induced an upregulation of MCP-1. Our study showed a positive correlation between the proportion of NK-22 in PB and 28-joint Disease Activity Score (DAS28) and the level of CRP, while the proportion of NK-22 in SF was positively related to DAS28 and the level of rheumatoid factor. Moreover, the proportion of NK-22 cells was closely related to the levels of IL-22 and TNF-α in both PB and SF. The results suggest that NK-22 cells are closely related to disease activity. Patients with larger numbers of NK-22 cells had a greater degree of disease activity.
Some evidence indicates that NK-22 cells are involved in the pathogenesis of RA. Further research into the functions of the features of NK-22 cells, especially their immunoregulatory role in autoimmunity, is warranted and will likely provide new insights into the pathogenesis of autoimmune disorders.
Acknowledgment
We are grateful to all the patients who kindly donated their synovial effusions and blood. We are also grateful to Prof. Yi Zhen-shan (Nanfang Hospital) for his help with the experiments.
Footnotes
-
Supported by the Natural Science Foundation of Guangdong Province and the Doctoral Foundation of the Ministry of Education of the People’s Republic of China.
- Accepted for publication May 3, 2011.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}