

Abstract
Objective. To determine serum concentrations of pentraxin 3 (PTX3) and its clinical associations in patients with systemic sclerosis (SSc).
Methods. Serum PTX3 levels from 45 patients with diffuse cutaneous SSc (dSSc), 46 with limited cutaneous SSc (lSSc), and 20 healthy controls were examined by ELISA. PTX3 expression in the sclerotic skin from SSc patients was evaluated immunohistochemically. Normal and SSc fibroblasts were cultured and PTX3 levels in the culture medium were also examined by ELISA.
Results. Serum PTX3 levels were elevated in patients with SSc relative to controls. PTX3 levels in dSSc patients were significantly higher than in controls and lSSc patients. PTX3 expression in the sclerotic skin from SSc patients was more intense relative to normal skin. Elevation of serum PTX3 levels was associated with more frequent presence of pulmonary fibrosis, cardiac disease, and pitting scar/ulcer and increased serum immunoglobulin levels and erythrocyte sedimentation rates. PTX3 levels correlated positively with modified Rodnan total skin thickness score, and negatively with percentage vital capacity and percentage DLCO in patients with SSc. PTX3 levels also correlated positively with serum levels of 8-isoprostane, a marker of oxidative stress, and hyaluronan, recently identified as an endogenous ligand for Toll-like receptors. PTX3 production from cultured SSc fibroblasts was increased by stimulation with hyaluronan.
Conclusion These results suggest that elevated serum PTX3 levels are associated with the disease severity of SSc.
Systemic sclerosis (SSc) is a connective tissue disease that is characterized by vascular damage, excessive accumulation of extracellular matrix, and fibrosis in the skin and other internal organs, in the setting of an autoimmune background1. Although the pathogenesis of SSc remains unknown, it has been proposed that oxidative stress, microvasculature, and the immune system play an important role in disease development2–4. Altered endothelial cell function and blood vessel reactivity (Raynaud’s phenomenon) are the earliest disease manifestations, and are accompanied by the presence of proinflammatory cytokines and the loss of redox control, leading to oxidative stress5. Indeed, serum levels of 8-isoprostane, a marker of oxidative stress, are elevated in patients with SSc and are correlated with disease severity6. Further, enhanced production of reactive oxygen species degrades hyaluronan, which is a major glycosaminoglycan component of the extracellular matrix and is identified as an endogenous ligand for Toll-like receptors (TLR), leading to production of hyaluronan fragments that induce inflammatory responses7. In addition, vascular changes are frequently associated with stimulation of both innate and adaptive immune responses, resulting in B cell and T cell activation, and in many cases, autoantibody production5. Thus, it would be important to identify and characterize the mediators affecting the vascular damage and tissue remodeling that lead to features typical of SSc.
Pentraxin 3 (PTX3) is a recently characterized member of the long-pentraxin superfamily8,9. It shares the C-terminal pentraxin domain with short-pentraxins such as C-reactive protein (CRP) and serum amyloid P component, but differs for the presence of an unrelated long N-terminal domain, cellular source, and regulatory mechanism: short-pentraxins are produced in the liver, mostly in response to inflammatory mediators, most prominently interleukin 6 (IL-6)10,11. By contrast, PTX3 is synthesized locally at the inflammatory site from various kinds of cells, including endothelial cells8, smooth muscle cells8, and fibroblasts9. Its production is induced by inflammatory mediators such as lipopolysaccharide (LPS), IL-1ß, and tumor necrosis factor-α (TNF-α), but not IL-612,13. In addition, PTX3 binds to C1q, by which action PTX3 can both inhibit and activate the classical complement pathway, depending on the way it is presented14. It also binds to discrete membrane domains of apoptotic cells and thereby inhibits their recognition by dendritic cells15. Thus, PTX3 plays an important role in the regulation of inflammation and innate immunity, and possibly in autoimmunity13,16,17. Consistent with this, levels of PTX3 are elevated and are correlated with the disease activity or severity in various inflammatory, cardiovascular, and autoimmune diseases18–24. Recently, Luchetti, et al reported that SSc fibroblasts constitutively express PTX3 in the absence of deliberate stimulation24. However, the clinical correlation of PTX3 in patients with SSc remained unknown in that study. We investigated clinical correlation of serum PTX3 concentrations with immunological measures, the extent of skin fibrosis and vascular damage, and the presence of various organ involvements in patients with SSc.
MATERIALS AND METHODS
Serum samples
Serum samples were obtained from 91 Japanese patients with SSc (81 women, 10 men). All patients fulfilled the criteria proposed by the American College of Rheumatology25. Patients were grouped according to the classification system proposed by LeRoy, et al26: 46 patients (44 women, 2 men) had limited cutaneous SSc (lSSc) and 45 (37 women, 8 men) had diffuse cutaneous SSc (dSSc). The age of patients (mean ± SD) was 47 ± 16 years. Patients with dSSc were aged 48 ± 18 years, those with lSSc 45 ± 17 years. The disease duration of patients with lSSc was 8.5 ± 9.5 years and with dSSc 3.1 ± 2.5 years. No patient was treated with oral corticosteroid, D-penicillamine, or other immunosuppressive therapy at the evaluation. Antinuclear antibody (ANA) was determined by indirect immunofluorescence using HEp-2 cells as substrate, and autoantibody specificities were further assessed by ELISA and immunoprecipitation. Anticentromere antibody was positive in 38 patients (1 dSSc and 37 lSSc), anti-topoisomerase I antibody in 36 (31 dSSc, 5 lSSc), anti-U1RNP antibody in 4 (all lSSc), anti-U3RNP antibody in 2 (dSSc), and anti-RNA polymerases I and III antibody in 9 (all dSSc). The remaining 2 patients were negative for autoantibodies. Twenty age and sex-matched healthy Japanese individuals were employed as normal controls. Smokers were excluded from the study. Fresh venous blood samples were centrifuged shortly after clot formation. All samples were stored at −70°C before use.
Clinical assessment
Complete medical histories, physical examinations, and laboratory tests, including vital capacity (VC) and diffusion capacity for carbon monoxide (DLCO), were conducted for all patients. Skin score was measured by the technique of the modified Rodnan total skin thickness score (modified Rodnan TSS) as described27: anatomical areas were rated as 0 (normal skin thickness), 1+ (mild but definite thickening), 2+ (moderate skin thickening), and 3+ (severe skin thickening), and the modified Rodnan TSS was derived by summing the score from all 17 areas (range 0–51).
Organ involvement was defined as described with some modifications28: pulmonary fibrosis = bibasilar fibrosis on chest radiography and high resolution computed tomography; isolated pulmonary hypertension = clinical evidence of pulmonary hypertension and increased systolic pulmonary arterial pressure (> 35 mm Hg) by Doppler echocardiography, in the absence of severe pulmonary interstitial fibrosis; esophagus = hypomotility shown by barium radiography; joints = inflammatory polyarthralgias or arthritis; heart = pericarditis, congestive heart failure, or arrhythmias requiring treatment; kidney = malignant hypertension and rapidly progressive renal failure with no other explanation; and muscle = proximal muscle weakness and elevated serum creatine kinase.
The study protocol was approved by Kanazawa University Graduate School of Medical Science and Kanazawa University Hospital, and informed consent was obtained from all patients.
ELISA
Serum PTX3 levels were measured using specific ELISA kits (R&D Systems, Minneapolis, MN, USA), according to the manufacturer’s protocol. Each sample was tested in duplicate. The detection limit of this assay was 0.1 ng/ml. Serum levels greater than the mean + 2 SD of normal controls were considered to be elevated.
ELISA for serum 8-isoprostane levels and hyaluronan were performed as described6,29 using specific ELISA kits (Cayman Chemical Co., Ann Arbor, MI, and Echelon Biosciences Inc., Salt Lake City, UT, USA), according to the manufacturer’s protocol. Each sample was tested in duplicate. The detection limit of each assay was 5 pg/ml and 50 ng/ml, respectively.
Immunohistochemical staining for PTX3
Formalin-fixed and paraffin-embedded tissues obtained from 10 SSc patients (5 dSSc, 5 lSSc) and 5 controls were cut into 4 μm sections, deparaffinized in xylene, and rehydrated in phosphate buffered saline (PBS). Deparaffinized sections were preincubated with 1% H2O2 for 5 min to block tissue peroxidase activity. Then the sections were incubated with bovine serum albumin in PBS for 30 min at 37°C, followed by overnight incubation at 4°C with rabbit anti-human PTX3 antibody (Santa Cruz Biotechnology, Santa Cruz, CA, USA). Rabbit IgG (Southern Biotechnology Associates, Birmingham, AL, USA) was used as a control for nonspecific staining. After washing with PBS, the slides were incubated with biotinylated goat anti-rabbit immunoglobulin G secondary antibody (Vector Laboratories, Burlingame, CA, USA), then horseradish peroxidase-conjugated avidin-biotin complexes. Sections were developed with 3,3′-diaminobenzidine tetrahydrochloride and hydrogen peroxide, and then counterstained with methyl green.
Fibroblast culture
Human dermal fibroblasts were obtained by skin biopsy from 6 patients with SSc and 6 healthy individuals, with informed consent from all subjects. Primary explant cultures were established in 25-cm2 culture flasks in modified Eagle’s medium (MEM) containing 10% fetal calf serum. Cultures of fibroblasts were maintained as monolayers at 37°C in 5% CO2, and studied between the third and sixth subpassages. Fibroblasts were grown to confluence, and the culture medium was replaced with serum-free MEM. There was no difference in growth rate between SSc and normal fibroblasts. In addition, the number of cells was similar between SSc and normal fibroblasts when fibroblasts were grown to confluence (data not shown). After a further 24-h period of incubation, fibroblasts were cultured with LPS (0111:B4; Sigma-Aldrich, St. Louis, MO, USA) or low molecular weight hyaluronan (15–40 kDa; R&D Systems) for 24 h. The endotoxin level of hyaluronan used in this study was < 0.01 EU/mg, and treatments with hyaluronan were in the presence of 10 μg/ml of polymyxin B (Calbiochem, Darmstadt, Germany) to exclude the effects of any contaminating LPS on experimental conditions. After the incubation, PTX3 levels in the culture medium were examined by ELISA.
Statistical analysis
Statistical analysis was performed using the Mann-Whitney U test for determining the level of significance of differences between sample means, Fisher’s exact probability test for comparison of frequencies, and Bonferroni’s test for multiple comparisons. Spearman’s rank correlation coefficient was used to examine the relationship between 2 continuous variables. A p value less than 0.05 was considered statistically significant.
RESULTS
Serum PTX3 levels in SSc
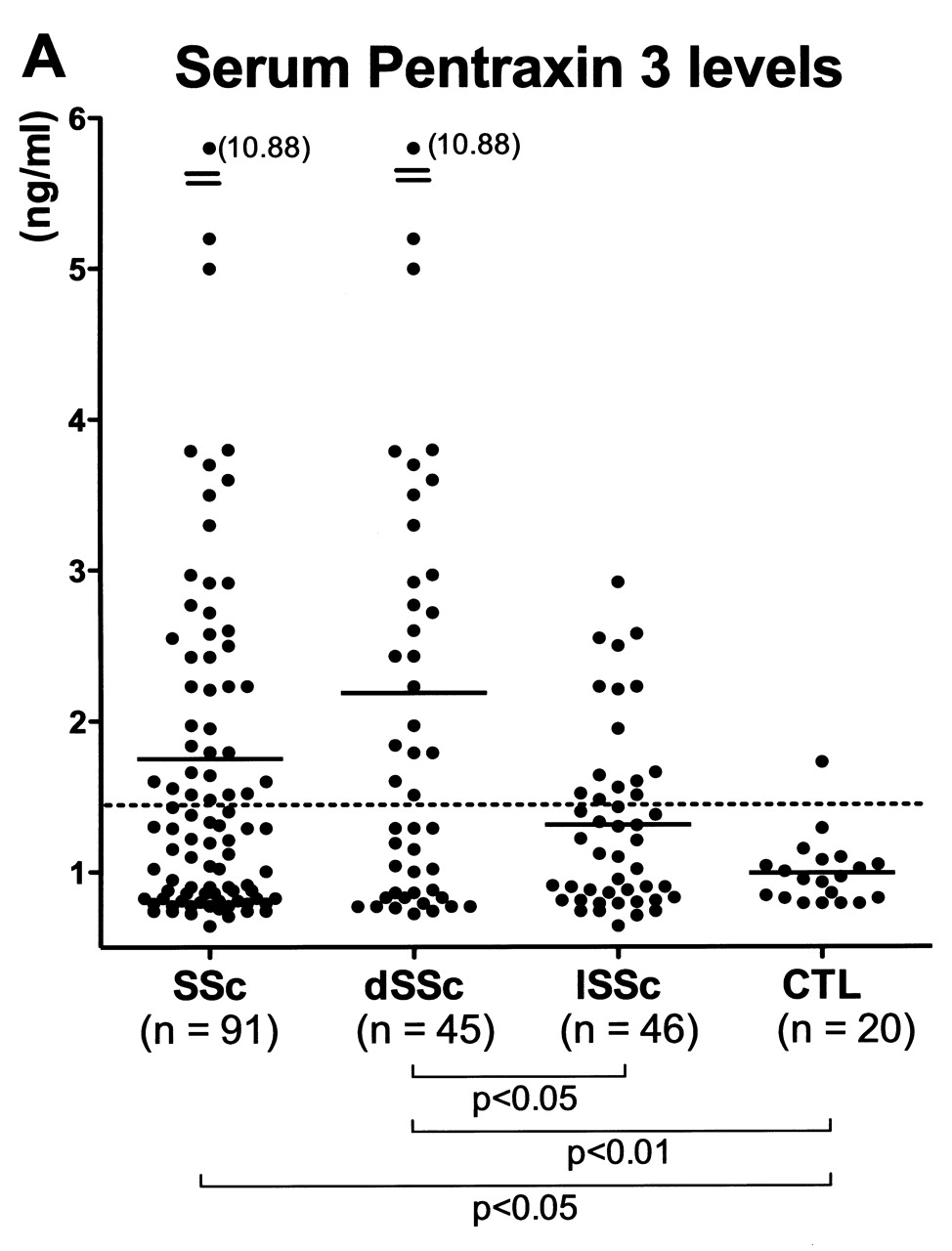
The levels of PTX3 in serum samples from SSc patients and controls were assessed by ELISA (Figure 1A). Serum PTX3 levels were significantly elevated in patients with SSc (mean 1.76 ng/ml, range 0.64–10.88) compared with controls (mean 0.98 ng/ml, range 0.79–1.73; p < 0.05). For the SSc subgroups, PTX3 levels in patients with dSSc (mean 2.19 ng/ml, range 0.71–10.88) were higher than in those with lSSc (mean 1.31 ng/ml, range 0.64–2.92; p < 0.05) and controls (p < 0.01). By contrast, there was no significant difference in serum PTX3 levels between patients with lSSc and controls (p = 0.1). Values higher than the mean + 2 SD (1.44 ng/ml) of the control serum samples were considered to be elevated. Elevated PTX3 levels were observed in 42% (38/91) of all patients with SSc, in 51% (23/45) of patients with dSSc, and in 33% (15/46) of patients with lSSc. By contrast, only 5% (1/20) of controls had elevated PTX3 levels. Thus, serum PTX3 levels were elevated in SSc, especially in dSSc patients.

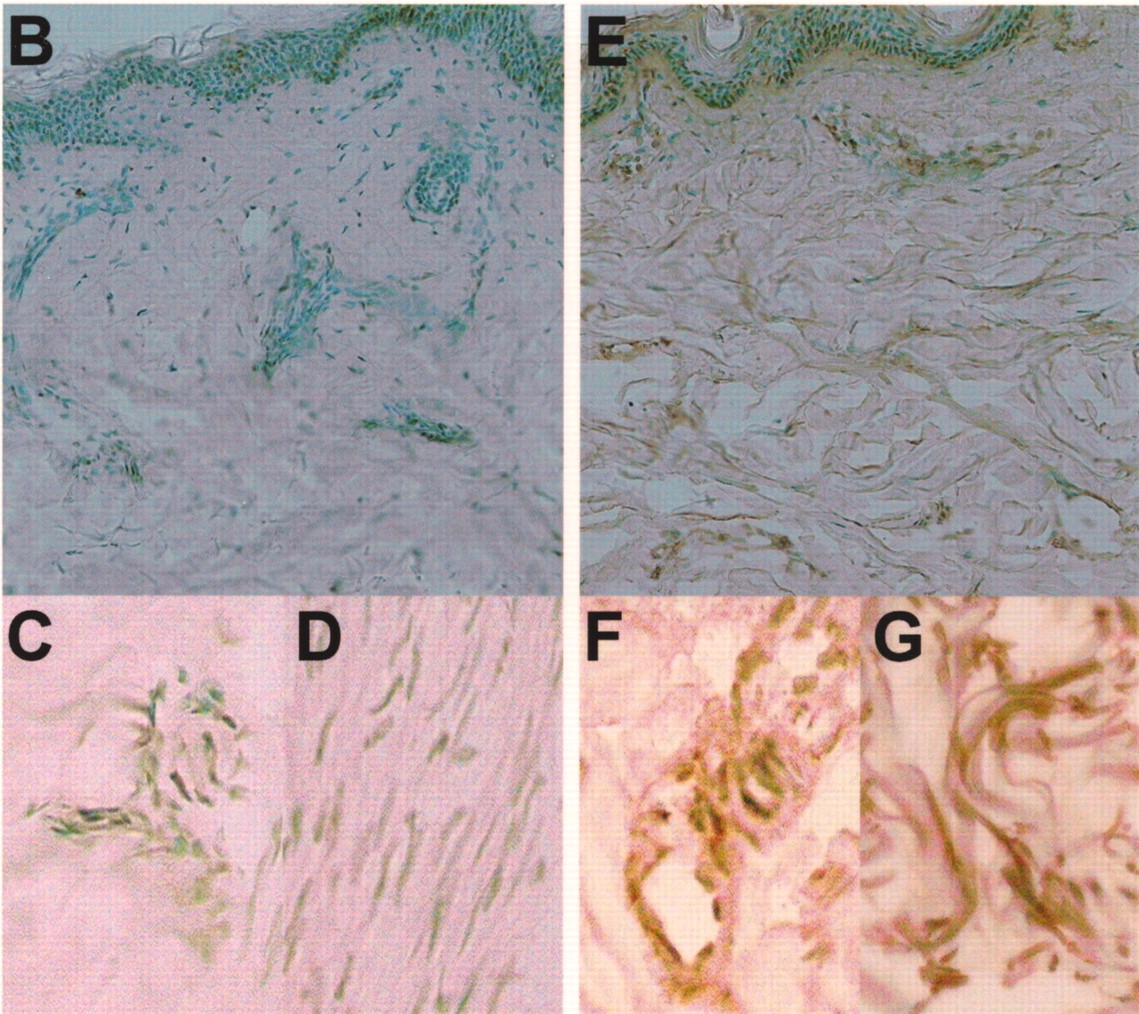
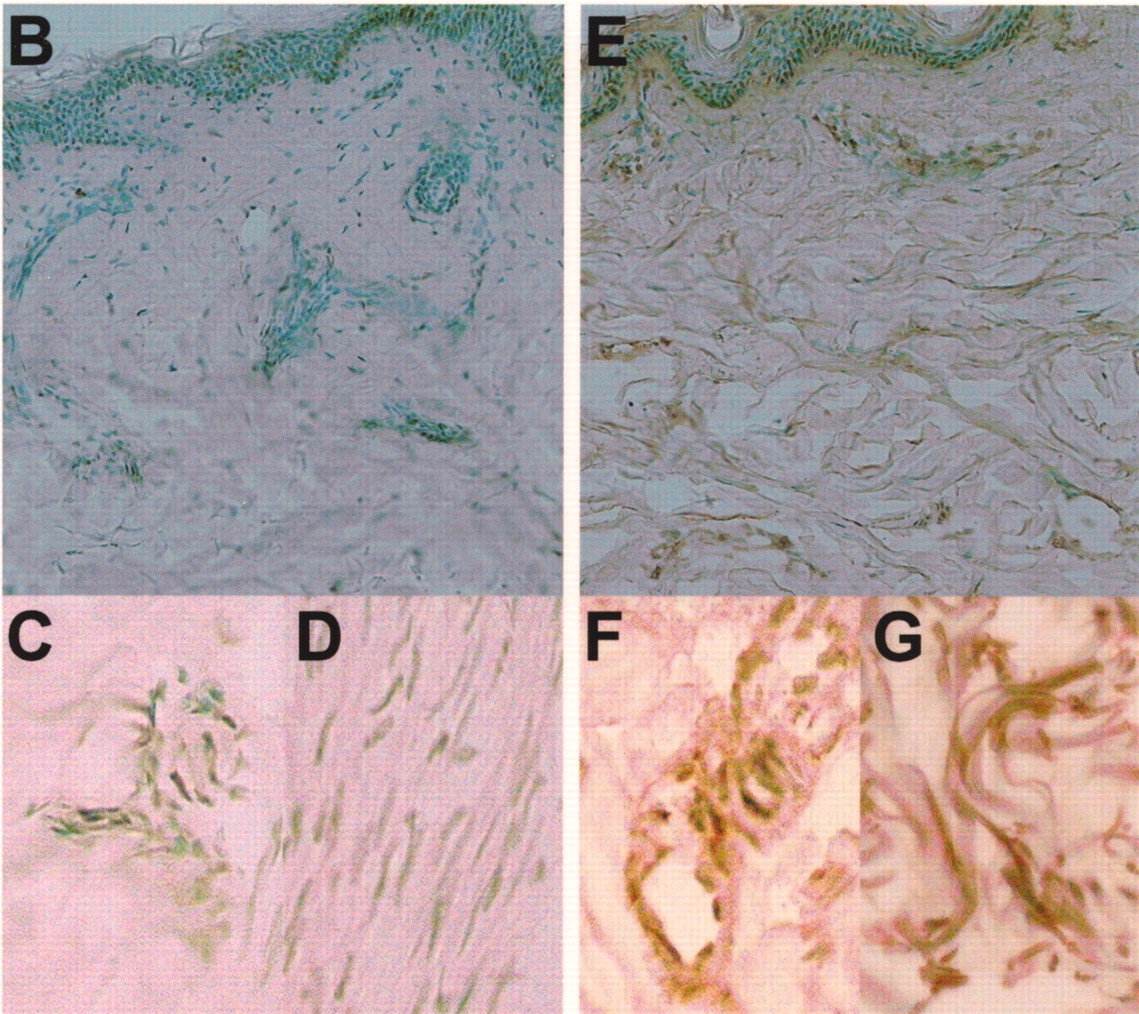
A. Serum levels of pentraxin 3 (PTX3) in patients with SSc, dSSc, or lSSc and healthy controls (CTL). Serum PTX3 levels were determined by a specific ELISA. Short bars indicate mean value in each group. Broken line indicates the cutoff value (mean + 2 SD of the healthy control samples). B–G. Representative PTX3 expression in normal skin and sclerotic skin from a patient with SSc. PTX3 expression was only faintly detected in the basal layer of normal skin (B), and was not observed in endothelial cells (C) and fibroblasts (D). Fibrotic skin from SSc patients had higher expression of PTX3 in the basal layer, endothelial cells (F), and fibroblasts (G) compared with normal skin. Original magnification: B and E ×100; C, D, F, G ×400.
PTX3 expression in fibrotic skin
PTX3 expression was detected only faintly in the basal layer of normal skin (Figure 1B), and was not observed in endothelial cells (Figure 1C) and fibroblasts (Figure 1D). By contrast, fibrotic skin from SSc patients had higher expression of PTX3 in the basal layer, in endothelial cells (Figure 1F), and in fibroblasts (Figure 1G) compared with normal skin. PTX3 expression in sclerotic skin of lSSc patients was similar to that of dSSc patients (data not shown).
Clinical correlations
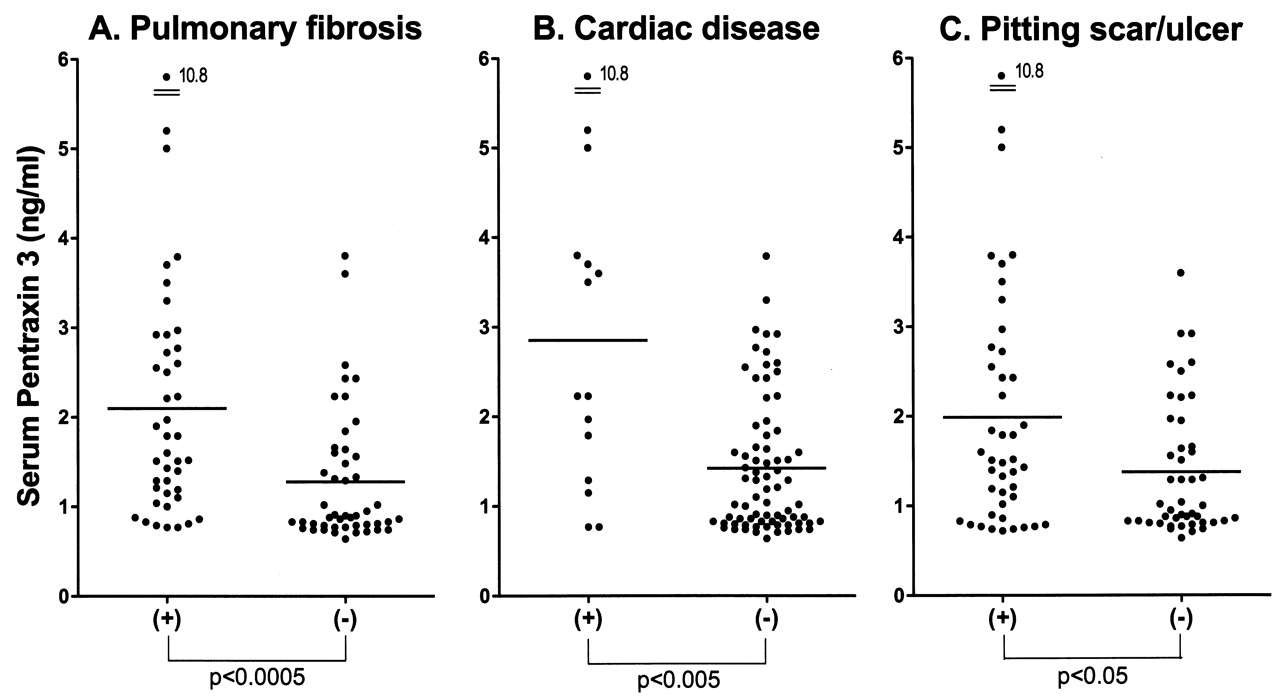
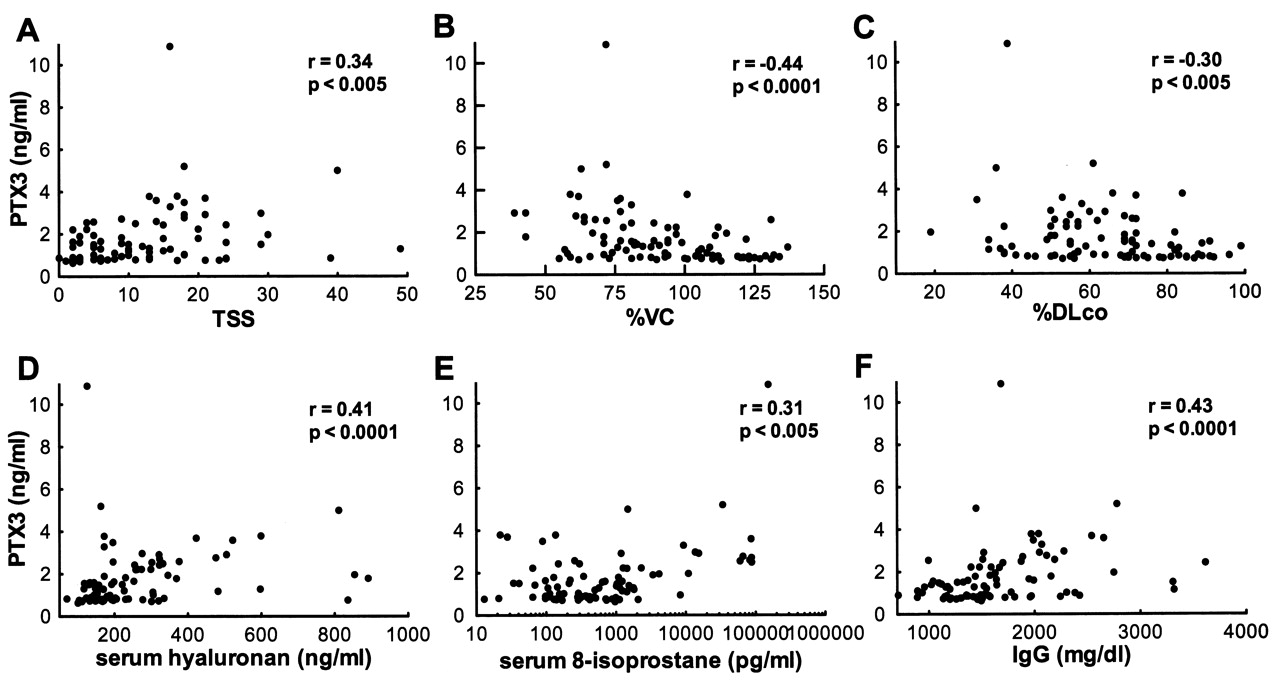
We assessed clinical correlations of serum PTX3 levels in SSc patients. Patients with elevated serum PTX3 levels had significantly higher modified Rodnan TSS (p < 0.05) and more frequent presence of contracture of phalanges (p < 0.01), pulmonary fibrosis (p < 0.01), and cardiac disease (p < 0.05) than those with normal PTX3 levels (Table 1). Consistent with this, PTX3 levels were significantly higher in SSc patients with pulmonary fibrosis (p < 0.0005; Figure 2A), cardiac disease (p < 0.005; Figure 2B), and pitting scar/ulcer (p < 0.05; Figure 2C) than in those without. When we analyzed dSSc patients (n = 45), PTX3 levels were also significantly higher in patients with pulmonary fibrosis (p < 0.01), cardiac disease (p < 0.05), and pitting scar/ulcer (p < 0.01). In addition, SSc patients with elevated serum PTX3 levels had decreased %VC (p < 0.05) and %DLCO (p < 0.05) compared to those with normal PTX3 levels (Table 1). Further, serum PTX3 levels correlated positively with modified Rodnan TSS (r = 0.34, p < 0.005; Figure 3A), and inversely with %VC (r = −0.44, p < 0.0001; Figure 3B) and %DLCO (r = −0.30, p < 0.005; Figure 3C). PTX3 levels also correlated with serum hyaluronan levels (r = 0.41, p < 0.0001; Figure 3D) and 8-isoprostane (r = 0.31, p < 0.005; Figure 3E).

Serum levels of pentraxin 3 (PTX3) in the presence and absence of pulmonary fibrosis (A), cardiac disease (B), and pitting scar/ulcer (C) in patients with SSc. Serum PTX3 levels were determined by ELISA. Short bars indicate mean value in each group.
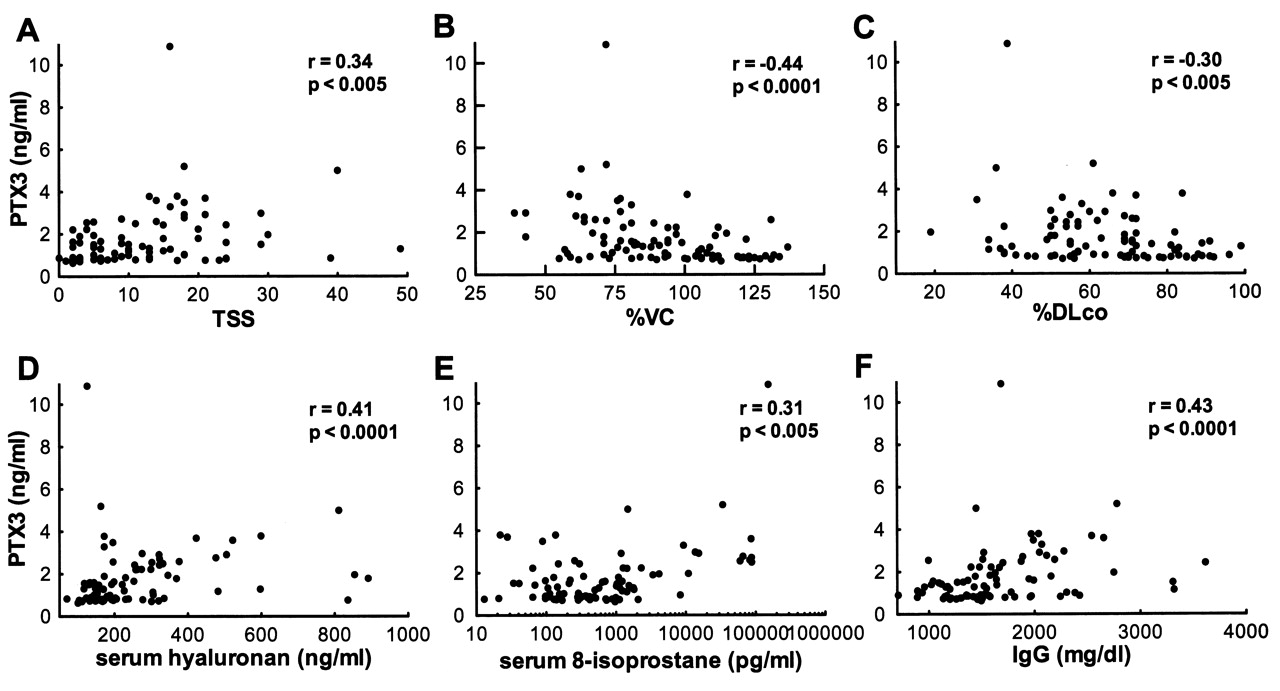
Correlation of pentraxin 3 (PTX3) levels with modified Rodnan total skin thickness score (TSS; A), %VC (B), %DLCO (C), serum hyaluronan levels (D), serum 8-isoprostane levels (E), and serum IgG levels (F) in patients with SSc.
Clinical and laboratory data of patients with SSc showing elevated serum PTX3 levels. Unless noted otherwise, values are percentages.
As for correlations of PTX3 levels with immunological measures, SSc patients with elevated PTX3 levels had significantly higher levels of serum IgG and IgA and erythrocyte sedimentation rates (ESR) than those with normal levels (p < 0.01, p < 0.05, and p < 0.05, respectively; Table 1). Moreover, serum PTX3 levels also correlated positively with levels of serum IgG (r = 0.43, p < 0.0001; Figure 3F) and IgA (r = 0.30, p < 0.005), and ESR (r = 0.37, p < 0.01). Anti-topoisomerase I antibody was detected more frequently in SSc patients with elevated PTX3 levels than in those with normal levels (p < 0.05; Table 1). However, PTX3 levels did not correlate with serum levels of anticentromere antibody, anti-RNP antibody, or CRP (data not shown). Thus, the elevation of PTX3 was associated with the extent of skin sclerosis, the presence and severity of pulmonary fibrosis, the presence of cardiac disease and pitting scar/ulcer, and serum levels of immunoglobulin, ESR, hyaluronan and 8-isoprostane in SSc.
PTX3 production by cultured SSc fibroblasts
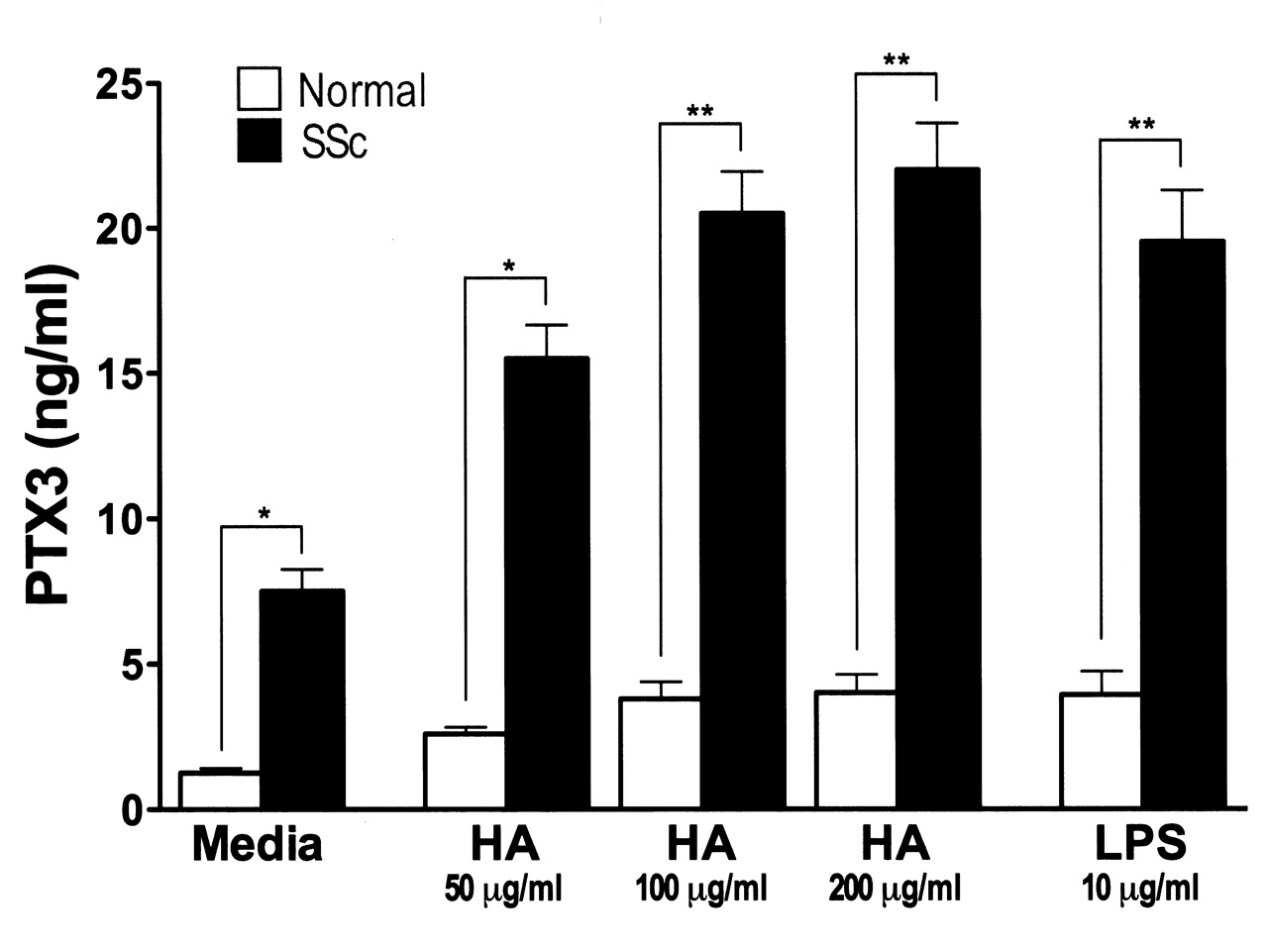
PTX3 is produced from various cells, including fibroblasts, by LPS stimulation30–32. Studies have shown that breakdown products of the extracellular matrix, especially hyaluronan, stimulate TLR as endogenous ligands for TLR, and thereby regulate inflammatory responses, including autoimmune disorders33–35. Indeed, hyaluronan expression is enhanced in fibrotic skin from patients with SSc36. Therefore, we assessed whether PTX3 production by fibroblasts was increased by hyaluronan (Figure 4). Unstimulated SSc fibroblasts produced higher levels of PTX3 relative to normal fibroblasts (p < 0.05). When fibroblasts were stimulated with LPS or hyaluronan, PTX3 production from both SSc and normal fibroblasts was significantly increased (p < 0.05). In addition, PTX3 levels by LPS or hyaluronan-stimulated SSc fibroblasts were higher than those of normal fibroblasts (p < 0.01). The minimal effective dose of hyaluronan was 50 μg/ml. Thus, PTX3 production from SSc fibroblasts was increased by hyaluronan stimulation in vitro.
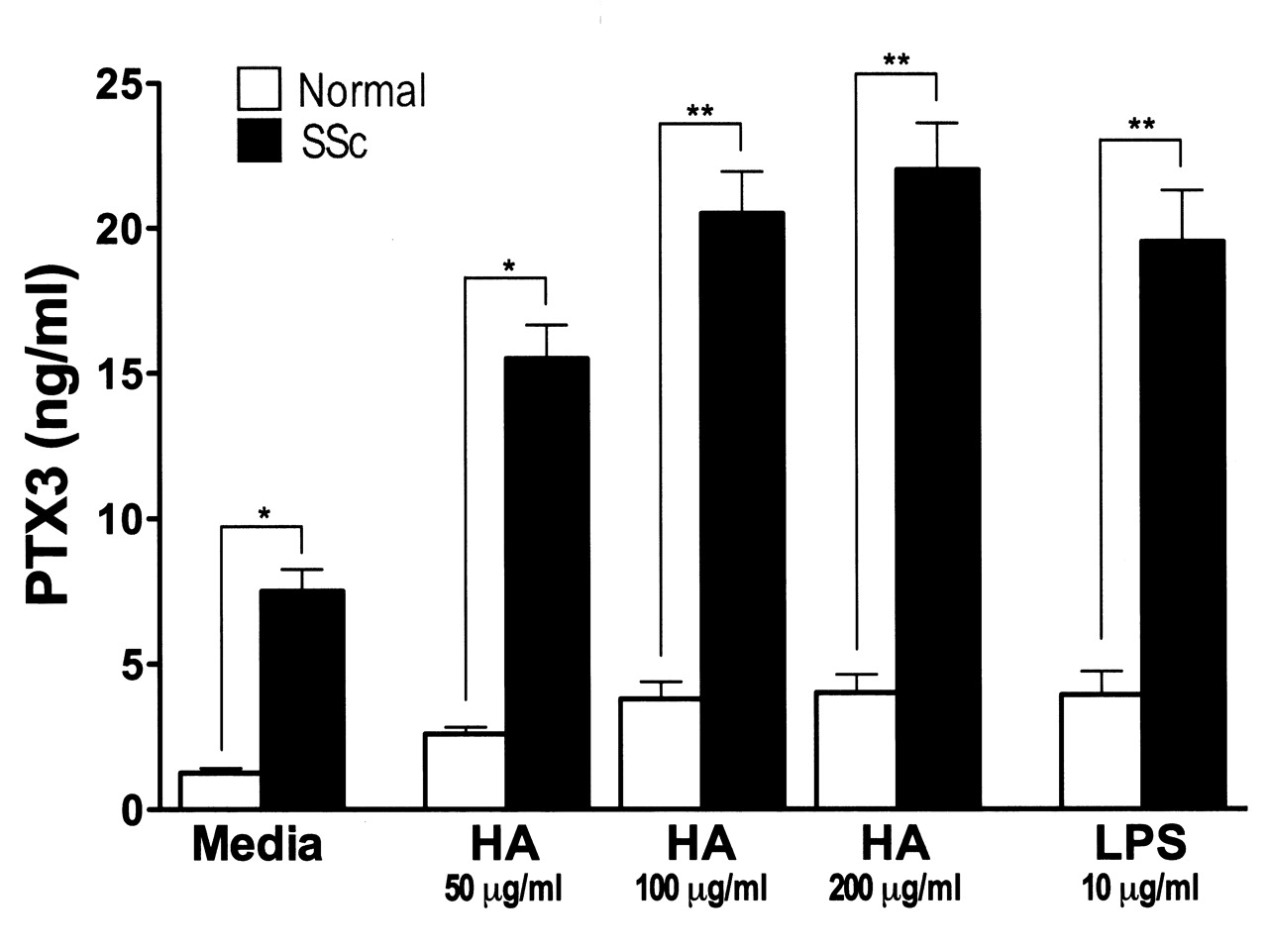
Pentraxin 3 (PTX3) production from LPS- or hyaluronan (HA)-stimulated normal (n = 6) and SSc fibroblasts (n = 6). Fibroblasts were grown to confluence and culture medium was replaced with serum-free MEM. After further incubation for 24 hours, fibroblasts were stimulated with LPS (10 mg/ml) or low molecular weight hyaluronan (50, 100, 200 mg/ml) for 24 hours. After incubation, PTX3 levels in the supernatants were analyzed by ELISA. Each histogram shows mean level (± SEM). *p < 0.05, **p < 0.01
DISCUSSION
Our study is the first to find that serum PTX3 levels were elevated in patients with SSc. Serum PTX3 levels were correlated with the extent of skin fibrosis and the severity of pulmonary fibrosis. Consistent with this, PTX3 expression in the fibrotic skin from SSc patients was enhanced relative to normal skin, by immunohistochemical analysis. Since PTX3 expression in sclerotic skin of lSSc patients was similar to that of dSSc patients, the difference of the serum PTX3 levels between dSSc and lSSc may reflect the accumulation of PTX3-producing fibroblasts in sclerotic skin and lung. In addition, elevation of serum PTX3 was also associated with higher frequency of cardiac disease and contracture of phalanges. Moreover, PTX3 levels were significantly elevated in SSc patients with pitting scar/ulcer and cardiac disease. Thus, serum PTX3 levels were associated with vascular damage as well as the extent of fibrosis in skin and lung, which are the main features in SSc. However, the percentages of dSSc patients with elevated and normal PTX3 levels were 61% (23/38) compared to 42% (22/53), respectively; this difference did not reach statistical significance (p = 0.09). The reason for this discrepancy may be due to the heterogeneity of SSc in the extent of skin or lung fibrosis. Indeed, when we analyzed dSSc patients, PTX3 levels were significantly higher in those with pulmonary fibrosis, cardiac disease, and pitting scar/ulcer, indicating that patients with “mild” dSSc may have normal PTX3 levels. Together, these results suggest that PTX3 is a useful serological marker for evaluating disease severity in SSc.
A previous study reported that the mean serum level of PTX3 in 30 patients with SSc was not significantly different from that of healthy controls24. Since clinical information (the ratio of dSSc and lSSc, extent of skin sclerosis, presence and severity of visceral involvement such as pulmonary fibrosis, etc.) was not given in that study, the severity of SSc disease in the patients is unknown. In our study, lSSc patients had only slight elevations of serum PTX3 compared with healthy controls (p = 0.36). By contrast, dSSc patients had significant elevation of serum PTX3 relative to controls (p < 0.005). Further, SSc patients with pulmonary fibrosis showed higher levels of serum PTX3 than those without pulmonary fibrosis. Thus, the discrepancy between the previous study24 and ours may be the result of the difference of patients’ disease severity. Alternatively, it may result from the differences of measuring systems, genetic background of patients, patient numbers, or other unknown reasons.
Since PTX3 is synthesized by various kinds of cells, including endothelial cells8, smooth muscle cells8, and fibroblasts9, increased serum PTX3 is not specific for SSc. Indeed, serum PTX3 levels were elevated in patients with other inflammatory and autoimmune disorders, especially in those with vascular diseases18–24. It was reported that the main sites of PTX3 expression in patients with vasculitis were endothelial cells and infiltrating leukocytes22,37. By contrast, elevation of PTX3 may reflect the overproduction from fibroblasts in SSc: PTX3 staining was observed in dermal fibroblasts and endothelial cells in SSc (Figure 1), and this is consistent with the previous report24. In addition, SSc fibroblasts constitutively overexpressed PTX3 relative to normal fibroblasts (Figure 4), and serum PTX3 levels were correlated with the extent of skin and lung fibrosis. Therefore, the difference of the main sites of PTX3 expression indicates that the mechanisms responsible for elevated serum PTX3 levels are different between vascular diseases and SSc.
In our study, serum PTX3 levels were found to be correlated with serum 8-isoprostane, suggesting that elevated PTX3 is associated with enhanced oxidative stress. It is possible that enhanced oxidative stress, which may reflect vascular damage due to ischemia and reperfusion injury, leads to the production of PTX3 from damaged endothelial or vascular cells. Substantial evidence shows that reactive oxygen species are responsible for hyaluronan degradation, leading to production of hyaluronan fragments that induce inflammatory responses7. Indeed, in a bleomycin-induced pulmonary fibrosis model, hyaluronan degradation products, which are generated after tissue injury and inflammation, stimulated macrophages to produce inflammatory mediators in a TLR4 and TLR2-dependent manner38. Further, serum hyaluronan levels are elevated and are correlated with the disease severity in SSc, and hyaluronan expression is enhanced in the fibrotic skin from patients with SSc36,39. Remarkably, PTX3 production from SSc fibroblasts was increased by stimulation with low molecular weight hyaluronan in vitro. In addition to TLR4, since fibroblasts express the hyaluronan receptor CD4440, it is possible that a CD44-mediated signaling pathway may affect PTX3 production by hyaluronan-stimulated fibroblasts. It was reported that CD44 expression is significantly decreased in dSSc fibroblasts compared with normal and lSSc fibroblasts40. However, there was no significant difference in PTX3 production between dSSc and lSSc fibroblasts (data not shown). Therefore, PTX3 production by hyaluronan-stimulated SSc fibroblasts may presumably be mediated by a TLR4 signaling pathway. Together, these results indicate that elevated serum PTX3 in SSc may result from PTX3 release from damaged endothelial cells and overproduction from fibroblasts induced by hyaluronan. To test this hypothesis, we must perform further studies using animal models of SSc or in vitro studies (i.e., downstream signaling and transcription events of hyaluronan-stimulated SSc fibroblasts).
In our study, serum PTX3 levels were correlated with serum immunoglobulin levels and ESR, but PTX3 levels were not correlated with CRP levels. This may be due to the difference of regulatory mechanisms: while CRP is produced in the liver by inflammatory mediators, mainly IL-6, PTX3 is produced at the site of inflammation by mediators such as TNF-α, not IL-612,13. It has been reported that reperfusion-induced injury involves the recruitment of activated leukocytes, especially neutrophils, and the release of soluble mediators of inflammation, notably TNF-α41. Further, after ischemia and reperfusion injury, PTX3 transgenic mice have an impaired survival rate, with a markedly increased inflammatory response and elevated concentration of TNF-α compared to wild-type mice41. Thus, it seems that a TNF-α/PTX3 loop is functioning in vivo, and it appears to amplify the inflammatory response41. Since ischemia and reperfusion are early and typical events of SSc, our results and previous reports suggest that PTX3 may play a role in the immunological abnormalities associated with SSc by amplifying inflammation.
Although the function of PTX3 has not been completely defined, it is thought to play important roles in the regulation of inflammation and angiogenesis10,11,17,42,43. PTX3 can bind to the complement fraction C1q, resulting in activation of the classic complement pathway14. The complement system has been shown to be involved in the pathogenesis of SSc; several studies have reported abnormal complement activation and subendothelial deposition of immune complexes in patients with SSc44,45. A complement system activated by increased PTX3 may impair the membranes of endothelial cells, resulting in endothelial cell death. In addition, PTX3 also binds basic fibroblast growth factor with high affinity and selectivity, leading to inhibition of its angiogenic activity in vitro and in vivo42,43. PTX3 may have a role in the development of SSc by amplifying inflammation and inhibiting angiogenesis.
Our study reveals that serum PTX3 concentrations are elevated in SSc and are associated with the extent of skin fibrosis, with the presence and severity of pulmonary fibrosis, and with vascular damage. Our results indicate that elevated serum PTX3 may result from overproduction by hyaluronan-stimulated fibroblasts and/or release from damaged cells due to enhanced oxidative stress. Although further studies are required to clarify the role of PTX3 in the development of SSc, it may be a useful serologic marker for the disease severity of SSc.
Acknowledgments
We thank A. Usui, M. Yozaki, and K. Shimoda for technical assistance.
Footnotes
-
Supported by a grant for Research on Intractable Diseases from the Ministry of Health, Labour and Welfare of Japan.
- Accepted for publication December 17, 2008.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}