

Abstract
Objective. Signaling lymphocytic activation molecule (SLAM) has been related to the pathology of systemic lupus erythematosus (SLE) through regulation of T cell-dependent humoral immune responses. We investigated the functional associations of the −262A/T and −188A/G polymorphisms of SLAM in Chinese patients with SLE.
Methods. Genotyping of −262A/T (rs2295614) and −188A/G (rs2295613) in SLAM was carried out in 248 cases and 278 controls. Promoter activities of haplotypes on the SLAM gene were evaluated with the dual-luciferase reporter system. The mRNA expressions of SLAM on peripheral blood mononuclear cells (PBMC) of SLE patients with different genotypes were determined by real-time polymerase chain reaction.
Results. Frequencies of −262A allele and −188G allele were significantly higher in SLE patients than in controls. Haplotype analysis and multifactorial logistic regression analysis showed that individuals with the AG/AG haplotype had increased susceptibility to SLE (p = 0.002, OR 1.478, 95% CI 1.152–1.897). In response to PHA stimulation, the SLAM mRNA expression on PBMC of SLE patients was significantly higher in −262A-188G haplotype homozygotes compared with −262A-188G heterozygotes and individuals with other genotypes.
Conclusion. Our findings suggest that −262A-188G haplotype in the SLAM gene promoter contributes to the risk of SLE by increasing the expression of SLAM.
Systemic lupus erythematosus (SLE) is a prototypic systemic autoimmune disease characterized by production of autoantibody, activation of complement, and deposition of immune complex1. SLE has long been appreciated to be a complex interplay of genetic and environmental factors. The complex patterns of inheritance in SLE suggest that multiple genes contribute to the etiology2. Recently, relevant genes and convincing genetic effects have been identified. IRF53, ITGAM4, STAT45, PDCD16, and PTPN227 have all been shown to have genetic associations with lupus.
The abnormal response to stimulation of T cells from patients with SLE exhibits a notable array of defects that probably contribute to the origin and development of the disease8. Signaling lymphocytic activation molecule (SLAM) is a member of the CD2 immunoglobulin super-family9. SLAM demonstrates homotypic binding and functions as a costimulatory molecule10. Enhanced expression of SLAM on T cells from the synovial fluid and synovial tissue of patients with rheumatoid arthritis (RA) has been reported. Synovial fluid mononuclear cells stimulated with anti-SLAM antibodies exhibited increased production of interleukin 10 (IL-10), interferon-γ, and tumor necrosis factor-α, indicating that homotypic SLAM-SLAM interactions between T cells and synovial mononuclear cells could contribute to the disease process11. A series of further genetic mapping studies in murine lupus identified a region on chromosome 1 that contains the Sle1b locus. This region contains the entire SLAM family gene cluster, and subsequently 12 nonsynonymous single-nucleotide polymorphisms (SNP) have been identified. Susceptibility to autoimmunity in B6.Sle1b mice is associated with extensive polymorphisms between 2 divergent haplotypes of the SLAM/CD2 family of genes. The autoimmunity may be mediated by a combination of genes in the SLAM/CD2 family cluster12.
We investigated SNP in the SLAM from HapMap and selected 2 high frequency polymorphisms, rs2295613 and rs2295614, located in the SLAM promoter. We explored the associations between the 2 SNP and SLE in Chinese patients, and then clarified the functional significance of the polymorphisms.
MATERIALS AND METHODS
Subjects
The study included 248 Han Chinese patients with SLE (235 women, mean age 34.28 ± 12.46 yrs; 13 men, mean age 31.54 ± 14.70 yrs) recruited consecutively from Southwest Hospital, Xinqiao Hospital, and Daping Hospital in Chongqing and the First Affiliated Hospital of Chongqing Medical College. Diagnosis was established based on the 1987 American College of Rheumatology criteria for SLE13,14. Then 278 ethnically matched healthy subjects (264 women, mean age 36.05 ± 10.81 yrs; 14 men, mean age 39.71 ± 11.44 yrs) matched for age and sex with patients were included as controls. Samples were obtained from subjects after they provided written informed consent. The study was approved by the ethics committees of Southwest Hospital. Clinical and laboratory manifestations of these patients have been summarized15.
SLAM genotyping
Genomic DNA from controls and cases was isolated from 5-ml blood samples. Two SLAM SNP were studied, −262A/T (rs2295614) and −188A/G (rs2295613) located in the promoter region. SLAM genotypes were determined by polymerase chain reaction-restriction fragment length polymorphism (PCR-RFLP) assay.
For −262A/T, an amplification using forward primer 5′-TCA TCG GGG TCA CAG GGT TTC TTC-3′ and reverse primer 5′-TAA AAA TAT CTG AGC AGT TCT CTC CGG TGA G-3′ was performed; a mismatch in the primer created a restriction site for Hinf1. Digested amplicons showed 262 bp, 230 bp, and 32 bp bands for heterozygotes. The primers for amplifying DNA containing the −188A/G site were 5′-TGA AAT AAA ACA GAA GTA AGA GG-3′ and 5′-GAG TGA CCA AAC ACA ATA CCC AG-3′. Digestion of the PCR product with Hin6I produced 763 bp, 567 bp, and 196 bp for heterozygotes. For every genotype, we took 3 samples separately as the templates and conducted PCR; PCR products were analyzed to confirm the results of genotyping by DNA sequencing.
Dual-luciferase reporter analysis
Two representative samples for 2 common haplotypes (−262A-188G, −262T-188A) were selected, and the fragment from −415 to +4 containing SNP −262A/T, −188A/G was amplified. The primers for amplification of the fragment were 5′-GAG GTA CCA GAC TGC ATG TTG AGG TGT-3′ and 5′-CAG AGC TCA ACC CGC TTC CTG TAT CAC TG-3′, which contained 5′-KpnI and 3′-SacI cloning sites, respectively. PCR product was digested with KpnI and SacI and ligated into an appropriately digested pGL3-Basic vector. The resulting construct was designated Basic-AG because sequence analysis showed it contained an A at −262 and G at −188 polymorphic sites. Basic-TA was similarly denoted. Both constructs used in this study were sequenced to confirm their authenticity. The 2 representative samples used in the dual-luciferase reporter constructs differed only at the SNP loci −262 and −188.
The human Jurkat T cell lymphoma cell line was grown in RPMI-1640 medium supplemented with 10% fetal calf serum in an atmosphere of 5% CO2. HeLa cells were maintained in Dulbecco modified Eagle’s medium supplemented with 10% fetal bovine serum at 37°C with 5% CO2. Cells were plated without antibiotic 1 day prior to transfection in 24-well plates to achieve 90% confluence on the day of transfection.
Transfection was carried out using Lipofectamine Reagent (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s protocol. Cells were cotransfected with 0.073 μg reporter plasmid and 0.73 g of pRL-TK; the latter was used to standardize transfection efficiency. After transient transfection for 48 h, cells were washed in paraformaldehyde in phosphate-buffered saline and harvested by addition of 100 ml passive lysis buffer. The activities of firefly luciferase and renilla luciferase were measured sequentially on a GloMax™ 20/20n Luminometer (Turner BioSystems, Sunnyvale, CA, USA) using Dual-Luciferase Reporter Assay reagents (Promega, Madison, WI, USA). The ratio of firefly luciferase level to renilla luciferase level was used to determine SLAM promoter activity and was expressed as relative luciferase activity. For each plasmid construct, 4 independent transfection experiments were performed, each repeated 4 times.
mRNA expression of SLAM in SLE patients
To determine the accumulation of the PCR product, SYBR Green I dye (Toyobo, Tokyo, Japan) was used as a real-time reporter of the presence of double-stranded DNA. Forty SLE patients were enrolled: 5 with homozygotes −262AA-188GG, 19 with heterozygotes −262A-188G, and 16 with other genotypes. Peripheral blood mononuclear cells (PBMC) from patients with different genotypes were stimulated with phytohemagglutinin (100 ng/ml) for 48 h. Total RNA was isolated from peripheral blood of SLE patients with different genotypes using Trizol reagent (Invitrogen). An aliquot of total RNA (0.5 μg) from each specimen was reverse transcribed into single-strand cDNA using oligodeoxythymidylic acid primer. Each single-strand cDNA was diluted for subsequent PCR amplification of SLAM and ß-actin, the latter used as an internal quantitative control. Primers used for amplification of SLAM were 5′-ACA CCA GAG ACC AAC AAA GGG ACT-3′ and 5′-CTG CTA CAA CAC AAA GAT GGA ACG-3′, which produced a 270-bp fragment. For ß-actin, the primers were 5′-CGT GGA CAT CCG CAA AGA C-3′ and 5′-CTC GCT CCA ACC GAC TGC T-3′, which generated a 455-bp fragment. Real-time PCR was carried out using SYBR Green Real-time PCR Master Mix.
PCR was carried out under the following conditions: an initial denaturing step of 5 min at 95°C, followed by 40 cycles of 20 s at 94°C, 45 s at 57°C, and 30 s at 72°C, and a final elongation step of 5 min at 72°C.
PCR products of SLAM and ß-actin were separately ligated into the pGEM T vector. Standard curves for SLAM and ß-actin were constructed using serial dilutions of PCR product-inserted plasmid DNA standards (1 × 101 to 1 × 105 copies/μl). After complete amplification, a dissociation curve was generated and analyzed for each sample. Mean cycle threshold (Ct) values of the test samples were used for statistical analysis. Relative expression of SLAM mRNA was normalized as the ratio of mRNA of SLAM to ß-actin.
Statistical analysis
Hardy-Weinberg equilibrium and distribution of SLAM genotypes and haplotypes of SLE patients and controls were analyzed by chi-square test. The linkage disequilibrium analyzer16 was used to measure pairwise linkage disequilibrium. Haplotype frequencies were estimated and calculated using the Phase package17. Odds ratios and 95% confidence intervals were calculated by unconditional logistic regression as a measure of association with risk for development of SLE. All OR were adjusted for age and sex.
Statistical analysis for luciferase differences and mRNA levels were determined by t test, and p < 0.05 was considered significant. Data are shown as mean and SD. Statistical analyses were performed with SPSS (version 13.0).
RESULTS
Genotypes and risk of SLE
The genotyping results are shown in Table 1. The distributions of allelic frequencies among cases and controls were in Hardy-Weinberg equilibrium (p > 0.05). Statistical analysis of allelic frequencies demonstrated that the frequency of SLAM −262A allele was overrepresented in cases (66.5%) compared with controls (55.2%; p = 0.0003), and frequency of SLAM −188G allele was overrepresented in cases (64.2%) compared with controls (54.6%; p = 0.002). Linkage disequilibrium was calculated and the results showed that these polymorphisms were under the linkage disequilibrium (r2 = 0.4042, D’ = 0.6443, d = 0.6376). We constructed the haplotypes with the 2 SNP using Phase software. Four haplotypes were detected; the frequency of the −262A-188G haplotype was significantly high in patients with SLE (p = 0.001; Table 2). Unconditional logistic regression analysis adjusted by inter-ferential factors revealed a significant association of haplotype −262A-188G with SLE (p = 0.002, OR 1.478, 95% CI 1.152–1.897); results are summarized in Table 3. Because −262A-188G and −262T-188A took 84.3% of all the haplotypes in the population, we studied the functional significance of these 2 haplotypes.
Genotype and allele frequencies of SLAM among cases and controls.
Distribution of estimated haplotype frequencies for SLAM gene in cases and controls.
Association risk analysis for SLE by the haplotypes constituted with polymorphisms of SLAM gene.
Effects of SLAM −262A-188G haplotype on transcriptional activity
To directly determine the allele-specific effects of SLAM promoter activity, we constructed 2 luciferase reporter gene constructs by PCR spanning −415 to +4 of the SLAM promoter region containing 2 experimentally detected SNP haplotypes, −262A-188G and −262T-188A, and they were used to transfect transiently into HeLa cells and Jurkat cells. The promoter activities were expressed as the ratio between firefly luciferase and renilla luciferase activity. Fold-increase was measured by defining the activity of the empty pGL-3 Basic vector as 1.
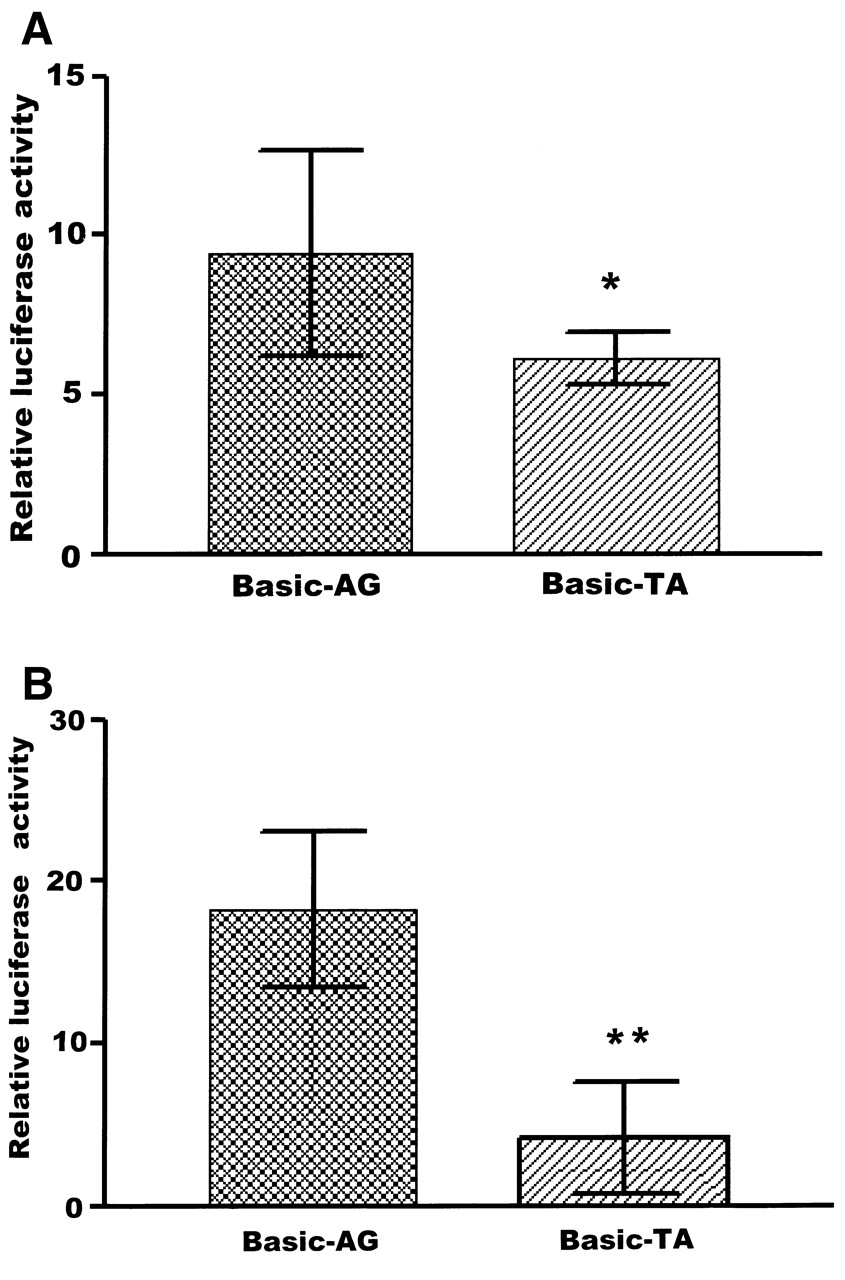
As shown in Figure 1A, the reporter gene expression driven by Basic-AG was significantly greater than that of Basic-TA in Hela cells (9.42 ± 3.24 vs 6.13 ± 0.81 relative luciferase activity, respectively; p < 0.05). Similarly, in Jurkat cells, reporter gene expression driven by Basic-AG was significantly greater than that of Basic-TA allele (18.29 ± 4.80 vs 4.19 ± 3.46; p < 0.01; Figure 1B).

Transient reporter gene expression assays; results are expressed as mean ± SD. A. Induction of luciferase expression of the 2 constructs in Hela cells. The reporter gene expression driven by the Basic-AG was significantly greater than that for Basic-TA, 9.42 ± 3.24 vs 6.13 ± 0.81 relative luciferase activity, respectively. *p < 0.05 compared with Basic-AG. B. Luciferase expression of the 2 constructs in Jurkat cells: reporter gene expression driven by Basic-AG was significantly greater than that for Basic-TA allele, 18.29 ± 4.80 vs 4.19 ± 3.46, respectively. **p < 0.01 compared with Basic-AG.
To verify whether the −262A/T and −188A/G SNP could influence the endogenetic expression of SLAM, we examined mRNA levels in PBMC of 40 SLE patients with different genotypes by quantitative RT-PCR. The results are shown in Figure 2. The mean SLAM mRNA expression levels of 5 samples with −262AA/-188GG genotypes were significantly higher than those of other samples with −262A-188G heterozygotes (0.037 ± 0.002 vs 0.025 ± 0.004; p < 0.01) and other genotypes (0.037 ± 0.002 vs 0.024 ± 0.004; p < 0.01).
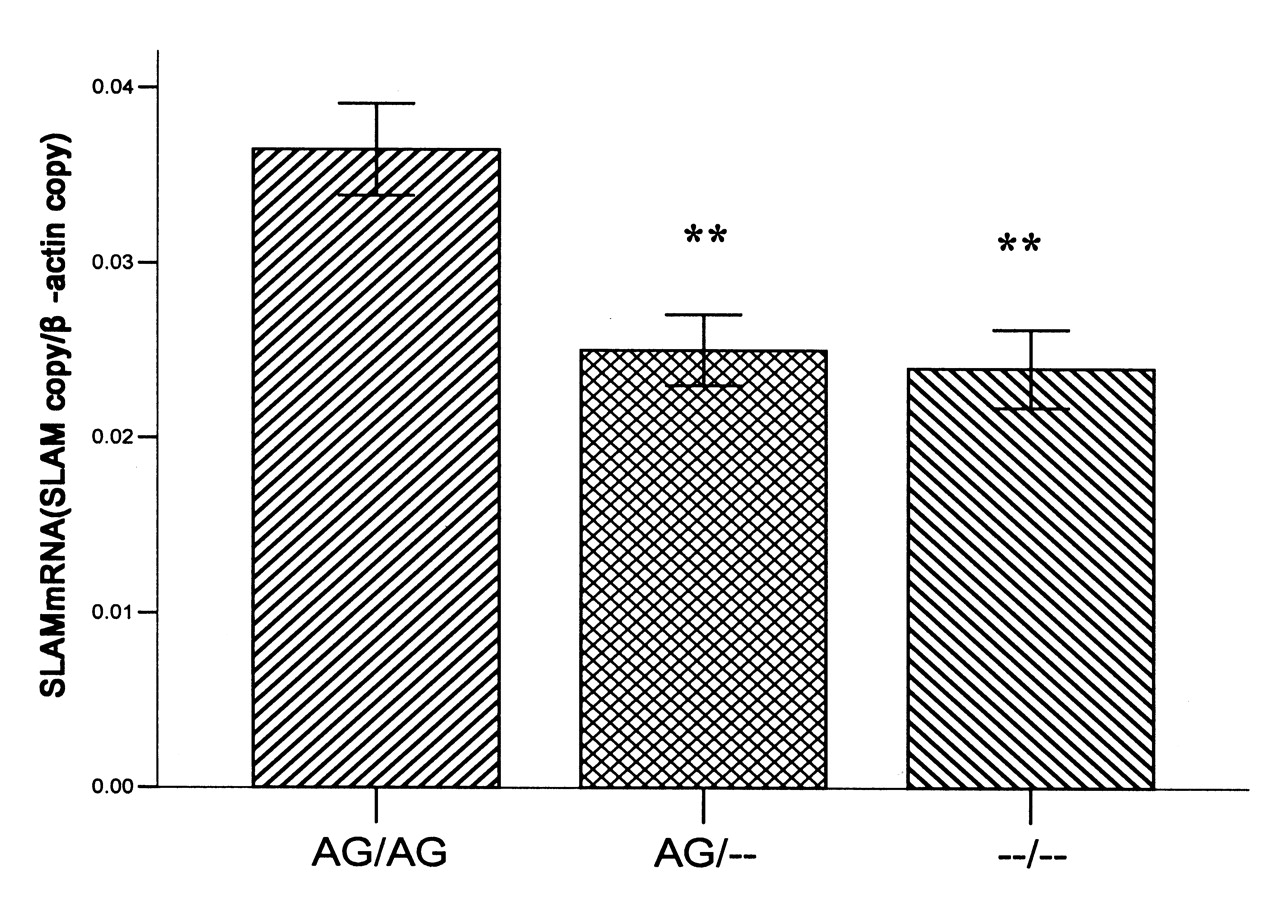
mRNA expression of SLAM in SLE patients with different genotypes; results are expressed as mean ± SD. Mean SLAM mRNA levels of 5 samples with −262AA/-188GG genotypes were significantly higher than for −262A-188G heterozygotes (0.037 ± 0.002 vs 0.025 ± 0.004, respectively; p < 0.01) and other genotypes (0.037 ± 0.002 vs 0.024 ± 0.004; p < 0.01). ** p < 0.01, compared with −262AA/−188GG genotypes.
DISCUSSION
We observed that −262A/T (rs2295614) and −188A/G (rs2295613) polymorphisms of SLAM gene were associated with increased risk for developing SLE in our Chinese study population. Carriers of the −262A-188G haplotype had 1.478-fold increased risk for developing SLE compared with noncarriers. Considered in our study, the 2 SNP at positions −262A/T and −188A/G are in the proximal upstream region of SLAM. Usually a promoter region is broadly defined as a 2-kb fragment of sequence located on the upstream region of the transcription start site. The position of these SNP within 2 kb upstream from the transcription start site suggests they may play a role in the process of transcription and regulate the gene expression18. We conducted the luciferase assay to investigate the functional relevance of promoter region variants of SLAM, and found that the −262A-188G construct had significantly higher luciferase reporter activity compared with −262T-188A. Moreover, the observations of SLAM expression of SLE with different genotypes were consistent with the reporter assays. We have shown that SLE patients with −262AA-188GG homozygotes had a statistically significantly higher mRNA expression of SLAM than the patients with −262A-188G heterozygotes and other genotypes. These findings suggest that the −262A-188G haplotype in SLAM gene promoter contributes to the increase of the transcription activities. Moreover, because only the SLE patients with −262AA-188GG homozygotes had statistically significantly higher mRNA expression, these results also may indicate a synergistic effect between −262A and −188G SNP in the context of haplotype on increasing the transcriptional activity of SLAM.
Some studies have investigated the association between polymorphisms of SLAM and autoimmune disease. Chabchoub, et al studied 3 SNP of the SLAM gene, rs2295612, rs1809963, and rs3796504; the results showed a lack of association of the SLAM gene and RA in French and Tunisian populations19. Suzuki, et al identified a linkage disequilibrium block associated with RA in the chromosome 1q region containing multiple SLAM family genes, but they failed to find an association between the SLAM gene and RA in their Japanese study population20. In that study, the frequencies of −262A were 72.73% in RA patients and 73.3% in controls, and frequencies of −188G were 72.1% in RA and 72.8% in controls20. In our study, the frequency of −262A allele in SLE (66.5%) was significantly higher than that in controls (55.2%). The frequency of −188G allele in cases and controls was 64.2% and 54.6%, respectively (p < 0.01). These results are consistent with the data from HapMap. The −262A allele and −188G allele are more prevalent in the Japanese population than in the Chinese population in the HapMap. Because different ethnic populations have different histories of mutation, migration, isolation, and genetic drift, there are substantial differences in allele frequencies among different ethnic groups21. While patients in our study were enrolled from several hospitals in Chongqing, they may not be representative of the general population. Further studies in larger or other populations will be required to clarify this.
The association of the signaling lymphocyte activation molecule (SLAM) family of receptors and their associated signaling adaptors is essential to regulate both innate and adaptive immune responses22. SLAM receptor is able to associate with the SLAM associated protein (SAP, SH2D1A) family adaptors, which is required for propagation of downstream signaling. They regulate lymphocyte-lymphocyte interactions involved in both cell-mediated and humoral immune responses. Recent evidence indicates that members of this family of receptors and signaling intermediates are also involved in autoimmunity. The creation of SLAM-deficient mice showed that SLAM deficiency severely compromises the production of IL-4 and IL-13; however, it has little effect on IFN-γ secretion. Such findings imply that SLAM is primarily involved in promoting TH2-cytokine secretion9. Another study also shows that deficiency in SH2D1A protected mice in an experimental lupus model, including the development of hypergamma-globulinemia, autoantibodies including anti-dsDNA, and renal disease. These findings demonstrate a specific role for the SLAM-SH2D1A system in regulation of T cell-dependent humoral immune responses23.
Our further evaluation of polymorphism associations showed that the SNP in the promoter region of SLAM altered its expression, and higher expression of SLAM was observed in individuals with susceptible alleles (−262A/−188G). Our findings might support the notion that genetic polymorphisms resulting in higher expression of SLAM may increase susceptibility to SLE. Considering that the SLAM functions as a costimulatory molecule and is primarily involved in promoting TH2-cytokine secretion, we expected that genetic polymorphisms resulting in higher expression of SLAM would influence the response of T cells. A more Th2-predominant immune response occurred in SLE patients, and probably was involved in the development of SLE disorder.
Our study showed there may be a synergistic effect between −262A and −188G SNP in the haplotype, which remains to be confirmed. Variations in the promoter region may potentially alter the affinities of existing protein-DNA interactions, or indeed may recruit new proteins to bind to the DNA, altering the specificity and kinetics of the transcriptional process24. Additional studies are required to conclusively determine whether potential transcription factors are involved in the different transcriptional activation of −262A/T and −188A/G.
Our study suggests that the −262A/T and −188A/G polymorphisms in the SLAM are genetic susceptibility factors for SLE in the Chinese population. Individuals possessing the AG/AG haplotype have increased susceptibility to SLE. Functional analysis indicates augmented expression of SLAM is associated with susceptibility to SLE. These findings suggest that the −262A-188G haplotype in the SLAM gene promoter contributes to risk for development of SLE by increasing the expression of SLAM.
Footnotes
-
Supported by grant No. 30800995 from the National Natural Science Foundation of China.
- Accepted for publication June 18, 2010.








{kind=link}
{kind=link}