

Abstract
Objective. To examine the influence of STAT4 rs7574865 gene polymorphism on patients with primary Sjögren’s syndrome (SS).
Methods. Two different cohorts were studied: 69 patients with primary SS and 296 controls from Colombia and 108 patients with primary SS and 227 controls from Germany. Samples were genotyped for the STAT4 rs7574865 single-nucleotide polymorphism with a predesigned TaqMan single-nucleotide polymorphism genotyping assay. We carried out a metaanalysis of our results combined with data published to date.
Results. Although no significant differences were observed in the allele frequencies of STAT4 rs7574865 gene polymorphism between patients and controls in Colombians (p = 0.28, OR 1.24, 95% CI 0.82–1.87) and Germans (p = 0.08, OR 1.40, 95% CI 0.96–2.02), the metaanalysis disclosed a significant effect of the T allele on disease (p = 4.7 × 10−6, OR 1.40, 95% CI 1.21–1.62).
Conclusion. These data reinforce the influence of STAT4 gene on primary SS and as a general autoimmune gene.
Sjögren’s syndrome (SS) is a chronic autoimmune disease characterized by progressive lymphocytic and plasma cell infiltration of the salivary and lachrymal glands, accompanied by the production of autoantibodies leading to xerostomia and keratoconjunctivitis sicca (sicca symptoms). SS may occur alone (primary SS; pSS) or in association with other autoimmune diseases (secondary SS; sSS), of which the most frequent are Hashimoto’s thyroiditis and rheumatoid arthritis (RA). The powerful influence of genetic predisposition on susceptibility is usually based on disease concordance rates in monozygotic twins. Although genetic studies in pSS twins have not been performed, the observed aggregation of autoimmune diseases in families of patients with pSS supports a genetic component in SS etiology1–4. The efforts to identify the genetic component of SS have relied on association studies for disease gene identification. However, robust analyses of candidate gene variants have not been undertaken and no linkage study has been reported to date. In the absence of chromosomal regions identified by linkage studies, research has focused on candidate gene approaches (by biological plausibility) rather than on positional approaches. Association with HLA genes has been largely described and confirmed5–7. Genes outside the HLA region have also been associated with pSS; nevertheless these positive associations, including PTPN228,9 and CTLA, have proven difficult to replicate. Recently, an association of STAT4 gene polymorphism (rs7574865) at 2q32 with SS has been reported12,13. Our objective was to assess the influence of STAT4 rs7574865 polymorphism on pSS in German and Colombian populations, and to evaluate the global influence of this variant in disease susceptibility with a metaanalysis.
MATERIALS AND METHODS
Study population
Peripheral blood samples were obtained from 2 unrelated cohorts of pSS patients and controls. The Colombian cohort consisted of 69 pSS patients and 296 healthy controls and the German cohort 108 pSS patients and 227 healthy controls. All patients fulfilled the international classification criteria14 for pSS. Patients and controls were all Caucasian and were matched for age (by mean age) and for sex (by frequency matching). Written informed consent was obtained from all subjects, and the study was approved by the local ethics committee of each center. Demographic characteristics of the patients and controls in each population have been described15–17.
For the metaanalysis we combined data from North American (case-control ratio 120/1112), Norwegian (case-control ratio 228/669), and Swedish (case-control ratio 140/148) cohorts12,13 with our findings. It is important to note that Nordmark, et al13 analyzed rs7582694 STAT4 gene polymorphism, which is a perfect proxy of rs7574865.
Genotyping for STAT4 polymorphism
Samples were genotyped for STAT4 rs7574865 using a polymerase chain reaction system with a predeveloped TaqMan allelic discrimination assay (Applied Biosystems, Foster City, CA, USA)18. Duplicate samples and negative controls were included to check the accuracy of genotyping. The genotyping success rate was ≥ 96%.
Statistical analysis
We used the chi-square test for Hardy-Weinberg equilibrium and statistical analysis to compare allelic and genotypic distributions. Odds ratios (OR) and 95% confidence intervals (95% CI) were calculated according to Woolf’s method19 using the Statcalc program (EpiInfo 2002, Centers for Disease Control and Prevention, Atlanta, GA, USA). P values < 0.05 were considered statistically significant.
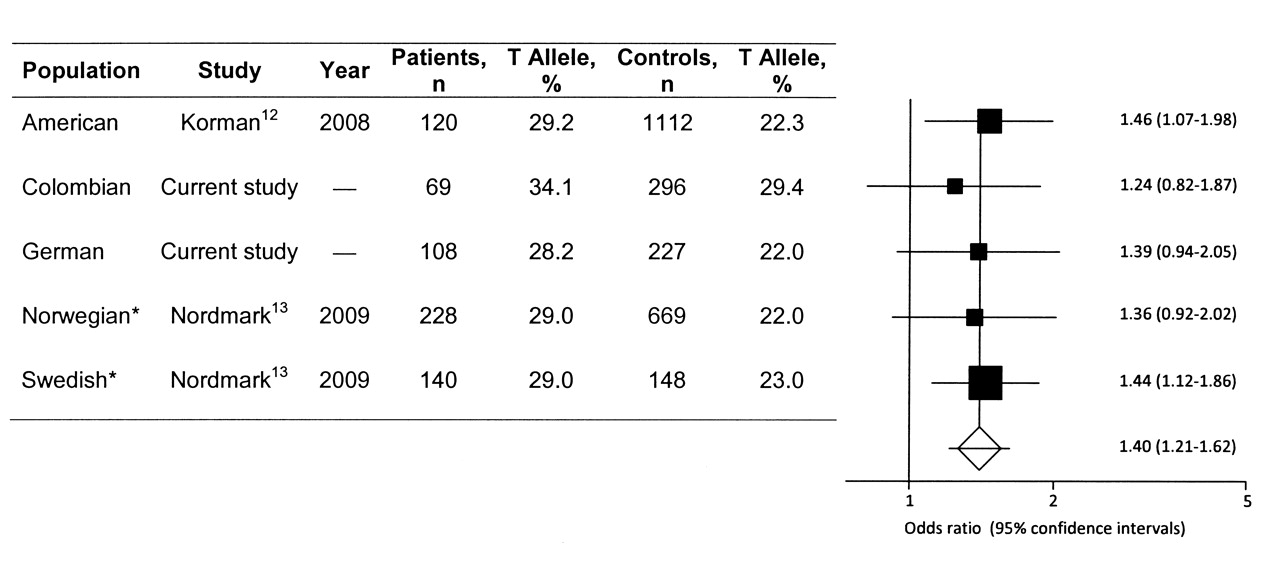
To estimate the common effect size of the T allele on disease, a metaanalysis of previous results together with our own was performed. Characteristics of studies included in the metaanalysis are shown in Figure 1. The metaanalysis was conducted using StatsDirect software (StatsDirect, Cheshire, UK). In all the studies, allele and genotyping frequencies were consistent with Hardy-Weinberg equilibrium in patients and controls. Since the number of studies and the average sample size were small, to calculate the inconsistency of the metaanalysis with a power > 80%, pooled calculations of OR were obtained using random effect models20. Publication bias was investigated by funnel plotting. The significance of the intercept was determined by the t test suggested by Egger.

Pooled data for the STAT4 rs7574865 T allele frequency of North American, Colombian, German, Norwegian, and Swedish populations. OR estimate of each study is marked with a square; the size of the square represents the weight that study exerts in the metaanalysis. Confidence intervals of pooled estimates are shown as a horizontal line. No asymmetry was found, as indicated by the p value of Egger’s t test: bias = −1.46 (95% CI −3.44 to 1.14), p = 0.20. *Data correspond to rs7582694 STAT4 polymorphism that is a perfect proxy of rs7574865.
The power of the study to detect an effect of a polymorphism on disease susceptibility was estimated for an OR of 1.4 (first described effect size), a type I error rate of 0.05, dominant inheritance mode, and 0.005% of population risk. The power was calculated using Quanto version 0.5 software (Department of Preventive Medicine, University of Southern California, Los Angeles, CA, USA).
RESULTS
No deviation from Hardy-Weinberg equilibrium was observed in the study cohorts for the rs7574865 single-nucleotide polymorphism (SNP). Table 1 shows the STAT4 rs7574865 genotype and allele distributions for Colombian and German populations. The T allele increased the risk of developing pSS by 24% in Colombian subjects. The same trend was found in the German cohort, in which the frequencies of the T allele showed a clear trend of association (p = 0.08, OR 1.40, 95% CI 0.96–2.02). Since the observed effect of the T allele was not significant due to low sample size (40% power for the Colombian and 45% for the German cohort), we combined data from previous studies in a metaanalysis in order to improve statistical power (> 99% in the metaanalysis). Interestingly, STAT4 rs7574865 T allele frequencies were quite similar in patients and controls in all the populations, except for the Colombian population, where the T allele frequency in pSS patients was similar to the frequencies in patients with RA and systemic lupus erythematosus (SLE) reported before18. Figure 1 shows results of the metaanalysis of all studies with published allele frequencies for STAT4 rs7574865 in pSS, including the present study. There was no statistically significant evidence for heterogeneity of OR among the studies for this polymorphism in pSS, and the fixed-effects model was chosen (I2 = 0%). Metaanalysis of all 5 cohorts using the Mantel-Haenszel test revealed a highly significant association of this variant with pSS (pooled OR 1.40, 95% CI 1.21–1.62, p = 4.7 × 10−6). No publication bias was found in these studies; the Egger test was not statistically significant (bias = 1.09, 95% CI = −1.35651 to 3.53, p = 0.19), with symmetrical funnel plots (data not shown).
Genotype and allele frequencies of the rs7574865 STAT4 polymorphism in healthy controls and patients with primary SS.
DISCUSSION
Our study adds further evidence indicating an influence of STAT4 rs7574865 T allele in pSS, and confirms STAT4 as one of the most important and replicable genes influencing susceptibility to pSS apart from the HLA region.
The association of rs7574865 variant in the STAT4 gene was initially reported in Korean patients with RA20a, and almost at the same time in Caucasian patients with RA and SLE21. These associations were replicated later in different populations18,22–25 and in autoimmune diseases such as type I diabetes26,27 and systemic sclerosis28. Together, these data suggest that STAT4 could be a useful genetic marker of autoimmunity, and they add further evidence for a common genetic origin of diverse autoimmune diseases29. Nevertheless, there are other autoimmune diseases in which STAT4 rs7465865 showed no influence, such as giant cell arteritis30 and multiple sclerosis26; the reason STAT4 has different effects in different autoimmune diseases remains unknown.
STAT4 genes encode a transcription factor that transmits signals induced by interleukin 12 (IL-12), IL-23, and type 1 interferon31. A major action of IL-12 through STAT4 signaling is to promote the differentiation of naive CD4+ T cells into T-helper 1 (Th1) cells, which produce interferon-γ. These Th1 cells are thought to drive the chronic autoimmune response. STAT4 is also important for the development of IL-17-secreting Th cells in response to IL-2332. As both lineages are master regulators of the etiopathology of pSS in humans33,34, STAT4 may exert its influence through defective signaling in these pathways.
Although it seems clear that STAT4 plays a key role in several pathways in autoimmunity, the functional role(s) of the associated polymorphism(s) remains to be elucidated. The rs7574865 STAT4 variant is located at the third intron of the gene, suggesting a role in STAT4 splicing. Nevertheless, Abelson, et al25 observed no differences in splicing of the gene in peripheral blood mononuclear cells (PBMC). Instead, they reported a correlation between expression levels of STAT4 in PBMC and the risk allele of the STAT4 rs7574865-associated SNP. Additional studies are needed to clarify the role of STAT4 as a novel marker for autoimmune diseases.
Acknowledgments
We thank all the participants in this study.
Footnotes
-
Supported by the Junta de Andalucía, grant CTS-1880, and by Universidad del Rosario, Medellin, Colombia.
- Accepted for publication November 12, 2009.








{kind=link}