
Abstract
Objective. We examined the genetic association of the promoter insertion/deletion (indel) in IRF5 gene with systemic lupus erythematosus (SLE) in distinct populations and assessed its role in gene expression.
Methods. Four IRF5 polymorphisms were genotyped in 1488 SLE patients and 1466 controls. Gene expression was analyzed by quantitative real-time PCR using RNA from peripheral blood mononuclear cells (PBMC).
Results. The promoter indel and rs2070197 had independent genetic effects, which accounted for the association of rs2004640 and rs10954213. Gene expression analysis revealed that rs10954213 exerted the greatest influence on IRF5 transcript levels.
Conclusion. We corroborated the association of the promoter indel with SLE in 5 different populations and revealed that rs10954213 is the main single-nucleotide polymorphism responsible for altered IRF5 expression in PBMC.
- INTERFERON REGULATORY FACTOR 5
- SYSTEMIC LUPUS ERYTHEMATOSUS
- GENE EXPRESSION
The type I interferon pathway bridges the innate and adaptive immune responses and appears to play an important role in the etiology of autoimmune diseases. Interferon regulatory factor 5 (IRF5), one of the genes of the interferon pathway, is strongly associated with many autoimmune diseases, including systemic lupus erythematosus (SLE)1,2, rheumatoid arthritis (RA)3, Sjögren’s syndrome4, inflammatory bowel disease5, and multiple sclerosis (MS)6. Several functional polymorphisms have been described for the gene, including the single-nucleotide polymorphism (SNP) rs10954213 altering poly(A) signal, exon 6 in-frame insertion/deletion, rs2004640 creating a splicing site, and the most recently identified promoter CGGGG insertion/deletion (indel) strengthening the binding site for the Sp1 transcription factor7–10. All the risk alleles of these polymorphisms segregate in a strongly associated haplotype tagged by the C allele of the SNP rs20701978,10.
The aims of our study were to replicate the genetic association of the promoter indel with SLE in a larger set of several independent populations; and to assess the relative contribution of the promoter indel and rs10954213 on gene expression in peripheral blood mononuclear cells (PBMC).
MATERIALS AND METHODS
Patients and controls
The total study sample comprised 1488 patients with SLE and 1466 healthy controls, including 241 cases and 249 controls from Argentina, 178 cases and 265 controls from Mexico, 596 cases and 520 controls from Spain, 263 cases and 247 controls from Italy, and 210 cases and 185 controls from Germany. All patients fulfilled the American College of Rheumatology classification criteria for SLE11,12.
Genotyping
The SNP rs2004640, rs2070197, and rs10954213 were genotyped by TaqMan® SNP allelic discrimination assays (ABI, Foster City, CA, USA). The promoter indel was amplified by polymerase chain reaction (PCR) and analyzed with the ABI Prism 3730×l DNA Analyzer and Gene Mapper software.
Statistical analysis
Genetic association tests and conditional analyses were performed using Unphased version 3.0.1313. Pairwise linkage disequilibrium estimates were calculated with Haploview version 4.114. For expression analysis, statistical calculations were performed using an unpaired 2-tailed t-test as implemented in the GraphPad Software.
Expression analysis
Total RNA was purified from PBMC obtained from 164 healthy Swedish donors and reverse-transcribed as described15. IRF5 expression was determined by quantitative real-time PCR using the forward 5’-GGA GCA TTT TCT CAA TGA GCT CAT C-3’ and reverse 5’-CTG CTA CAG GCA CCA CCT GTA CAG T-3’ primers and SYBR Green for signal detection. We performed initial denaturation at 95°C for 5 min followed by 45 cycles of PCR (95°C for 15 s, 62°C for 10 s, and 72°C for 15 s). PCR buffer was supplemented with 3 mM MgCl2, 200 μM of each of dNTP, primers, SYBR Green, cDNA, and 0.5 U of Platinum Taq polymerase. Expression levels were normalized to the levels of TBP using the comparative 2−ΔΔCt method16. All experiments were run in triplicate. Independent cDNA synthesis was carried out twice.
RESULTS
The genetic analysis revealed a strong association for the insertion allele of the promoter indel in the combined samples (OR 1.60, 95% CI 1.44–1.80, p = 1.5810−19; Table 1). An even stronger association was observed with rs2070197 (OR 2.11, 95% CI 1.81–2.45, p = 2.24 × 10−23), a SNP with unknown function except being reported as a tag of the risk haplotype in previous studies8,10. In order to determine if these 2 markers could account for all the IRF5 association, we performed association tests conditioning on the promoter indel and rs2070197. The results indicated that the indel and rs2070197 had independent effects and no other markers remained associated after conditioning on both of them together (Table 1). Interestingly, the effect size of rs2004640 and rs10954213 as well as their conditional association varied across populations. They were independent of the promoter indel in all populations except Argentina and Spain, which can be explained by the higher linkage disequilibrium (r2) observed between these 2 SNP and the indel in these populations (Table 1). The haplotype formed by the risk alleles of the 4 polymorphisms (iTCA) displayed a very strong association with SLE (OR 2.72, 95% CI 2.19–3.32, p = 3.17 × 10−25; Table 2).
Genetic association and conditional analysis of IRF5 polymorphisms with systemic lupus erythematosus.
Genetic association of IRF5 haplotypes with SLE in the combined samples.
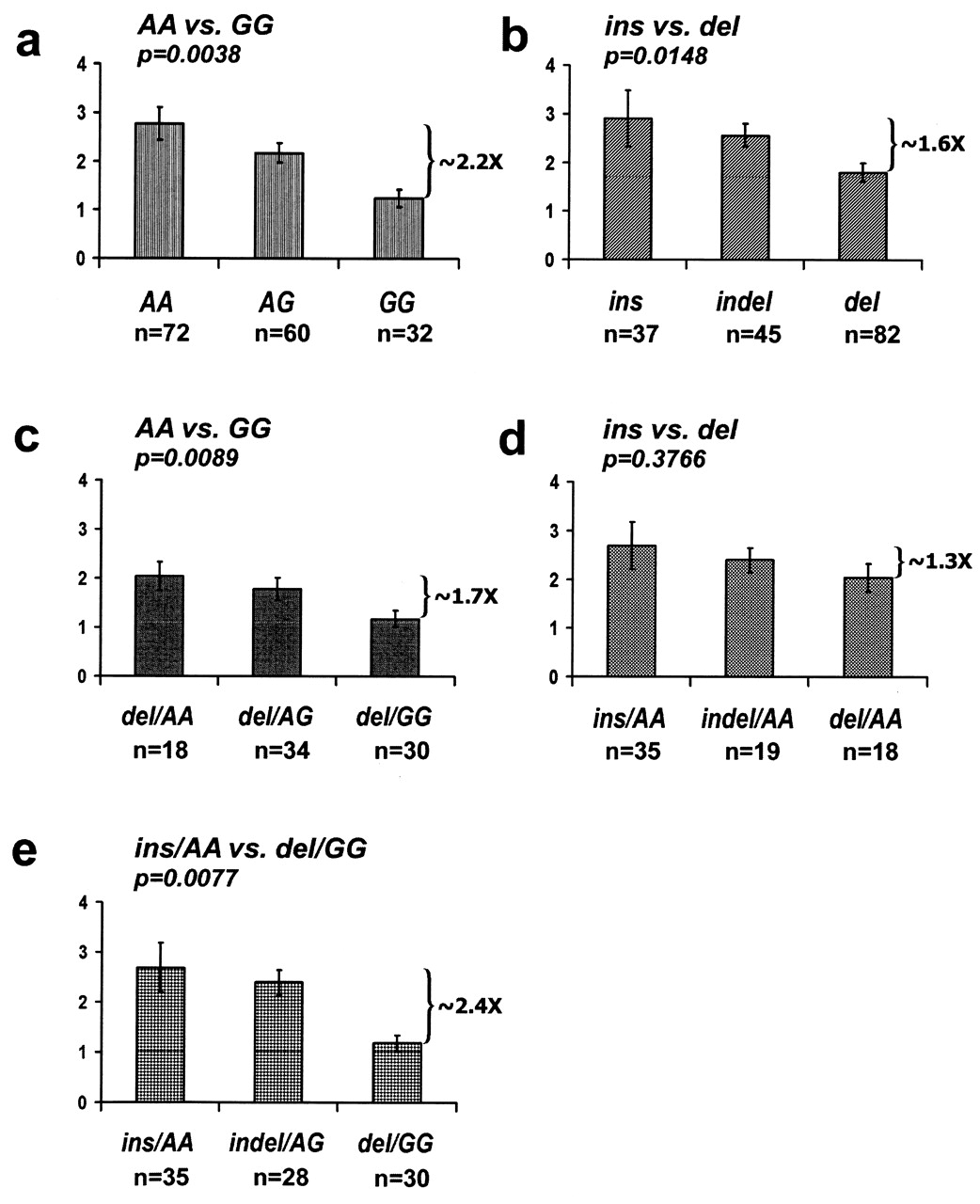
Next, we measured IRF5 gene expression in PBMC. The experiment was performed using healthy volunteers to exclude influence of treatments and/or disease activity on gene expression pattern. The presence of the insertion enhanced IRF5 expression, although to a lesser extent than the risk allele of rs10954213 (Figure 1). When conditioned on rs10954213, the effect of indel became even less pronounced and lost significance (1.3-fold difference between the opposite genotypes; p = 0.3766). While conditioning on the deletion (there were no individuals with insertion/GG), rs10954213 remained strongly associated with the transcript levels (p = 0.0089). Nevertheless, both associated variations exerted an additive effect on gene expression. SNP rs2070197 did not affect the IRF5 transcript levels (data not shown).

{kind=link}
Relative mRNA expression levels of IRF5 in peripheral blood cells. Fold-difference in expression between the opposite homozygous genotypes is shown on the right side of each graph. Data represent mean values of the corresponding number of individuals shown beneath each graph with standard deviation error bars. Samples stratified by the SNP: (A) rs10954213; (B) promoter indel; (C) rs10954213 conditioned on the promoter deletion; (D) promoter indel conditioned on A allele of rs10954213; (E) expression levels for the groups homozygous for both associated alleles (insertion and A-rsl0954213) versus both protective alleles (deletion and G-rs10954213).
DISCUSSION
The importance of IRF5 gene for the development of many autoimmune diseases is now very much appreciated, and genetic associations have been replicated in many populations. In our study, we demonstrated that the promoter indel and rs2070197 have strong independent genetic association with SLE, which seems to account for the association of rs2004640 and rs10954213. But as the eventual aim of any genetic study is to provide an explanation of how associated genetic variants affect gene function, we determined the influence of the promoter indel on gene regulation in comparison with the effect of another functional SNP, rs10954213.
Among many associated variants in the IRF5 gene, 3 are known to have potential functions: promoter indel, exon 6 indel, and 3’UTR poly(A). SNP rs2004640, giving rise to the alternative transcript, seems to play a small role as it encodes essentially the same protein isoform as the transcripts with exon 1A. In addition, the level of this transcript is negligible and does not alter the overall amount of the IRF5 protein10. The exon 6 indel may affect the function of the PEST domain of the protein, but apparently does not play any role in regulating gene expression. Moreover, exon 6 indel has not been shown to be independently associated with SLE8–10. The insertion risk allele has also been found in the protective haplotype in both Europeans and Mexicans, suggesting that its effect on SLE disease might be small17. It was shown independently that only rs10954213 and the promoter indel might affect the levels of the IRF5 gene expression8,9.
It was previously shown in a minigene model that insertion of CGGGG in the proximal promoter of the IRF5 gene increased transcription of the reporter gene9. We measured IRF5 gene expression in PBMC. Surprisingly, the indel showed only a modest, nonsignificant effect, while the poly(A) SNP altered gene expression significantly. Such a minor effect of the indel on gene expression in our investigation compared to the minigene experiments of Sigurdsson, et al9 could be accounted for by a more complex architecture of the promoter region in leukocytes compared to the promoter, in the context of plasmid DNA transfected into unrelated cells, such as the HEK293T used in the minigene experiment9. Our results suggest, however, that both associated variations exert an additive effect on gene expression. SNP rs2070197 located in the 3’UTR and showing an independent genetic effect did not alter the transcript levels and thus its function at present remains unclear.
Our study replicated the association of the promoter indel with SLE in Argentinians, Mexicans, Spaniards, Italians, and Germans, and revealed its relative effect on gene expression in PBMC in comparison to the poly(A) SNP rs10954213.
Acknowledgments
The authors express their gratitude to all patients, healthy volunteers, and clinicians that participated in the research.
Footnotes
-
* Contributed equally to this work.
-
Supported in part by Clas Groschinskys foundation, the Swedish Rheumatism Association, Gurli and Edward Brunnbergs foundation for rheumatology research to SVK, The Swedish Research Council, the Torsten and Ragnar Söderbergs Foundation, the Gustaf Vth 80th-Year Jubilee to MEAR, and by grants SAF2006 and CTS-1180 to JM. Dr. Alarcón-Riquelme is supported by the Knut and Alice Wallenberg Foundation through an award from the Royal Swedish Academy of Sciences. She is also a Greenberg Scholar at the Oklahoma Medical Research Foundation, USA.
- Accepted for publication October 8, 2009.
REFERENCES
- 1.
- 2.
- 3.
- 4.
- 5.
- 6.
- 7.
- 8.
- 9.
- 10.
- 11.
- 12.
- 13.
- 14.
- 15.
- 16.
- 17.