

Abstract
Objective. Synovial angiogenesis, a critical determinant of juvenile idiopathic arthritis (JIA) pathogenesis, is sustained by various mediators, including vascular endothelial growth factor (VEGF) and osteopontin (OPN). We characterized the contribution of the local hypoxic environment to VEGF and OPN production by monocytic cells recruited to the synovium in JIA.
Methods. Paired synovial fluid (SF) and peripheral blood (PB) samples were collected from 20 patients with JIA. Mononuclear cells (MC) were isolated, and monocytic cells were purified by adherence, maintained in a hypoxic environment, or subjected to reoxygenation. VEGF and OPN protein concentrations were tested in SF, plasma, and culture supernatants by ELISA, and mRNA expression was assessed in freshly purified and cultured cells by reverse transcriptase-polymerase chain reaction. Synovial tissue was obtained at synovectomy from 4 patients with JIA, and analyzed by immunohistochemistry for VEGF, OPN, CD68, and hypoxia-inducible factor-1α (HIF-1α).
Results. VEGF mRNA expression was increased in SFMC and SF monocytic cells compared to matched PBMC and PB monocytic cells or SF lymphocytes, correlating with significantly higher protein levels in SF relative to plasma samples. Accordingly, OPN mRNA expression in SF monocytic cells was associated with significant increase of SF protein. Immunohistochemistry revealed the presence of both factors in synovial tissues at the level of the lining and sublining layers, which colocalized with intense CD68 and HIF-1α staining, suggesting production by hypoxic synovial monocytic cells. VEGF and OPN expression was abrogated upon SF monocytic cell reoxygenation and maintained by exposure to prolonged hypoxia.
Conclusion. Hypoxic synovial monocytic cells are a likely source of VEGF and OPN in JIA. These data point to a role for hypoxia in the perpetuation of synovitis in JIA.
- JUVENILE IDIOPATHIC ARTHRITIS
- MONOCYTES
- HYPOXIA
- ENVIRONMENT
- SYNOVIAL FLUID
- VASCULAR ENDOTHELIAL GROWTH FACTOR
- OSTEOPONTIN
Juvenile idiopatic arthritis (JIA) consists of a clinically heterogeneous group of chronic pediatric arthritides with poorly defined etiology characterized by synovial joint inflammation, which results in synovial tissue hyperplasia, progressive cartilage destruction, and bone erosion1. The chronicity of inflammation is crucial to the pathogenesis of JIA, and persists as a consequence of extensive angiogenesis and progressive infiltration and activation of inflammatory cells in synovial tissue and synovial fluid (SF) of affected joints1–4.
Local release of proangiogenic mediators is responsible for the uncontrolled endothelial cell (EC) proliferation that takes place in many angiogenesis-dependent diseases, including rheumatic arthropathies5–7. The EC-specific mitogen, vascular endothelial growth factor (VEGF), plays a central role in driving blood vessel neo-formation in the rheumatoid synovium via stimulation of EC survival, proliferation, and chemotaxis, and in monocytic cell recruitment/activation3,6,8–10. Another proangiogenic mediator potentially implicated in the pathogenesis of inflammatory arthritis is the cytokine/extracellular matrix protein, osteopontin (OPN)11,12, which is endowed with EC adhesive capacity and represents a chemotactic stimulus for EC, monocytic cells, and T lymphocytes9,13–16. Abundant VEGF and OPN expression was detected in the inflamed joints of patients with JIA, showing a good correlation with the degree of vascularization and disease activity measures3,17,18. However, the contribution of individual cell types and the role of environmental signals to VEGF and OPN production in JIA have not been assessed, hindering understanding of the pathogenetic mechanisms of the disease and the design of targeted therapies1.
One of the major promoters of angiogenesis is hypoxia, a local decrease in partial pressure of oxygen (pO2) that characterizes inflammatory and neoplastic lesions and is mediated by release of proangiogenic factors10,19–22. The hypoxic microenvironment is a hallmark of the inflamed synovium, and its importance in the pathogenesis of adult arthropathies was documented20,21,23–26.We have recently demonstrated the hypoxic nature of the synovium of patients with JIA and the constitutive expression of hypoxia-inducible factor-1α (HIF-1α) and HIF-2α, key regulators of hypoxia transcriptional response27, in monocytic cells recruited to inflamed joints in JIA28. Further, we demonstrated hypoxic induction of the chemokine CCL20 in JIA synovial monocytic cells28, suggesting that the hypoxic synovial microenvironment may directly contribute to the persistence of JIA synovitis by increasing inflammatory cell infiltration in the affected joints.
Activated monocytic cells are proangiogenic in different physiopathological conditions29,30, and induction of the monocytic cell proangiogenic phenotype is associated with expression of HIF-1α31,32. We investigated the role of monocytic cells infiltrating the JIA synovium in the production of VEGF and OPN and the contribution of the local hypoxic microenvironment to their expression, using an experimental protocol designed to prevent cell reoxygenation after the harvest from inflamed joints.
MATERIALS AND METHODS
Study population
Twenty patients were enrolled consecutively in this study provided they met the International League of Associations for Rheumatology Durban criteria for classification of JIA1 and had active knee disease, with clinical effusion and swelling of the joint at the time of therapeutic arthrocentesis. Patients with JIA were classified into 3 subtypes depending on the onset and course of disease: persistent oligoarticular JIA (pOJIA, oligoarticular onset and course); extended oligoarticular JIA (eOJIA, oligoarticular onset and polyarticular course); and polyarticular JIA (pJIA, polyarticular onset and course). Thirteen patients belonging to all disease subtypes had been previously treated with methotrexate (MTX) and/or nonsteroidal antiinflammatory drugs (NSAID), and only one patient with pJIA had been treated with a tumor necrosis factor inhibitor; the remaining 6 patients with pOJIA had never received any systemic antiinflammatory drugs. All patients underwent SF aspiration from the knee for steroid injection due to a recent relapse of the disease. A previous steroid injection in the same joint in the last 6 months was considered as an exclusion criterion. Patients who were having systemic antiinflammatory treatment at the time of arthrocentesis were also excluded. No power calculation was used to determine potential statistically significant differences between the groups and no attempt was made to segregate patients on the basis of disease duration, number of active joints, or therapeutic regimen. Informed consent was obtained prior to sample collection according to the procedure approved by our institution’s ethical committee. Clinical characteristics of patients, various laboratory measures of disease activity, and previous therapeutic regimens are reported in Table 1.
Clinical and laboratory features of patients with juvenile idiopathic arthritis at the time of sampling. Results are expressed as mean (range).
SF samples were collected at the time of therapeutic knee arthrocentesis into sodium-heparin tubes under vacuum, and their pO2 levels were monitored with a portable trace oxygen analyzer (Oxi 315i/set, WTW, Germany). Paired peripheral blood (PB) samples were obtained on the occasion of routine venepuncture and collected as for SF. PB from 5 age-matched control subjects undergoing routine venepuncture for minor surgical procedures was collected after informed consent of the parents. A history of inflammatory or infection disorders in the 4 weeks before examination, and clinical or laboratory signs of inflammation at the time of blood sampling, was considered as an exclusion criterion. Specimens were centrifuged to prepare cell-free SF and plasma and stored at −80°C. Synovial tissues were obtained from 4 patients with oligoarticular-onset JIA undergoing synovectomy or arthroscopic biopsy after informed consent of the parents, fixed in 10% formalin, and embedded in paraffin.
Monocytic cell purification and culture
SF mononuclear cells (SFMC) and PB mononuclear cells (PBMC) were isolated by density gradient centrifugation over a Ficoll cushion (Amersham, Milano, Italy) and resuspended in RPMI-1640 (Euroclone, Celbio, Milano, Italy) supplemented with 10% heat-inactivated fetal calf serum (HyClone, Celbio). SFMC and PBMC were separated into adherent (monocytic) and nonadherent (lymphocyte) cells by incubation for 1 h at 37°C in tissue culture plates (Costar, Celbio), and their purity was assessed by flow cytometry (see below). For a subset of samples, monocytic cells were purified by negative selection using MACS magnetic beads according to standard methods (Human Monocyte Isolation kit-II, Miltenyi Biotec, Bologna, Italy). To avoid cell reoxygenation, SF-derived samples were handled in a 37°C humidified anaerobic work station incubator (BUG-BOX, ALC-International, Milano) flushed with a gas mixture of 94% N2, 5% CO2, and 1% O2. In a few experiments, SF monocytic cells were reoxygenated for 16 h by incubation at 37°C in a humidified atmosphere containing 75% N2, 5% CO2, and 20% O2 or maintained for 16 h under prolonged hypoxia (1% O2). pO2 in culture medium was monitored with the oxygen analyzer, as described33. The endotoxin content, determined by the Limulus amebocyte test (Bio-Whittaker, Walkersville, MD, USA), was < 0.125 EU/ml in all reagents.
Flow cytometry analysis
The purity of the adherent and nonadherent cell fractions isolated from SFMC and PBMC was determined by flow cytometry. Cells were preincubated for 20 min at 4°C with phosphate buffered saline (PBS) supplemented with 2% human AB serum and 0.05% NaN3 to block nonspecific labeling. Surface staining was carried out with FITC or phycoerythrin (PE)-conjugated anti-CD14 (a pan-monocytic cell marker), PE-conjugated anti-CD3 (a pan-T cell marker), and IgG2a isotype-matched monoclonal antibodies (mAb; BD-Pharmingen, Milano, Italy) for 30 min at 4°C. Intracellular staining with FITC-conjugated anti-CD68 (a pan-monocytic cell marker) or IgG1 isotype-matched mAb (BD-Pharmingen) was performed after cell fixation in 4% paraformaldehyde and permeabilization in PBS containing 1% AB serum and 0.1% saponin (Sigma). Fluorescence was quantified on a FACScan flow cytometer equipped with CellQuest software (BD Biosciences). The percentage of monocytic cells in the adherent cell population ranged between 93% and 95% in the different samples, as determined by positivity for the CD14 and CD68 markers, whereas the nonadherent fractions were negative for CD14 and composed predominantly by CD3+ T cells at a purity of ≥ 83%. Viability was ≥ 95%, as determined by propidium iodide staining.
ELISA
Secreted VEGF and OPN were measured in SF, plasma, and cell-free supernatants by ELISA, using specific Quantikine immunoassay kits (R&D Systems, Space, Milano, Italy). Optical density was determined by a Tecan Spectrafluor Plus plate reader (Tecan, Mannedorf, Switzerland) at 450 nm. Data were analyzed with the Graph Pad Prism 3 Software (Graph Pad, San Diego, CA, USA).
Reverse transcriptase-polymerase chain reaction (RT-PCR)
Total RNA was purified from single-cell suspensions by the Qiagen RNeasy Mini Kit. Control of RNA integrity was carried out by electrophoresis with an Agilent Bioanalyzer 2100 (Agilent, Waldbroon, Germany). RNA (1 μg) was reverse-transcribed into double-stranded cDNA on a GeneAmp PCR System 2700 (Applied Biosystems, Milano, Italy), using the Advantage RT-for-PCR Kit (BD-Pharmingen). For semiquantitative RT-PCR, cDNA was amplified with recombinant TaqDNA polimerase (Invitrogen), as described33. Products were electrophoresed on a 1.2% agarose gel and visualized by ethidium bromide staining. Real-time quantitative PCR (qRT-PCR) was performed on a 7500 Real-time PCR System (Applied Biosystems), using SYBR Green PCR Master Mix and 300 nM of the following sense and antisense oligonucleotide primers (TIBMolbiol, Genova, Italy): VEGF, forward, 5’-GCA GCT TGA GTT AAA CGA ACG-3’; reverse, 5’-GCA GCG TGG TTT CTG TAT C-3’; OPN, forward, 5’-TGA CCC ATC TCA GAA GCA G-3’; reverse: 5-ATG GCT TTC GTT GGA CTT AC, as described33.
Immunochemistry
5 ×104 SF monocytic cells were applied to polylysine glass slides, fixed in 4% PAF, and permeabilized with 0.1% Triton X-100. VEGF protein expression was assessed by immunostaining of cytocentrifuge slides with anti-hVEGF rabbit antibody (Santa Cruz Biotechnology, Santa Cruz, CA, USA), and detection was carried out with anti-rabbit Ig conjugated to HRP-labeled dextran polymer (Dako, Glostrup, Denmark), as described33. Isotype-matched mAb was used as control.
Immunohistochemical labeling of tissue specimens was performed by a 3-step immunoperoxidase technique18. Paraffin-embedded serial tissue sections were incubated at room temperature for 30 min with anti-CD68 (Dako), anti-CD34 (Dako), anti-VEGF (Santa Cruz), and anti-OPN (Abcam, Cambridge, UK) antibody, or at 4°C overnight with anti-HIF-1α mAb (Abcam). Sections were reacted with anti-mouse (for CD68 and CD34) and anti-rabbit (for VEGF and OPN) Ig conjugated to peroxidase- labeled dextran polymer (Dako). For HIF-1α detection, the tyramide signal amplification system kit was used (Dako). Isotype-matched antibodies were used as negative controls. Chromogenic diaminobenzidine substrate was then applied. Slides were counterstained with Mayer’s hematoxylin and examined under a phase-contrast microscope (Olympus, Milano, Italy).
Statistical analysis
VEGF and OPN concentrations in SF or plasma were compared among the 3 subgroups of patients and controls using the nonparametric Mann-Whitney U-test (p < 0.05 as statistically significant). Concomitant cytokine determinations in paired SF and plasma specimens were evaluated by the Wilcoxon rank test.
RESULTS
Monocytic cells are an important source of VEGF in JIA SF
Initial experiments were carried out to measure VEGF release into SF from patients with JIA. Elevated amounts of VEGF were detected in the SF from all disease subtypes, although with some degree of patient to patient variability (Figure 1A), with higher concentrations in children with polyarticular onset (pJIA, median 1321 pg/ml) or course of disease (eOJIA, median 1090 pg/ml), compared with patients with an oligoarticular onset and course (pOJIA, median 820 pg/ml). Globally, median protein concentrations were significantly increased (p < 0.0001) in SF relative to paired plasma samples (Figure 1A, inset), ranging from 2.8-, 4.2-, and 5.9-fold in pOJIA, eOJIA, and pJIA subgroups, respectively. When these data were analyzed in relation to the clinical characteristics of patients, we did not find any statistically significant correlation between VEGF levels and disease duration or severity, number of active joints involved, or therapeutic regimen.

VEGF expression in JIA SF and SF-derived MC. (A) VEGF concentrations were quantified by ELISA in SF and plasma from 20 patients with JIA. Individual samples were run in triplicate. Results for each sample (dots) are pg/ml. Horizontal lines represent median values for each group. *Polyarticular disease. Inset: VEGF concentrations were compared in paired SF and plasma samples. Boxes show 25th–75th percentiles, whiskers the highest and lowest values for each subgroup. p value byWilcoxon rank test. (B) VEGF mRNA expression was analyzed by semiquantitative RT-PCR on total RNA purified from the indicated pairs of freshly isolated PBMC and SFMC from patients with JIA. ß-actin mRNA was used to control for input RNA. (C) VEGF transcript levels were quantified by qRT-PCR in SFMC- and PBMC-derived RNA pools from samples shown in (B), and expression was calculated in relation to values of 2 reference genes. Data are expressed as mean normalized values from triplicate determinations.
The cellular source of VEGF in JIA SF was then investigated. VEGF expression was first assessed by RT-PCR in SFMC from a subset of 8 children with different forms of JIA and compared with that of paired PBMC (Figure 1B). VEGF mRNA was constitutively expressed in all SFMC samples, although it varied among different individuals, with high levels present in 7/8 samples and a lower expression detectable in one sample. The variability in VEGF transcript levels did not correlate with the disease subtype or type of treatment. In contrast, minimal or undetectable mRNA levels were found in matched PBMC samples, suggesting that VEGF expression is upregulated in cells recruited to the inflamed synovium. Equal amounts of RNA purified from the samples shown in Figure 1B were combined according to disease subtype, and RNA pools were subjected to qRT-PCR. VEGF mRNA expression was significantly higher in SFMC than in PBMC from all disease subtypes (Figure 1C), confirming the semiquantitative analysis.
The monocytic cell and lymphocyte cell fractions of SFMC (Figure 2A) were then isolated from 3 randomly selected patients, each representative of a specific disease subtype, and tested for VEGF expression by RT-PCR (Figure 2B). Both cell types constitutively expressed VEGF mRNA, but transcript levels were higher in SF monocytic cells (SF-Mn) from all the patients tested, suggesting that VEGF was transcribed predominantly in the monocytic cells than in the lymphocyte cell population present in the JIA SF. These findings were confirmed by qRT-PCR, which demonstrated a 15-fold higher VEGF expression in SF monocytic cell-derived compared to SF lymphocyte (SF-Ly)-derived RNA pools (data not shown). Interestingly, SF monocytic cells expressed increased VEGF mRNA compared to monocytic cells isolated from paired PBMC (PB-Mn; Figure 2C, upper panel). These findings were confirmed by qRT-PCR mRNA quantification, which revealed 3.9-, 4.1-, and 4.4-fold increases in SF monocytic cells relative to matched PB monocytic cell preparations (Figure 2C, lower panel). Comparable VEGF transcript levels were detectable in PB monocytic cells purified from patients with JIA and controls (Figure 2C). A similar pattern of results was observed in monocytic cells purified from the other 5 patients (data not shown). mRNA expression in SF monocytic cells was paralleled by VEGF protein immunostaining (Figure 2D). These data indicate that VEGF was significantly upregulated in monocytic cells recruited to the inflamed joints of patients with JIA, suggesting a likely role for SF monocytic cells as a source for this factor and raising the issue of the stimuli controlling its production in the synovial environment.

Comparison of VEGF expression in monocytes (Mn) and lymphocytes (Ly) from JIA SF. Monocyte and lymphocyte fractions were isolated in parallel from pairs of SFMC and PBMC by adherence to plates. (A) Flow cytometric analysis. Purity of adherent and nonadherent cell cultures was determined by FACS after staining with indicated mAb. Left panel: Full and shaded histograms represent fluorescent profile of cells stained with anti-CD14 and anti-CD68 mAb within the whole population of adherent cells, relative to cells stained with isotype-matched antibody (empty histogram). Percentage of cells positive for both monocyte markers is indicated (M1). Right panel: Full and shaded histograms indicate fluorescent profile of cells stained with anti-CD3 and anti-CD14 mAb within the whole population of nonadherent cells, relative to cells stained with control antibody (empty histogram). Percentage of CD3+ cells is indicated (M1). (B and C, upper panel) Total RNA from SF monocytes, SF lymphocytes, and PB monocytes from children with JIA and PB monocytes from healthy controls was tested for VEGF expression by semiquantitative RT-PCR (see Figure 1B). Data shown are from 3 representative patients and 2 controls. (C, lower panel) VEGF transcript levels were quantified by qRT-PCR in paired PB monocytes (open bars) and SF monocytes (hatched bars) from 3 patients with JIA and PB monocytes from 2 controls (shaded bars), as detailed in Figure 1C. Numbers represent mRNA fold-increase in SF monocytes relative to matched PB monocytes (arbitrarily defined as = 1). (D) VEGF protein expression was evaluated by immunostaining on cytocentrifuge slide preparations of SF monocytes. Hematoxylin-counterstained slides were examined under a phase-contrast microscope (original magnification 40×). VEGF immuno-reactivity is detectable in cell cytoplasm (dark staining). Staining with a control mAb (IgG1) is shown for Patient 17.
Hypoxia induces VEGF production by SF monocytic cells
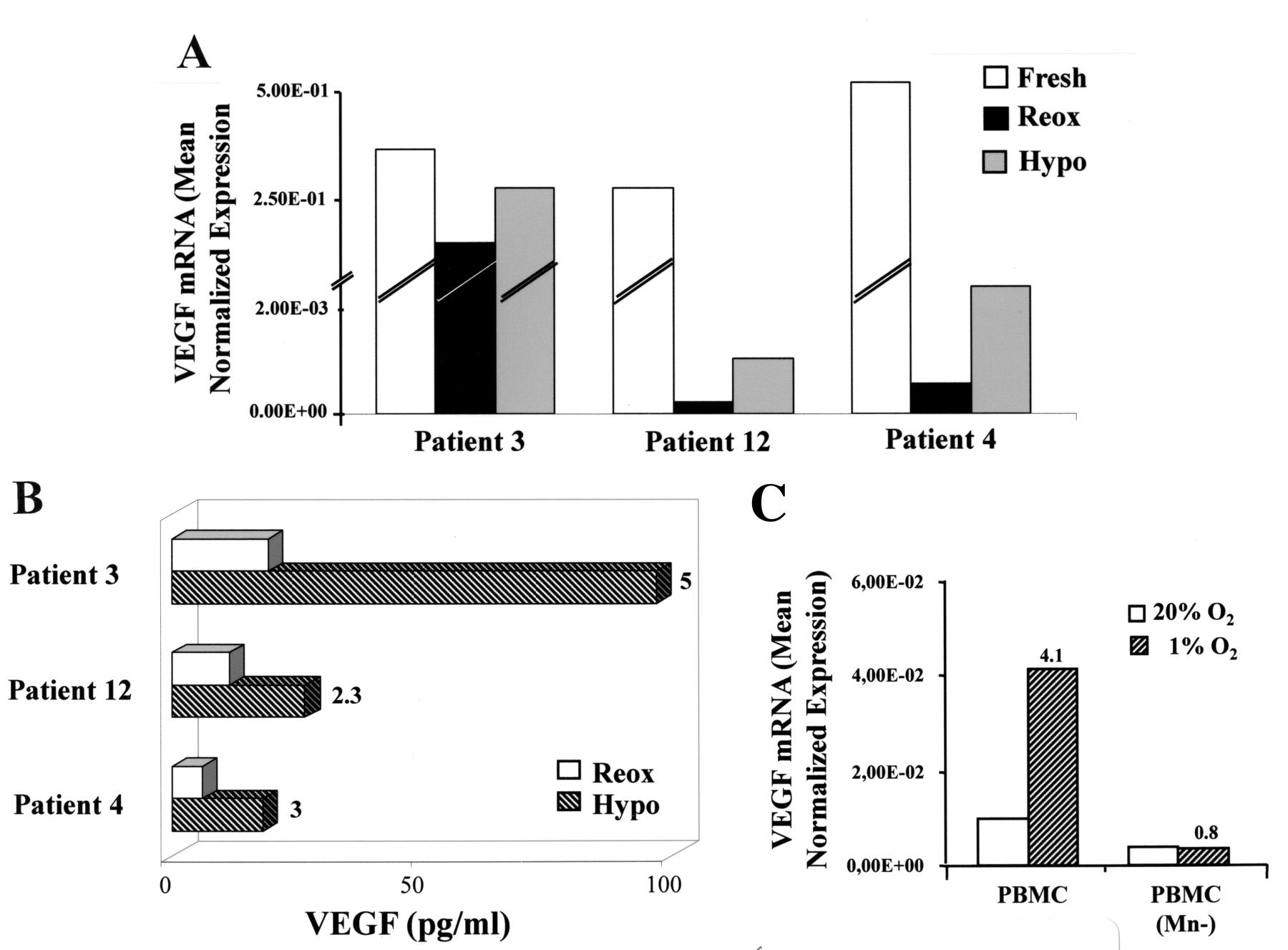
We next investigated whether the pathological low O2 levels of the rheumatoid synovial environment23,25 induced VEGF expression in SF monocytic cells from patients with JIA. Freshly purified SF monocytic cells were exposed for 16 h to ambient pO2 (20% O2, hereafter referred to as reoxygenation) or were maintained under a controlled hypoxic environment (1% O2) for the same length of time, and VEGF transcript levels were then quantified (Figure 3A). Interestingly, reoxygenation caused a pronounced reduction of VEGF mRNA expression relative to fresh cells, and this effect was in part counteracted by culture under hypoxia. Accordingly, VEGF secretion by SF monocytic cells maintained under the hypoxic environment exceeded by 2.3- to 5-fold that of reoxygenated cultures (Figure 3B). A similar pattern of results was observed in SF monocytic cells purified from all the patients listed in Figure 1B (data not shown), suggesting that hypoxia is a stimulus for VEGF production by SF monocytic cells in JIA. The potential role of monocytic cells as a source for VEGF within the hypoxic synovial environment was supported by data showing that VEGF mRNA expression was upregulated by more than 4-fold in PBMC cultured under low O2 concentrations (1%) similar to those present in the JIA SF28, and that this response was completely abrogated upon depletion of monocytic cells (Figure 3C).

Hypoxic regulation of VEGF production by SF monocytes. (A) SF monocytes purified from 3 randomly selected patients with JIA were cultured for 16 h under normoxic (Reox) or hypoxic (Hypo) conditions. Total RNA from fresh (open bars), reoxygenated (full bars), or hypoxic (shaded bars) SF monocytes was tested for VEGF mRNA levels by qRT-PCR. Data are expressed as in Figure 1C. (B) Conditioned medium from reoxygenated (open bars) and hypoxic (hatched bars) cultures was assayed for VEGF content by ELISA. Results are expressed as pg per 5 × 105 cells/ml. Numbers indicate fold-increase of secreted protein in hypoxic relative to reoxygenated cells (arbitrarily defined as = 1). (C) VEGF transcript levels were quantified by qRT-PCR in PBMC-derived RNA pools from 3 representative patients exposed to 20% or 1% O2 for 16 h, before or after monocyte depletion (Mn–). Data are expressed as mean normalized values from triplicate determinations. Numbers represent mRNA fold-increase at 1% relative to 20% O2 (defined as = 1).
OPN production is enhanced by hypoxia in SF monocytic cells from patients with JIA
A crosstalk between VEGF and OPN was previously established in endothelial cells and shown to amplify the angiogenic response9,14. To determine whether VEGF overexpression in JIA SF was associated with increased OPN release, OPN protein concentrations were assessed in the same specimens tested for VEGF (Figure 4A). A significant increase in median OPN levels (p < 0.0001) was detected in SF relative to paired plasma samples (Figure 4A, inset), paralleling that of VEGF. Higher OPN concentrations were found in SF from pJIA (median 325.3 ng/ml) and eOJIA (median 319.9 ng/ml) subgroups compared to the pOJIA subtype (median 235.2 ng/ml), with an average increase of 9-, 7.6-, and 6.2-fold with respect to the corresponding plasma specimens (Figure 4A), thus showing a trend for a positive correlation between OPN and VEGF production in JIA SF. As for VEGF, OPN levels were independent of disease duration or activity, number of active joints involved, or type of treatment. Quantification of OPN transcript levels by qRT-PCR in SF monocytic cell and PB monocytic cell-derived RNA pools demonstrated increased OPN expression in SF monocytic cells isolated from patients with JIA compared to paired PB monocytic cells or to PB monocytic cells purified from controls (Figure 4B), reflecting the protein results, and higher OPN transcript levels were quantified in the monocytic cells than in the lymphocyte fractions of SFMC (data not shown), as shown for VEGF.
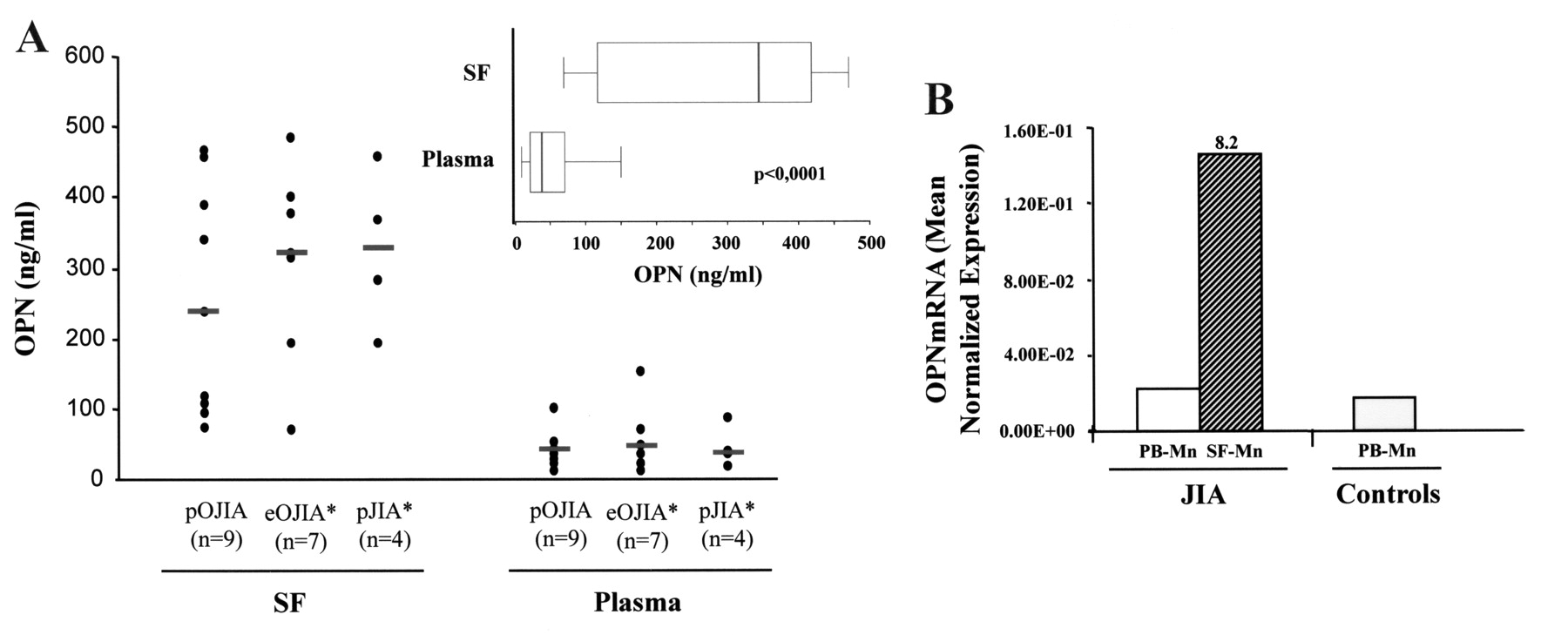
OPN expression in JIA SF and SF-derived monocytes. (A) OPN concentrations were quantified by ELISA in the same pairs of JIA SF and plasma specimens tested for VEGF content. Individual samples were run in triplicate. Results for each sample (dots) are ng/ml. Inset: OPN concentrations were compared in paired SF and plasma samples, as in Figure 1A. (B) OPN transcript levels were quantified by qRT-PCR in RNA pools from paired PB monocytes (open bar) and SF monocytes (hatched bar) isolated from 5 patients with JIA, and from control PB monocytes (shaded bar), and expression was calculated in relation to values of reference genes. Data are expressed as in Figure 1C.
Next, we investigated the possible relationship between hypoxia and OPN production by SF monocytic cells. As for VEGF, fresh SF monocytic cells constitutively expressed elevated levels of OPN mRNA, which were strongly reduced by reoxygenation, whereas OPN transcript expression was significantly higher in hypoxic relative to reoxygenated cells (Figure 5A). Accordingly, OPN protein secretion was increased in hypoxic relative to reoxygenated SF monocytic cells, ranging between 2.4- and 4.8-fold in the samples tested (Figure 5B). These data suggest a regulatory role for hypoxia in SF monocytic cell production of OPN in JIA. OPN mRNA expression was upregulated by more than 3-fold in PBMC cultured at 1% O2, and this response was partially abrogated upon monocytic cell depletion (Figure 5C), confirming the potential role of these cells as OPN producers in response to the low pO2 present in the JIA synovial environment23,25,28.

Hypoxic regulation of OPN in SF monocytes. (A) Total RNA purified from the same SF monocyte samples as in Figure 3 was tested for OPN mRNA levels by qRT-PCR. Data are expressed as in Figure 3A (B) Conditioned medium from reoxygenated (open bars) and hypoxic (hatched bars) SF monocyte cultures was assayed for OPN content by ELISA. Results are expressed as ng per 5 × 105 cells/ml. Numbers represent fold-increase at 1% relative to 20% O2 (defined as = 1). (C) OPN transcript levels were quantified by qRT-PCR in PBMC-derived RNA pools from 3 representative patients exposed for 16 h to 20% or 1% O2, before or after monocyte depletion (Mn–). Data are expressed in Figure 3C.
Monocytic cells recruited to the hypoxic JIA synovium express VEGF and OPN proteins
Experiments were carried out to measure VEGF and OPN expression in vivo in monocytic cells infiltrating the hypoxic areas of the JIA synovium. Tissue speciments obtained at synovectomy from a subset of patients with JIA were analyzed by immunohistochemistry. Serial sections were immunostained for the hypoxia marker HIF-1α (Figure 6A), the pan-monocytic cell marker CD68 (Figure 6B), the endothelial cell marker CD34 (Figure 6E), and for VEGF (Figure 6C) and OPN (Figure 6D), and counterstained with hematoxylin-eosin (Figure 6F). Clear positivity for HIF-1α was observed in the lining and, with lower intensity, in the sublining layers of synovial membranes, in areas characterized by intense CD68 staining, suggesting its induction in infiltrating monocytic cells, although a few positive fibroblast-like cells (CD68–) were also detectable. VEGF and OPN immunoreactivity was observed in the same areas of serial tissue sections, confined almost entirely to cells positive for HIF-1α and CD68. No staining was detectable with isotype-matched control antibodies (data not shown). These data suggest that monocytic cells recruited to the JIA synovium respond to the local hypoxic environment with production of VEGF and OPN. Staining with anti-CD34 mAb showed some positivity for VEGF and OPN around areas of vascularization, consistent with the proangiogenic role of both cytokines.
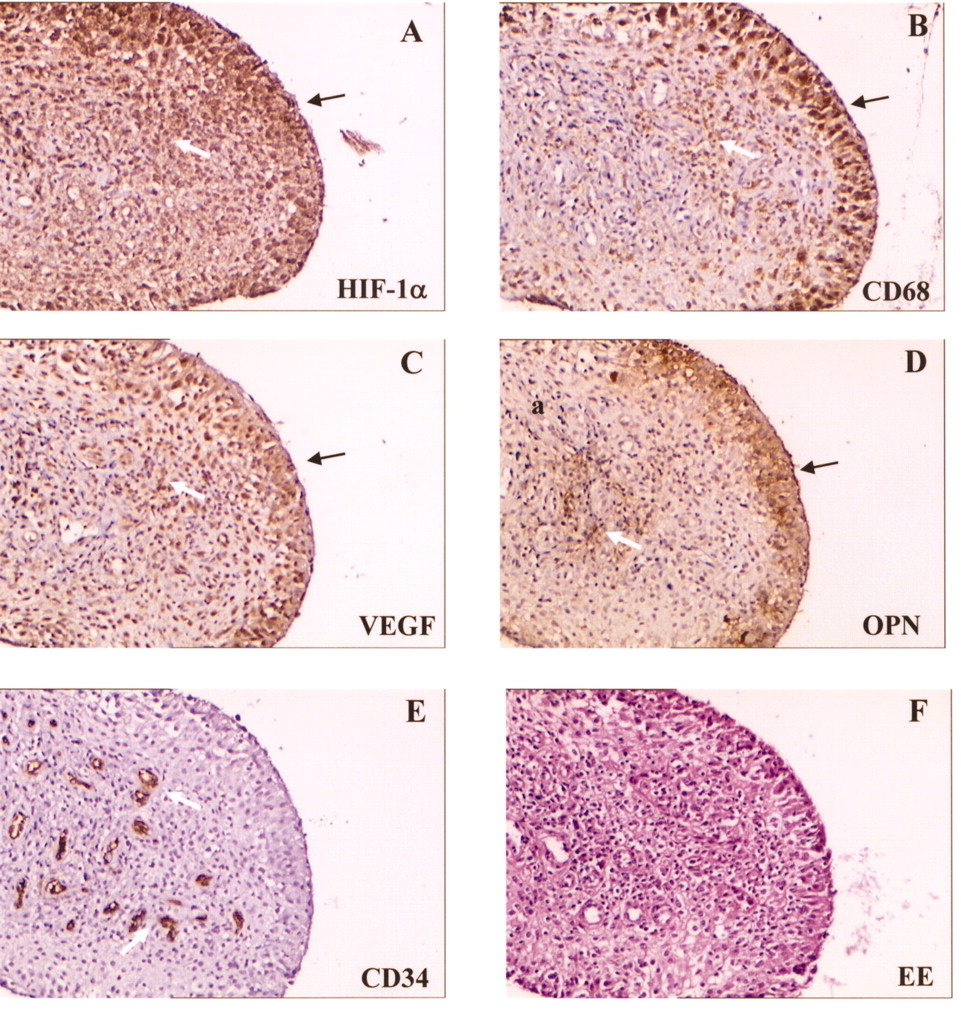
Immunohistochemical detection of VEGF and OPN proteins in hypoxic monocytes infiltrating the JIA synovium. Synovial tissue was obtained at synovectomy from a representative patient (Patient 3), and staining was carried out on serial sections with the indicated mAb. Hematoxylin-eosin staining (EE) is shown in panel F as a control. Slides were examined under a phase-contrast microscope (original magnification 10×). Positive cells (stained brown) are detectable at level of lining layer (black arrows) and sublining layer (white arrows). Results are representative of 4 patients.
DISCUSSION
Angiogenesis and chronic inflammation are codependent because an inflammatory state can promote new vessel formation, which, in turn, allows the migration and progressive infiltration of other inflammatory cells and perpetuation of the inflammatory state34. Soluble mediators produced by stromal and inflammatory cells drive angiogenesis and are crucial to the perpetuation of synovial inflammation in rheumatoid arthritides2,5–8,12,15,21,35. In our study, we observed abundant release of the proangiogenic and chemotactic mediators VEGF and OPN into the inflamed joints of patients with JIA and identified synovial monocytic cells as a likely cellular source for both factors. Further, we provide the first evidence that the hypoxic synovial environment is a stimulus for their production in patients with JIA.
Elevated VEGF levels accumulated in JIA SF as compared to matched plasma samples, in particular in patients with pJIA and eOJIA relative to the pOJIA subgroup, confirming previous findings18. A parallel significant increase in the amounts of OPN was measured in SF with respect to matched plasma specimens. These results suggest a trend for a positive correlation between OPN and VEGF production in JIA SF. A possible relationship between the 2 factors is also suggested by the observation of their similar distribution at the level of the lininig layer of JIA synovial membranes. These data are in accord with our previous findings demonstrating a clear positive correlation between VEGF and OPN synovial expression and degree of angiogenesis, and suggest a functional cooperation between the 2 factors in the control and stimulation of vascularization in the inflamed joint of patients with JIA. A crosstalk between VEGF and OPN was previously established and shown to amplify the angiogenic response9,14,36. Moreover, expression of various angiogenic factors, including VEGF, in fresh JIA membranes was shown to correlate with increased tissue angiogenesis and revascularization of human synovium engrafted into SCID mice3. Thus, production of higher amounts of VEGF and OPN in patients with pJIA and eOJIA, relative to the pOJIA subgroup, may be clinically relevant, resulting in a greater extent of joint vascularization and inflammation and in a major severity of the disease.
SFMC, and in particular the monocytic cell fraction, were identified as a possible source for both cytokines. Expression of VEGF and OPN was significantly higher in SF-derived than in matched PB-derived cells from all disease subtypes or PB monocytic cells from control subjects, indicating that recruitment to the local synovial environment is a stimulus for their production. Further, mRNA was expressed at increased levels in SF monocytic cells compared to SF lymphocytes present in the JIA SF, and was paralleled by protein expression, expanding previous evidence on the predominant role of synovial monocytic cells rather than synovial lymphocytes for the production of proinflammatory factors such as CCL2028, CCL3, and CXCL1037 in JIA. In contrast, findings reported for adult arthritides, such as rheumatoid arthritis (RA) and osteoarthritis (OA), suggest that the distribution of VEGF and OPN in the inflamed synovium was preferentially confined to fibroblasts and T cells12,21,38.
Hypoxia is a common feature of the rheumatoid synovium and has been implicated in the pathogenesis of adult arthropathies21,23–26,39. HIF-1α was found constitutively expressed in the lining and, to a lesser extent, the sublining layers of the JIA synovium, confirming the presence of reduced pO2 in the synovia of patients with JIA. Positivity for HIF-1α was mostly colocalized with CD68-specific staining in serial tissue sections, and was associated with intense immunoreactivity for both VEGF and OPN. These findings support an active role of monocytic cells migrating to the JIA synovium as VEGF and OPN producers, and suggest that cytokine overexpression in synovial monocytic cells is probably a result of activation of the HIF-1α pathway by local hypoxia.
The possibility that hypoxia stimulates synovial monocytic cells to produce VEGF and OPN in JIA is supported by the observation that constitutive VEGF and OPN mRNA levels detectable in freshly isolated SF monocytic cells decreased upon normoxic in vitro cell culture, whereas maintenance of the hypoxic environment sustained their expression. Accordingly, cell incubation in prolonged hypoxia resulted in increased production of both cytokines compared to reoxygenated cells. These data extend earlier findings that VEGF can be upregulated by hypoxia in synovial fibroblasts from patients with RA and OA21,24,38, and provide the first evidence that hypoxia can induce OPN expression in synovial cells, linking hypoxia signaling to activation of the angiogenic process in JIA. The observation that VEGF and OPN induction in PBMC cultured under low O2 concentrations similar to those present in the inflamed JIA synovium23,25 was almost completely abrogated upon depletion of monocytic cells is noteworthy, and is consistent with the possible role of hypoxic synovial monocytic cells as a source for both factors. However, the observation of higher levels of VEGF and OPN expression in fresh SF monocytic cells than in cells maintained in vitro under hypoxic conditions suggests that additional factors, such as proinflammatory cytokines present in the rheumatoid synovial environment10,14,26,34,40, may contribute to the hypoxia stimulatory effects in vivo. Evidence that hypoxia may interact with immune-derived stimuli in the regulation of gene expression in monocytic cells and in modulating proinflammatory mediator production in synovial cells was previously reported38,40–43. Future studies to examine the effects of hypoxia and proinflammatory cytokines in combination will yield a more complete picture of the functional behavior of monocytic cells within the pathological intraarticular environment in JIA.
The relationship between synovial hypoxia and monocytic cell expression of VEGF and OPN expands previous observations on the relevance of hypoxia for the activation of monocytic cell proangiogenic phenotype in other pathological conditions10,30–32, and indicates that a dynamic change in production of proangiogenic mediators by synovial monocytic cells may occur depending on the O2 availability. These findings also highlight the importance of performing in vitro experiments under conditions that mimic those occurring in vivo within JIA rheumatic joints for understanding the pathogenetic mechanisms of the disease. Because mobilization causes arthritic joints to undergo cycles of hypoxia-reoxygenation23,25,40,44, SF monocytic cells may be subjected to different degrees of oxygenation within diseased joints, resulting in shifting gradients of secreted VEGF and OPN and consequently in the variable extent of synovial vascularization. Extreme O2 situations were tested in this study. pO2 in rheumatic joints ranged between 6 and 50 mm Hg (0.8%–7% O2), whereas physiological O2 concentrations in healthy joints range between 50 and 70 mm Hg (7%–10% O2)23. One percent of O2, a common experimental condition of hypoxia21,33,40,41, is in the low range of the pathological O2 levels present in the inflamed synovium23,28,40, whereas 20% O2, a standard in vitro cell-culture condition33,40, is likely to represent a supraphysiological situation. Future studies will aim at assessing SF monocytic cell responses at intermediate O2 concentrations to mimic more closely the in vivo physiopathological O2 fluctuations in the JIA synovium.
VEGF and OPN promote angiogenesis by directly stimulating EC proliferation and survival, adhesion, and migration3,9,45. However, their proangiogenic activity is also exerted indirectly through the control of leukocyte trafficking10,12,13,15,46,47. VEGF is a potent chemoattractant for monocytic cells10,47, and OPN acts as an adhesive, chemotactic, and activating factor for monocytic cells and T lymphocytes, promoting their recruitment into the inflamed synovium and stimulating proinflammatory cytokine production12–16. The importance of these factors in the pathogenesis of inflammatory synovitis is emphasized by the observations that inhibition of VEGF-R1 signaling47,48 or OPN production11,49 is associated with marked reduction of joint inflammation in murine RA models. Previous studies in mouse models of arthritis have suggested a critical role for synovial monocytic cells in the regulation of arthritic joint inflammation, because their local removal from the knee joints by intraarticular administration of selective depleting agents, such as clodronate-encapsulated liposomes50, prior to induction of experimental arthritis results in a substantial decrease of influx of inflammatory cells and arthritis severity51,52. Thus, we can hypothesize that sustained VEGF and OPN induction by intraarticular hypoxia in synovial monocytic cells may represent an important mechanism of amplification of the inflammatory process through the recruitment of other inflammatory cells, which in turn will enhance the inflammatory response in the arthritic joints, ultimately leading to the chronicity of JIA synovitis.
Angiogenesis is a complex process, involving multiple gene products expressed by different cell types, all contributing to an integrated sequence of events. Elevated levels of other proangiogenic factors in addition to VEGF and OPN, such as interleukin 8 (IL-8), have been detected in synovial fluids from patients with JIA4,53 and may contribute to the pathogenesis of the disease. Interestingly, IL-8 is inducible by hypoxia in monocytic cells54, and a positive feedback loop among IL-8,VEGF, and OPN production was previously established. VEGF was reported to stimulate endothelial cells to produce both IL-855 and OPN9,14, and OPN was shown to trigger a proangiogenic phenotype in human monocytes by inducing IL-8 expression both in vitro and in vivo12,14. On the other hand, IL-8 can increase VEGF production56,57 and enhance VEGF-mediated neovascularization by concomitantly triggering EC proliferation and migration58. Hence, it is conceivable that the proangiogenic state of monocytic cells within the local hypoxic environment of inflamed joints will ultimately be dictated by the crosstalk among these mediators, which synergistically stimulate each other’s production and function as 2-way signaling molecules between monocytic cells and EC. Further investigations are under way in our laboratory to define the functional cooperation among these factors in JIA.
Our findings point to a role for hypoxia and the HIF-1 pathway in the persistence and amplification of synovial inflammation associated with JIA and as potential pharmacological targets for treatment of the disease.
Footnotes
-
Supported by grants from the Ministero Istruzione Universita’ e Ricerca (MIUR-FIRB) and the Fondazione Italiana per la Lotta al Neuroblastoma.
- Accepted for publication December 9, 2008.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}