

Abstract
Objective. To investigate in vivo simulation of the microenvironment in which osteoarthritis (OA) chondrocytes are cultured in vitro.
Methods. Human articular chondrocytes were cultured under normoxic and hypoxic conditions. Cells were cultured on standard culture plastic or a porous polyHEMA surface that closely resembles the in vivo cartilage microarchitecture. Morphological changes to the cells were demonstrated by fluorescent staining with DAPI and vinculin. Proteoglycan and type II collagen protein levels were assessed using established techniques. Matrix metalloproteinase-1 (MMP-1) production was assessed by ELISA. The gene expression of type II collagen and SOX9 was measured using real-time polymerase chain reaction.
Results. Cells grown on culture plastic were seen to be flat and hexagonal. Cells cultured on the porous polyHEMA surface exhibited morphology in keeping with the in vivo microenvironment. Glycosaminoglycan release in hypoxia was high from cells cultured on standard culture plastic. Transcriptional expression of type II collagen was upregulated in hypoxia and by culture on the polyHEMA surface. Transcriptional expression of SOX9 in hypoxia was upregulated compared to normoxia; no significant effect was seen by varying the culture surface. Translational expression of type II collagen was upregulated at 20% oxygen on the polyHEMA surface compared to culture plastic and this was related to MMP-1 expression.
Conclusion. Culture of chondrocytes in hypoxia and on a porous surface simulates the in vivo microenvironment and illustrates the molecular mechanisms of OA.
Osteoarthritis (OA) is the most common form of arthritis and is known to affect millions of people worldwide. It is a heterogeneous and multifactorial disease, characterized by the loss of articular cartilage1. Chondrocytes are the only cell type in normal adult articular cartilage; they are known to synthesize all components of the cartilage matrix under normal conditions. The interactions between the chondrocyte and the extracellular matrix (ECM) are important in regulation of the development, maintenance, and repair of the cartilage matrix, which in turn protects the chondrocytes from damage and allows them to maintain their shape and phenotype2,3. The failure of chondrocytes’ ability to maintain the balance between synthesis and degradation of their surrounding matrix is of major clinical importance in the pathogenesis of OA. These chondrocytes resemble terminally differentiated chondrocytes in growth plates and actively produce matrix-degrading enzymes such as the collagenases matrix metalloproteinase-1 (MMP-1) and MMP-134.
A number of in vitro models of OA are currently used. These range from the use of monolayers to the use of explant culture systems, many of which are carried out under normoxic conditions. The culture of chondrocytes in monolayer under normoxic conditions is particularly problematic, with one key issue being the early loss of phenotype from chondrocytic to fibroblastic5. The phenotype of chondrocytes is characterized by expression of specific genes such as type II collagen (COL2A1) and transcription factor SOX96. Type II collagen is known to be abundant in articular cartilage7, while SOX9 is highly expressed and is thought to control a number of genes in articular cartilage. The expression of SOX9 is known to rapidly decline in chondrocytes in monolayer cultures and is coupled with a reduction in type II collagen8–10. However, a recent study has shown that hypoxia has an important role in maintaining the chondrocytic phenotype11. The growth of chondrocytes in monolayer can be considered to be an alien environment for the cells, as in vivo chondrocytes are typically found in a 3-dimensional structural matrix environment. Therefore the use of different surfaces and scaffolds in the culture of chondrocytes that more closely simulate the in vivo microenvironment has an important effect on cell behavior12.
Our objective was to investigate in vivo simulation of the microenvironment in which OA chondrocytes are cultured.
MATERIALS AND METHODS
All chemicals and biochemicals were commercial analytical grade reagents obtained from Fisher (Loughborough, UK), BDH (Poole, UK), Abcam plc (Cambridge UK), and Applied Biosystems (Warrington, UK).
Porous polymer surface preparation
The powder form of Poly [2-hydroxy-ethyl methylacrylate] (polyHEMA) was obtained from Sigma (Poole, UK). A stock solution of polyHEMA 120 mg/ml was dissolved in 95% ethanol. The stock solution was further diluted with 95% ethanol to 1 mg/ml for coating 12-well tissue culture plates. Each well was coated with 1 ml of polyHEMA solution. Plates were left to dry overnight in a tissue culture hood. The porous polyHEMA surfaces were checked the next day for uniformity by light-phase microscopy and the wells washed with phosphate buffered saline (PBS) twice before cell culture.
Cell culture
Human articular chondrocytes (HAC) were derived from articular cartilage obtained from consenting patients with OA following knee arthroplasty. Enzymatic digestion of tissue and culture of cells was performed as described13. Cells were cultured to 80% confluence over a period of 7 days in T75 cm2 flasks in normoxia. Cells were passaged once and seeded at 12.5 × 104 cells per well onto the 12-well plates, then cultured in either 20% or 1% oxygen on culture plastic and on the polymer surface for a further 4 days. Day 4 was taken as the experimental endpoint to avoid cells reaching 100% confluence and therefore behaving as they would in monolayer. Cells were harvested for the various experiments as described below. Experiments were repeated in quadruplicate from a total of 4 different samples from patients with OA.
Hypoxic cell culture
The Sanyo Trigas incubator was used to culture cells at 1% oxygen.
Proteoglycan assay
Media samples were assayed for sulfated glycosaminoglycans (GAG) as a measure of proteoglycan release at Day 4. A range of GAG standards (bovine chondroitin sulfate) from 0–40 mg/ml were prepared in phosphate buffer. Supernatants (n = 4) were diluted in phosphate buffer and plated alongside the standards at 40 μl in duplicate into a 96-well plate; 250 μl of 1, 9-dimethylmethylene blue (DMB) was added and the absorbance read immediately at 530 nm.
Western blotting
Cytoplasmic samples were extracted from cells as follows: cells were lysed with ice-cold buffer [50 mM Tris-Cl, pH 7.5, 1.2 M glycerol, 1 mM EGTA, 1 mM EDTA, 1 mM Na3VO4, 10 mM 2-glycerophosphate, 50 mM NaF, 5 mM sodium pyrophosphate, 1% (v/v) 2-mercaptoethanol, with protease inhibitor (Roche Applied Sciences)]. Particulate matter was removed by centrifugation at 13,000 g, 5 min at 4°C, and lysates were stored at –80°C until use. Bradford protein analysis was performed to allow equalization of protein content before running the blot. Heat-denatured samples were run on 10% gels followed by transfer to nitrocellulose membranes (Protran®; Whatman plc, Brentford, UK). Blots were probed with the collagen type II primary antibody (Abcam, Cambridge, UK) and horseradish peroxidase-conjugated secondary antibody (Dako UK Ltd., Cambridgeshire, UK). This was followed by detection of immunoreactive bands by enhanced chemiluminescence (GE Healthcare UK Limited).
Matrix metalloproteinase-1 (MMP-1) ELISA
A specific MMP-1 ELISA was utilized to assess expression of MMP-1 in the culture supernatant12. This assay was performed at Day 4.
RNA extraction
RNA was purified from the cells using a Qiagen RNeasy mini kit (Qiagen Ltd., West Sussex, UK) according to the manufacturer’s instructions.
Reverse transcription
The RNA was reverse-transcribed into cDNA using the Superscript™ II kit. Random hexamers [pd(N)6] (1 μg/μl) was added to 9 μl of total RNA with a concentration of 1 μg and annealed at 70°C for 10 min before being chilled on ice. A quantity of 5× first-strand buffer (4 μl), 2 μl DTT, 1 μl of 10 mM dNTPs, and 1 μl Superscript reverse transcriptase were added. To minimize variations in efficiency, all samples from a single experiment were reverse-transcribed simultaneously.
Real-time polymerase chain reaction (PCR)
Ready-designed primers for type II collagen (ID: Hs00156568 m1) and SOX9 (ID: Hs00165814 m1) were obtained (Applied Biosystems). Gene expression was measured by TaqMan real-time PCR (7900HT system; Applied Biosystems)14. Cycling conditions for Taqman PCR (Jumpstart Taq Readymix; Sigma) were 2 min at 50°C, 10 min at 95°C, then 40 cycles of 15 s at 95°C and 1 min at 60°C. Quantification of the results was achieved by production of a standard curve using a cDNA sample known to express the type II collagen and SOX9 genes, with a top standard of 2.5 μg of cDNA. Lower standards were produced by carrying out doubling dilutions of the cDNA. Correction was obtained using ribosomal 18S RNA levels. Results were analyzed using the Sequence Detection System 2.2 software program (Applied Biosystems).
Fluorescent staining
4',6-diamidino-2-phenylindole (DAPI) nuclear stain and vinculin cytoskeleton stain were used (Sigma). Media was removed and the cells washed with PBS. 500 μl of 4% paraformaldehyde was added to each well for 10 min. Coverslips were washed with PBS and removed from the wells for staining. 400 μl of PBS containing DAPI and vinculin was added to each coverslip. The coverslips were incubated in the dark for 45 min and then wet-mounted for microscopy. These experiments were performed at Day 2 in order to show the chondrocyte morphology clearly.
Statistical analysis
Statistical analysis was by comparison of the means using one-way ANOVA with Bonferroni post-hoc test. SPSS v15 software was used.
RESULTS
Effect of altered culture surface on chondrocyte morphology
Human articular chondrocytes (HAC) were cultured on either glass coverslips or on coverslips coated with the porous polyHEMA surface. The cells were stained with vinculin to aid visualization of altered cell morphology. Cell morphology on the porous polyHEMA surface differed markedly from cells on standard culture plastic. The cells grown on plain coverslips were seen to be flat against the surface and spread out in a polygonal shape (Figure 1A). The cells on the polyHEMA surface inserted pseudopods through pores onto the coverslip below, while maintaining a rounded shape (Figure 1B).

The effect of altered culture surface on chondrocyte morphology. Cells grown on plain coverslips were flat against the surface and spread broadly in a polygonal shape (A), while cells on the polyHEMA surface were rounded and utilized pseudopods inserted through pores to anchor to the glass below (B). DAPI and vinculin stain.
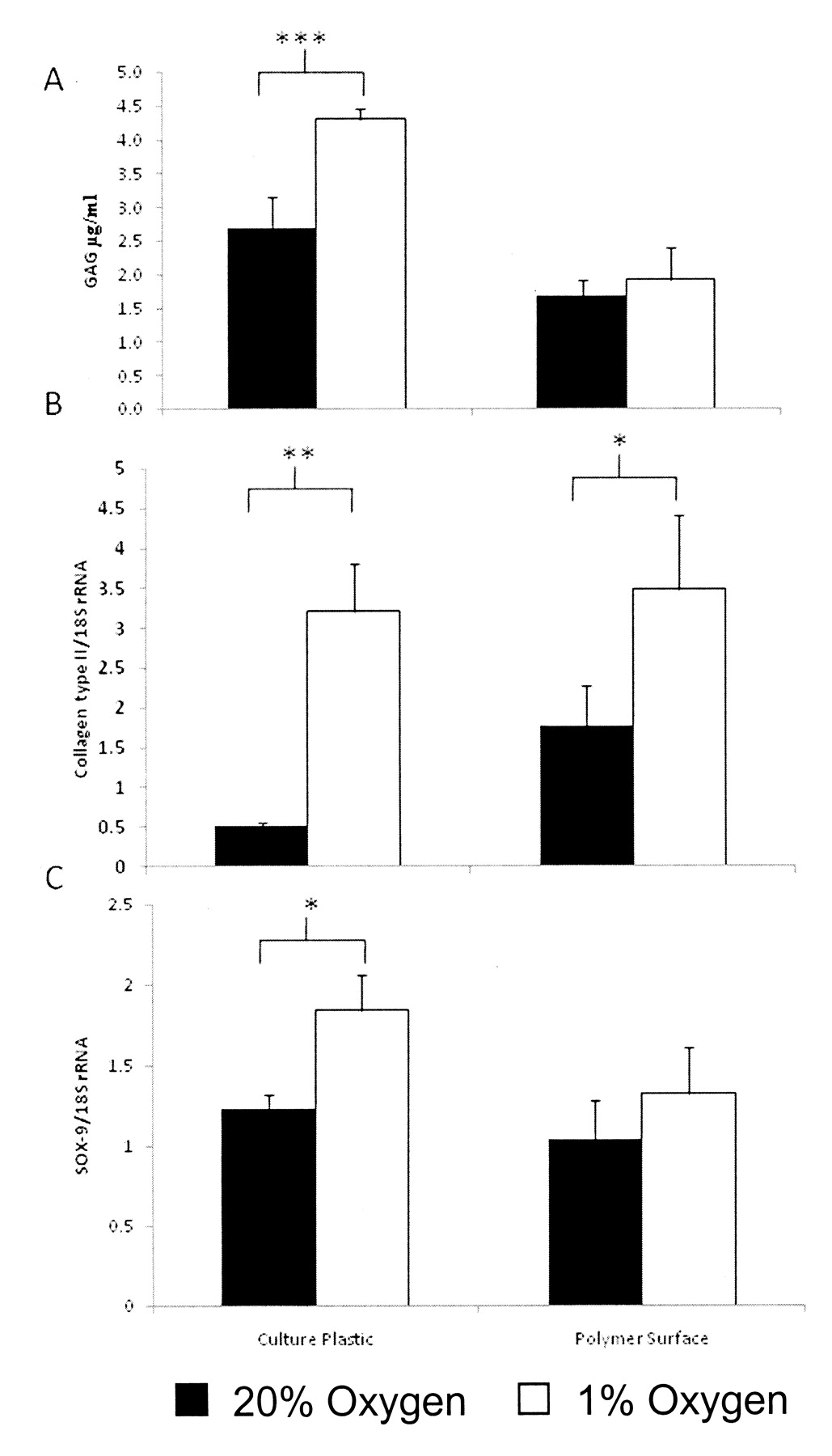
Effect of altered atmospheric conditions and culture surface on proteoglycan discharge into the culture supernatant
HAC were cultured on either culture plastic or on the porous polyHEMA surface. Cells on each surface were cultured for 4 days at either 1% oxygen or normal atmospheric conditions (20% oxygen). Media was harvested from the cells and assayed to measure proteoglycan release. Cells grown on normal tissue culture plastic showed a variation in proteoglycan release, with cells cultured at 1% oxygen releasing significantly more proteoglycan than the cells cultured at 20% oxygen, 4.3 ± 0.15 μg/ml and 2.7 ± 0.45 μg/ml, respectively (Figure 2A). However, cells grown on the polyHEMA surface showed a different effect; there was no significant difference, with cells cultured at 20% oxygen releasing 1.7 ± 0.24 μg/ml and cells cultured at 1% oxygen releasing 1.9 ± 0.47 μg/ml.

The effect of altered oxygen tension and culture surfaces on glycosaminoglycan (GAG) release and type II collagen and SOX9 gene transcription in human articular chondrocytes (HAC). HAC were cultured in 1% or 20% oxygen on culture plastic or a porous polyHEMA surface. Release of GAG by cells into culture medium was assayed as described in Materials and Methods. Cells on normal culture plastic at low oxygen tension showed a significant increase in GAG release (μg/ml) compared to cells grown on the same surface in normoxia (A). cDNA was analyzed for type II collagen expression by real-time PCR (B); HAC cultured on the porous polymer surface in 1% oxygen had the highest expression of type II collagen compared to cells on culture plastic in 20% oxygen. cDNA was also analyzed for expression of SOX9 by real-time PCR (C); HAC cultured in hypoxia showed increased expression of SOX9, regardless of the surface, compared to cells in 20% oxygen. Results expressed as mean ± SE. *p < 0.05, **p < 0.005, ***p < 0.001.
Effect of altered oxygen tension and culture surface on type II collagen and SOX-9 gene expression in human articular chondrocytes
cDNA was obtained from RNA purified from HAC cultured on either culture plastic or the polyHEMA surface. In addition, these cells were cultured at varying oxygen tensions (1% and 20% oxygen). The expression of type II collagen was seen to differ markedly depending upon the oxygen tension and also the culture surface. On culture plastic the expression of type II collagen was greatly improved by reducing the oxygen levels to 1% from 20%, with expression of 3.2 ± 0.6 ng/μl cDNA and 0.5 ± 0.1 ng/μl cDNA, respectively (Figure 2B). Type II collagen expression was also improved by culturing the cells on the polyHEMA surface in contrast to culture plastic at 20% oxygen, with expression of 1.8 ± 0.5 ng/μl cDNA. The culture of HAC at 1% oxygen on a polyHEMA surface gave the greatest expression of type II collagen, at 3.5 ± 0.9 ng/μl cDNA.
Results for SOX9 showed that, in normoxia, SOX9 expression was not affected by the variation in culture surface, with expression of 1.2 ± 0.1 ng/μl cDNA on culture plastic and 1.0 ± 0.2 ng/μl cDNA on the polyHEMA surface (Figure 2C). The cells cultured in 1% oxygen showed a slight increase in SOX9 expression; again there was no significant difference between the surfaces, with expression of 1.8 ± 0.2 ng/μl cDNA on the culture plastic and 1.3 ± 0.3 ng/μl cDNA on the polyHEMA surface. The only significant difference in SOX9 expression was the increase from cells on culture plastic in hypoxia (1.8 ± 0.2 ng/μl cDNA) compared to their counterparts in normoxia (1.2 ± 0.1 ng/μl cDNA).
Effect of altered atmospheric conditions and culture surface on type II collagen and MMP-1 production in HAC
Cytoplasmic fractions were extracted from HAC that had been cultured on culture plastic or the polyHEMA surface and at either 1% or 20% oxygen. The Bradford assay was used to normalize the fractions before Western blotting with probes for type II collagen (Figure 3A). The results showed a greater amount of type II collagen present in the fractions from cells grown at 20% oxygen on the polyHEMA surface than those grown at the same oxygen tension on normal culture plastic. In contrast, the presence of type II collagen was greater at 1% oxygen on culture plastic than on the polyHEMA surface.

The effect of altered oxygen tension and culture surfaces on the release of MMP-1 and translational type II collagen from human articular chondrocytes (HAC). HAC were cultured at 1% oxygen or 20% oxygen on culture plastic or the porous polyHEMA surface. Cytoplasmic fractions were extracted from HAC and analyzed by Western blot for type II collagen (A). At 20% oxygen, cells cultured on the polyHEMA surface increased their production of type II collagen. However, cells grown at 1% oxygen on the polyHEMA surface produced less type II collagen than the cells grown on culture plastic at 1% oxygen. Culture medium was assayed for MMP-1 production (B); HAC cultured on the porous polymer surface in 1% oxygen produced significantly more MMP-1 compared to their counterparts on culture plastic in 1% oxygen. Results expressed as mean ± SE. *p < 0.05, **p < 0.005, ***p < 0.001.
Culture medium was also assayed for MMP-1 production by ELISA. The chondrocytes grown on culture plastic showed that in normoxia the production of MMP-1 of 55.6 ± 2.6 ng/ml was more than that in cells cultured in hypoxia, with MMP-1 production of 47.7 ± 2.5 ng/ml (Figure 3B). In contrast, cells grown on the polyHEMA surface at 1% oxygen showed increased MMP-1 production, 67.0 ± 5.1 ng/ml, compared to cells cultured at 20% oxygen, 44.3 ± 1.0 ng/ml. The increase in MMP-1 from cells cultured on the porous polymer surface at 1% oxygen was significantly greater than MMP-1 production by cells grown on culture plastic at 1% oxygen.
DISCUSSION
Interactions that occur between chondrocytes and their extracellular matrix are highly important to regulate the development, maintenance, and repair of the matrix. This in turn protects the chondrocytes from damage, allowing them to maintain their shape and phenotype2,3. Chondrocyte metabolism operates under low oxygen conditions within the cartilage matrix, where oxygen levels range from below 1% at the articular surface to 10% in the subchondral zone1. During physiological hypoxia, hypoxia-inducible factor-1α (HIF-lα) acts as a survival mechanism for the chondrocyte, giving it the capacity to react to environmental change1. Chondrocytes are surrounded by a narrow pericellular layer in which high concentrations of sulfated proteoglycans can be found. Once cartilage matrix is formed within the adult, chondrocytes are able to maintain a low rate of turnover of these matrix proteins; however, there are regional differences in the remodeling of chondrocytes, with matrix turnover being faster in the immediate pericellular zones1. The pathogenesis of OA involves a breakdown of this regulation, with MMP-1 and MMP-13 the key drivers of matrix destruction2.
We investigated the effect of culturing chondrocytes under various conditions. Chondrocytes were cultured under both normoxic and hypoxic conditions in order to study the effects of changes in atmospheric conditions. The microarchitectural effects on HAC culture were also assessed using our novel porous polyHEMA surface. The combination of hypoxia and the microarchitechural simulation of the in vivo chondrocyte setting provided a robust and novel in vitro model of OA. Our results showed a number of interesting factors. The morphology of the cells on the porous polyHEMA surface was particularly interesting; we believe this surface allows a closer simulation of the in vivo HAC microarchitectural environment. The morphology of the cells in our model closely resembled that of chondrocytes following histological in situ staining of OA cartilage matrix15. The cells on the porous surface exhibited fewer specific vinculin-positive focal contacts with the ECM and more diffuse vinculin staining throughout the cytoplasm. This provides further evidence of the altered balance of cell:ECM interactions in cells on the polyHEMA surfaces, leading to apparently more abundant cytoplasmic localization of vinculin as less is recruited to integrin complexes at the cell periphery. Culture on this surface alters cell to cell and cell to ECM interactions, and the combination of this factor and hypoxia produced behavioral changes in the HAC, as indicated by the significant rise in type II collagen gene expression. That the same effect was not seen at a protein level with Western blotting could be explained by the high level of production of MMP-1 seen under the same conditions. The lack of type II collagen in the cytoplasmic fractions may be a cellular response by the chondrocyte: it is possible that the previous production of MMP-1 by the cell has caused the cell to release collagen into its environment as it attempts to maintain the balance between synthesis and degradation that occurs in the in vivo microenvironment. Investigation including assays such as radioactive tritiated collagenase activity would be needed to confirm this. Another possibility is that it may take longer to achieve the translational effect as evidenced by a study of cultures of porcine chondrocytes on polyHEMA. That study showed that the chondrocytes expressed the largest amounts of type II collagen at 6 weeks of continuous culture in suspension16. The lack of effect seen with GAG release from the cells into the media under the conditions of our model suggests the chondrocytes were reacting as they would in their normal in vivo environment: it is known that GAG turnover is decreased in the low oxygen tensions seen in normal articular cartilage2. Moreover, in our model it is likely that, rather than being discharged into the supernatant, the GAG was being incorporated into the matrix being formed around the porous polyHEMA as the chondrocytes anchored themselves to the surface.
While the expression levels of SOX9 did not change on different surfaces, the increase seen in 1% oxygen compared to 20% oxygen suggests that hypoxia enhances the ability of chondrocytes to maintain their phenotype in vitro. We believe that SOX9 regulation is oxygen-dependent17, but not necessarily subject to changes in cell morphology. In addition, a study has shown that HAC cultured in vitro from OA tissue initially express high levels of SOX9 compared to HAC from healthy tissue. This expression diminished significantly when the cells from OA tissue were “re-differentiated” by further culture in millicell filters coated with type II collagen18. The increased expression of MMP-1 was another interesting finding in this setting. These data suggest that hypoxia may influence MMP-1 expression and therefore could be an important feature in the progression of OA.
Our study indicates that the current widely used model of chondrocyte culture in normoxia and in monolayer requires refinement to more closely simulate the in vivo microenvironment. This will allow understanding of the molecular mechanisms of OA and pursuit of novel potential therapeutic targets.
Footnotes
-
Supported by the Royal College of Surgeons of England, the Shears Foundation, Rosetrees Trust, Arthritis Research Campaign, and the Dunhill Medical Trust.
- Accepted for publication August 6, 2009.








{kind=link}
{kind=link}
{kind=link}