
Abstract
Objective. Minor salivary gland specimens were analyzed to investigate dysregulation of the proteasome system in patients with Sjögren’s syndrome (SS) and patients with sicca syndrome.
Methods. Labial biopsy specimens from 17 patients with SS and 11 patients with non-autoimmunesicca syndrome were analyzed by immunohistochemistry for expression of the inducible proteasomal subunits ß1i, ß2i, and ß5i. The infiltrating subsets of lymphocytes were characterized using immunofluorescence stainings against the cell-surface markers CD20 and CD27. Two-dimensional electrophoresis and immunoblotting were used for detection of the proteasomal subunits ß1 and ß1i in peripheral blood monocyte cells. Gene expression of the constitutive subunits ß1, ß2, and ß5 and the corresponding inducible subunits ß1i, ß2i, and ß5i was further investigated at the mRNA level in small lip biopsies using real-time polymerase chain reaction.
Results. The expression of ß1i in infiltrating and peripheral immune cells was altered in patients with SS compared to patients with non-autoimmune sicca syndrome and healthy controls. No significant differences were found in ß2i and ß5i expression between the same groups in small lip biopsies. Chisholm-Mason grade and ß1i expression were found to be inversely correlated (Spearman r = −0.461, p = 0.014). The phenotype and distribution of the lymphocytic infiltrate showed no differences between patients with primary and secondary SS regardless of ß1i expression.
Conclusion. The proteasomal ß1i subunit is dysregulated in peripheral white blood cells and in inflammatory infiltrates of minor salivary glands in patients with SS.
- PROTEASOME SYSTEM
- SJÖGREN’S SYNDROME
- ß1i EXPRESSION
- SALIVARY GLANDS
Sjögren’s syndrome (SS) is a systemic autoimmune disorder characterized by dysfunction and inflammatory destruction of exocrine glands, leading to the manifestation of sicca symptoms1,2. Involvement of salivary glands represents a common feature of several autoimmune diseases, implicating a susceptibility of the exocrine epithelium for the development and maintenance of an autoimmune response. This phenomenon could be attributed to a preferred exposure to environmental factors such as recurrent viral infections or to intrinsic factors, e.g., altered antigen presentation.
In the minor salivary glands of patients with SS, infiltrating macrophages, B and T cells, and ductal epithelium release distinct chemokines and cytokines, including type 1 interferons3⇓–5. This proinflammatory milieu can alter basic cellular functions and might also cause structural and functional changes in the ubiquitin-proteasome system. The proteasome is not only an essential degradation apparatus for cellular homeostasis, but is also involved in regulation of apoptosis and the cell cycle and immune response6⇓–8. In eukaryotic cells, the cylindrical 20S core complex consists of 7 different alpha-type subunits forming 2 outer rings and 7 beta-type subunits bearing 3 active sites in both inner rings9. However, under the influence of proinflammatory cytokines such as interferon-γ or interferon-α, 3 different inducible proteolytic subunits, ß1i (LMP2), ß2i (MECL1), and ß5i (LMP7), are upregulated to replace their constitutive counterparts10⇓⇓⇓–14. By favoring the generation of distinct antigenic peptides for the MHC class I pathway, the so-called immuno-proteasomes are critical for an effective immune response12,14⇓⇓⇓–18. Thus, the proteasome is a multicatalytic protease with several active sites and regular structural modifications that have been shown to have a striking influence on the function of the entire complex.
Recently, a disease-specific dysregulation of the proteasome system was recognized in peripheral blood monocyte cells (PBMC) of patients with SS with altered expression of ß1i at the protein level19. The aim of our study was to investigate the expression of the inducible proteasome subunits in minor salivary glands of patients with SS and to clarify whether ß1i expression is also modified in the targeted tissue. In addition to standard histological examination of minor salivary gland biopsies by the Chisholm and Mason grading system, immunostaining of minor salivary glands and gene expression analysis were undertaken to compare the expression of the inducible proteasomal subunits at the protein and mRNA levels and to characterize the lymphocyte subsets20.
MATERIALS AND METHODS
Patients
Biopsies from minor salivary glands were obtained from 28 consecutive patients with sicca symptoms. All patients were treated at the Department of Rheumatology and Clinical Immunology, Charité–Universitätsmedizin Berlin. The study was performed after approval by the local ethical committee, and informed consent was obtained from all donors before lip biopsies were taken.
Primary SS was diagnosed in 14 patients according to the US-European classification criteria21 (all female, mean age 50.4 yrs, range 18–70 yrs). Secondary SS was diagnosed in 2 patients with primary biliary cirrhosis (PBC) and in one patient with undifferentiated connective tissue disease (UCTD) (all female, mean age 42.7 yrs, range 28–51 yrs). Sicca symptoms without an autoimmune sialadenitis were confirmed in 11 patients (9 female, 2 male, mean age 54.7 yrs, range 27–73 yrs; 7 with no autoimmune disorder, 2 with Hashimoto’s thyroiditis, one with UCTD, one with PBC).
At the time of the biopsy, 4 of 14 patients with primary SS were receiving immunomodulating therapy (hydroxychloroquine, n = 2; hydroxychloroquine plus prednisolone, n = 2). One of the patients with sicca symptoms was taking prednisolone.
Immunohistochemistry
Paraffin-embedded sections (1–3 μm) of minor labial gland biopsy tissues were deparaffinized and rehydrated. After antigen retrieval (pressure cooker, citrate buffer, pH 6) rabbit anti-human polyclonal antibodies anti-ß1i (K463 and K464), anti-ß2i, and anti-ß5i22 were incubated in a dilution of 1:1000. Staining was done using the labeled streptavidin-biotin method (LSAB-Kit; Dako, Glostrup, Denmark). Fuchsin (Dako) was used as a chromogen for the reaction with alkaline phosphatase. In a final step, nuclear counterstaining was performed with hematoxylin. Nonspecific binding of the secondary antibody was investigated by omitting the primary antibody.
Intensity of staining reactions was assessed on a scale from 0 (no staining) to 3 (strong reaction) by 2 independent pathologists blinded to the diagnosis.
Immunofluorescence
Paraffin-embedded sections of minor labial gland biopsy tissues (1–2 μm) were deparaffinized and rehydrated. After antigen retrieval (pressure cooker, EDTA buffer, pH 8), proteins were blocked for 5 min in blocking solution (Zytomed Systems, Berlin, Germany). Next, rabbit polyclonal antibody anti-CD20 (dilution 1:50; Zytomed Systems) and the mouse monoclonal antibody anti-CD27 (clone 137B4, dilution 1:10; LabVision, Fremont, CA, USA) were incubated overnight at 4°C. The following steps were performed in darkness: Cy3-conjugated donkey anti-mouse antibody (dilution 1:200; Dianova, Hamburg, Germany) and Cy2-conjugated donkey anti-rabbit antibody (dilution 1:100; Dianova) were incubated for 30 min. Nuclear staining was performed using DAPI (dilution 1:2500; Fluka Chemie AG, Buchs, Switzerland). Slides were mounted with fluorescent mounting medium (Dako).
The slides were viewed on a Leitz DMRBE microscope (Leica, Wetzlar, Germany) with a fluorescent light application using filters for Cy2 (resulting in green fluorescent staining), Cy3 (red fluorescence), and DAPI (blue nuclear fluorescent staining). Slides were viewed under 50×, 100×, 200×, and 400× magnification; areas with inflammatory infiltrate were photographed successively using the 3 filters and an overlay was generated. Attention was given to potentially double-positive cells, which should appear yellow on the overlay pictures.
Isolation of PBMC
PBMC of 6 patients with SS and 3 healthy controls were isolated from 20 ml heparinized peripheral blood by density gradient centrifugation using Ficoll lymphocyte isolation solution (GE Healthcare Bio-Science, München, Germany). The cells’ interphase containing the PBMC was harvested and washed with phosphate buffered saline (PBS). Remaining erythrocytes were lysed using erythrocyte lysis buffer (Qiagen, Hilden, Germany) according to the instructions of the manufacturer.
Protein isolation
For protein isolation, the PBMC pellet was lysed in 500 μl solution of 9 M urea and 50 mM DTT by short sonication and incubation for 1 h at 30°C. After centrifugation, the supernatant was precipitated with TCA and redissolved in 100 μl solution of 9 M urea, 50 mM DTT, 2% Ampholyte, pH 7–9, to get a higher protein concentration. The protein concentration was determined by the BCA method according to the manufacturer’s instructions (Pierce, Thermo Fisher Scientific; Rockford, MA, USA).
Two-dimensional electrophoresis
Two-dimensional electrophoresis was performed on a mini-vertical gel electrophoresis unit SE 250 with SE 220 tube gel adapter (Hoefer Pharmacia Biotech, San Francisco, CA, USA).
For the first dimension, the tube gel used for isoelectric focusing was 1.5 mm × 70 mm. Samples from different individuals were normalized based on protein concentration. For each run, a volume of 10 μl sample corresponding to 100 μg of protein per gel tube was used.
For the second dimension, the gel used for sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) was 60 mm × 80 mm × 1.5 mm. A 15% gel was used to allow appropriate separation of the proteasome subunits.
Blotting and immune detection
Immunoblotting using 10 μg of total proteins derived from PBMC of 3 patients (SS) and 2 healthy donors as antigens was performed as described19. For detection of proteasomal subunit ß1i, 3 different polyclonal antibodies (K463, K464, and PW8345; Biomol, Hamburg, Germany) were used, each in a 1:1000 dilution.
K463 and K464 were generated against recombinant murine ß1i and PW8345 directed against a peptide sequence derived from human ß1i.
After 2-dimensional PAGE, samples were transferred by semidry blotting to nitrocellulose membranes (Schleicher & Schüll, Dassel, Germany). All subsequent incubation and washing steps were performed at room temperature. Unspecific binding sites were blocked with 1% skim milk powder in PBS and 0.1% Tween 20 for 1 h. Membranes were then incubated with a mixture of polyconal rabbit antibodies against proteasome subunits ß1 (K43) and ß1i (K463) all in 1:1000 dilution for 1 h.
Membranes were washed 5 times for 20 min each time before being incubated with anti-rabbit immunoglobulin-POD conjugate for 30 min. Immunoreactive spots were visualized on photographic X-OMAT UV Plus film (Kodak, Stuttgart, Germany) by enhanced chemiluminescence using ECL reagent (Roth, Karlsruhe, Germany). An exposure time of 1 min was always applied.
Salivary gland biopsies for isolation of RNA
A total of 10 salivary gland biopsies were carried out for diagnostic purposes from consecutive patients presenting with sicca symptoms. From these, 4 patients were diagnosed as having primary SS. The remaining 6 patients showed no evidence of autoimmune sialadenitis and were used as controls. In order to avoid degradation of RNA, biopsies were stabilized in RNAlater (Qiagen). RNA was isolated using the NucleoSpin RNA XS kit (Macherey–Nagel, Düren, Germany) according to the manufacturer’s instructions, including on-column DNA digestion. RNA was reverse-transcribed into cDNA using Superscript III reverse transcriptase (Invitrogen, Karlsruhe, Germany).
Primer design for real-time polymerase chain reaction (PCR)
For analysis of gene expression of the 3 constitutive proteasomal catalytic subunits ß1, ß2, and ß5, and the corresponding inducible immunosubunits ß1i, ß2i, and ß5i in salivary gland biopsies, forward and reverse primers were selected based on the respective transcript sequences. Relative quantification of gene transcription was done in relation to the housekeeping gene ß-actin.
Primers for PCR were stringently designed using the Primer 3 software (Version 0.4.0; Whitehead Institute for Biomedical Research, Cambridge, MA, USA)23 with 19–21 bp length and an optimal annealing temperature of 62° ± 2°C to amplify cDNA fragments between 180 and 210 bp. Designed primer sequences are listed in Table 1.
Sequences of selected polymerase chain reaction (PCR) primers.
Primers were synthesized by TIB MolBiol (Berlin, Germany) and were tested in conventional PCR using the human monocytic cell line U937. Amplification products were controlled by sequencing (DLMB, Rüdersdorf, Germany). Primer efficiency was determined according to the protocols of Applied Biosystems (Foster City, CA, USA).
Relative quantification of gene expression
Relative quantification of gene expression was performed with real-time PCR using SYBR Green PCR Master Mix (Applied Biosystems) and 10 pmol from each forward/reverse primer. Amplifications were run in triplicates for each sample/primer pair using the ABI Prism 5700 sequence detection system (Applied Biosystems), including an initial step of 2 min at 50°C, an initial denaturation step of 10 min at 95°C, and 40 cycles with 95°C over 15 s for denaturation and 62°C over 60 s for annealing and elongation. Unspecific amplifications were controlled by running a dissociation protocol starting at 60°C after each experiment, which showed the specific melting point of each PCR product.
Measurement of gene expression
Relative quantification of gene expression at the mRNA level was done by comparison with expression of the housekeeping gene ß-actin. After checking the correct melting point of the PCR products, arithmetic means of CT values from triplicate samples were calculated. Differences in the expression of target genes were determined using the formula ECt housekeeping gene/ECt target gene, as described11, where E represents the efficiency of the respective primers. Standard deviations of CT triplicates < 0.5 were accepted for analysis.
Statistical analysis
Correlations between expression of proteasomal immunosubunits measured as the intensity of immunohistochemical staining (grade 0–3) and inflammatory infiltrates according to Chisholm-Mason grading were calculated using the Spearman r test.
To compare protein expression between different patient groups, non-parametric Mann-Whitney U-test and Fisher’s exact test were applied. Differences in the relative expression of investigated genes were analyzed using the nonparametric Mann-Whitney t-test. P values < 0.05 were considered significant.
RESULTS
Expression of proteasomal immunosubunits in minor salivary glands
The expression pattern of proteasomal immunosubunits was investigated in minor salivary glands using immunohistochemistry and immunofluorescence. To exclude an unspecific reaction of the secondary antibody, staining was also performed by omitting the primary antibody. This showed positive reactivity in the epithelium of the salivary ducts, but not in the glandular acini or in the inflammatory infiltrate. Therefore, the duct staining was considered as nonspecific background reactivity in the subsequent analysis (Figure 1A).

Staining pattern of proteasomal immunosubunits in minor salivary gland tissue. Results are representative for ß1i staining using K463. A. Epithelium cells of salivary ducts show nonspecific background reactivity using the secondary antibody (solid arrow). In contrast, acinus cells (broken arrow) and inflammatory infiltrates (arrowhead) were not stained (original magnification 200×). B. Representative anti-ß1i staining with a weak to moderate homogeneous pattern of epithelium acinus cells (broken arrow). Staining of the salivary ducts (solid arrows) is considered nonspecific (original magnification 50×). C. Detailed analyses revealed moderate ß1i staining in acinus cells (broken arrow) as well as in stromal lymphocytes and plasma cells (arrowheads). Staining of salivary ducts (solid arrow) considered nonspecific (400×).
Tissue-specific expression of inducible proteasomal subunits in minor salivary glands was identified in stromal lymphocytes and plasma cells as well as in the acinus epithelium by different primary antibodies against ß1i (K463 and K464), ß2i, and ß5i. Representative results for one patient with primary SS exhibiting moderate expression of ß1i using K463 are shown in Figure 1B and 1C.
Comparison off ß1i, ß2i, and ß5i staining in salivary glands of patients with SS and controls using immunohistochemistry
In all cases of positive staining, expression of the inducible subunits ß2i and ß5i was predominantly detectable in the inflammatory infiltrate, but also in acinus cells (Figure 2C, 2D). Importantly, staining of ß1i using K463 was significantly reduced in the majority of patients with SS (Figure 2B). Indeed, only 4 patients with SS showed detectable staining of ß1i using K463 in infiltrating immune cells and within serous and, to some extent, mucous acinus cells (Figure 2A). Using the polyclonal anti-ß1i antibody K464, no clear differences were observed between the analyzed groups (data not shown).

Staining of proteasomal immunosubunits in minor labial salivary glands of patients with primary SS, determined by immunohistochemical techniques. A. Strong staining of ß1i using K463 in infiltrating immune cells and within serous and to some extent mucous acinus cells of one patient with primary SS (Patient 3). B–D. Representative stainings for proteasomal immunosubunits in one patient with primary SS (Patient 4). B. Negative signal for ß1i using K463. C and D. Staining for ß2i and ß5i showed expression within acinus cells, infiltrating lymphocytes, and plasma cells (original magnification).
Comparing expressions of the inducible proteasomal subunits, there were no differences in the immunohistochemical stainings of ß2i or of ß5i between patients with SS (primary and secondary SS) and patients with sicca symptoms (Figure 3C and 3D, respectively). Only a slight but nonsignificant increase of ß5i expression was observed in patients with primary and secondary SS. In contrast, staining of ß1i using K463 was significantly reduced in the infiltrating lymphocytes as well as in the acinus cells of patients with SS (Mann-Whitney test, p = 0.0487, Fisher exact test, p = 0.0189; Figure 3A). This difference was even more pronounced when patients receiving hydroxychloroquine or prednisolone (SS, n = 4; sicca symptoms, n = 1) were excluded (Mann-Whitney test, p = 0.0122, Fisher exact test, p = 0.0059; Figure 3B).

Statistical analysis of proteasomal immunosubunits in minor salivary glands after immunohistochemical staining. A. ß1i staining (K463) was significantly decreased in patients with primary (pSS) and secondary SS (sSS) in comparison to patients with sicca symptoms (SiS) without autoimmune sialadenitis (Mann-Whitney test, p = 0.0487, Fisher exact test, p = 0.0189). B. This difference was even more pronounced in patients receiving hydroxychloroquine or prednisolone (Mann-Whitney test, p = 0.0122, Fisher exact test, p = 0.0059). C. ß2i staining showed no significant differences between patients with primary and secondary SS and patients with sicca symptoms without autoimmune sialadenits (Mann-Whitney test, p = 0.9811). D. Staining of ß5i was slightly but not significantly increased in patients with primary and secondary SS compared to patients with sicca symptoms without autoimmune background (Mann-Whitney test, p = 0.5520). ns: nonsignificant.
Expression of proteasomal immunosubunits and Chisholm-Mason grading
To investigate whether expression of proteasomal subunits was related to the magnitude of the inflammatory reaction in the salivary tissue, histological scoring was performed independently by 2 pathologists blinded for the diagnosis. Overall, staining for ß1i using K463 was found to be negatively correlated to Chisholm-Mason grade in patients with primary and secondary SS (Spearman r = −0.4611, p = 0.0135), but not in patients with sicca symptoms (Table 2). Further, no significant correlation was found between Chisholm-Mason grade and expression of the subunits ß2i and ß5i. Expression of proteasomal subunits was not related to other autoimmune manifestations or to autoantibodies against Ro and La antigens.
Autoantibody profiles, histological scores, and ß1i staining using K463 of different patient groups. Staining of ß1i using K463 and Chisholm-Mason grade were negatively correlated (Spearman r = −0.4611, p = 0.0135). No correlation was found between Chisholm-Mason grade and ß2i or ß5i staining (data not shown). Anti-Ro and anti-La antibodies were determined using automated fluorescence immunoassays (EliA, UniCap 100, Phadia, Freiburg, Germany.
Immunofluorescence comparison of lymphocytic infiltrates in salivary glands
Lymphocytic inflammatory infiltrates were identified in the minor labial tissue biopsies. Phenotypic analysis was based on immunofluorescence stainings with anti-CD20 and anti-CD27 antibodies. In our study, CD20-positive, CD27-negative cells represent naive B lymphocytes, which should appear green, while CD20 and CD27 double-positive cells represent memory B lymphocytes and should appear yellow on the overlays. Red-stained cells are CD27-positive, representing specifically T lymphocytes. Thus, no differences were observed between patients with primary or secondary SS and between patients with strong or weak ß1i expression of phenotype or distribution of infiltrating lymphocytes (Figure 4). Follicles could be identified with areas rich in CD27-naive B lymphocytes but with relatively few CD27-positive memory B cells. Areas with T lymphocytes could also be clearly distinguished.
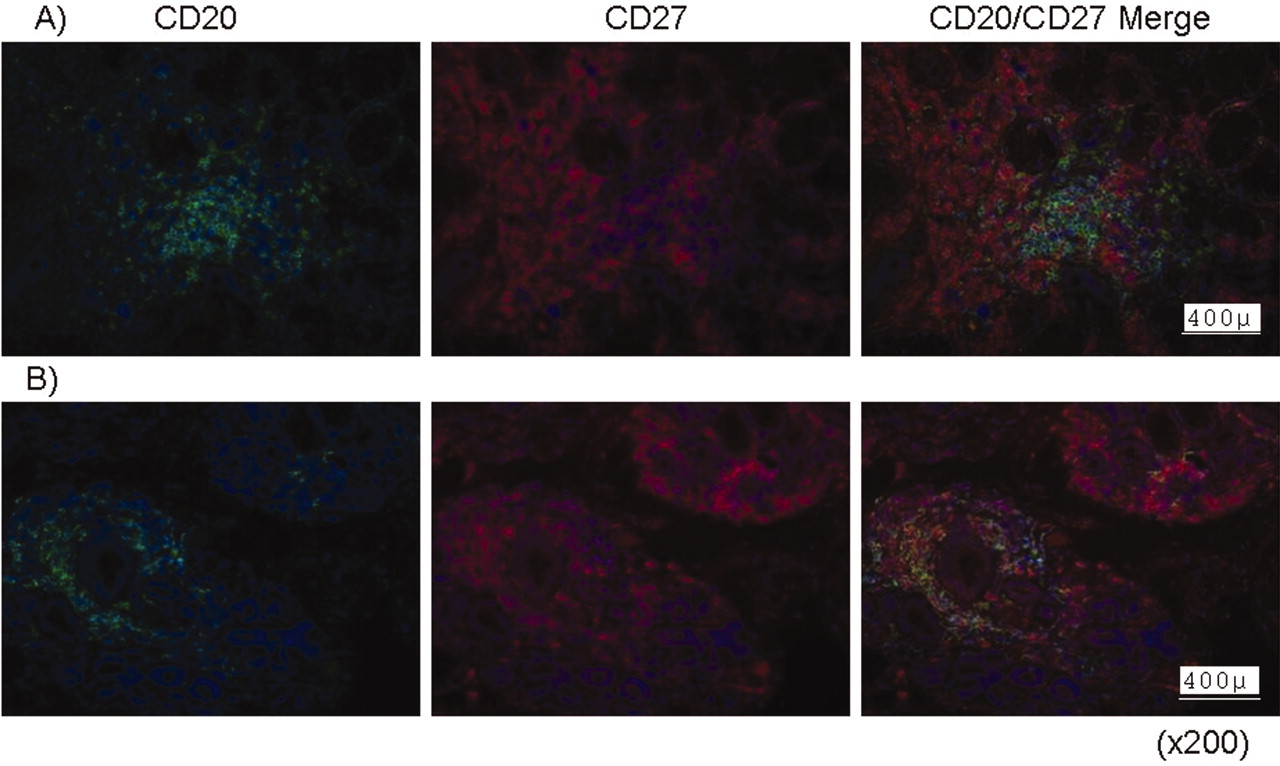
Representative immunofluorescence staining of lymphocyte subsets in infiltrates in labial biopsies from patients with primary SS (A) and secondary SS (B). CD20+/CD27– naive B cells (green), CD20+/CD27+ memory B cells (yellow), and CD27-positive T lymphocyte subsets (red) were detectable in both conditions with similar cellular distribution.
mRNA expression of ß1i, ß2i, and ß5i in patients’ salivary glands
At the mRNA level, expression of all the inducible proteasome subunits ß1i, ß2i, and ß5i was significantly higher in patients with SS compared to controls in PCR analyses (Figure 5). In contrast, no significant differences were found for levels of the respective constitutive subunits ß1, ß2, and ß5. Since inducible subunits replace their constitutive counterpart to form the 20S proteasome, we calculated the ratio of the respective subunits to compare the expression of inducible subunits. Using this approach, ß5i was identified as the subunit with the strongest expression (ratio of ß5 to ß5i = 0.88 in controls, ß5:ß5i = 0.16 in patients), followed by ß2i (ß2:ß2i = 6.15 in controls, ß2:ß2i = 1.88 in patients) and ß1i (ß1:ß1i = 20.03 in controls, ß1:ß1i = 1.70 in patients).

Comparisons of gene expression of inducible (ß1i, ß2i, and ß5i) and the corresponding constitutive (ß1, ß2, and ß5) proteasome subunits in salivary glands of SS patients (n = 4) and controls (n = 6). Nonparametric Mann-Whitney test with 95% confidence interval. For better appreciation of the magnitude of expression of each constitutive subunit compared with its inducible counterpart, the same scale was used for each subunit pair.
Detection of ß1 and ß1i subunits in patients’ PBMC using immunoblotting
To analyze expression of ß1i in the peripheral immune compartment, 3 different polyclonal anti-ß1i antibodies (PW8345, K463, and K464) were used for detection of ß1i subunit expression in PBMC from patients with SS (n = 3) compared to healthy controls (n = 2). No clear differences were observed using PW8345 and K464, while results with K463 showed slightly weaker signals for ß1i subunits in patients with SS (Figure 6A). To confirm this result, we compared expression of ß1 and ß1i in PBMC using identical amounts of protein in 2-D electrophoresis. Interestingly, the expression signal for ß1i was significantly stronger compared to the ß1 subunit in controls as well as in patients with SS. In subsequent analyses, we used one healthy control as a standard in comparison to 6 patients with SS. In densitometric evaluation comparing the stained dots for ß1i, a clearly weaker signal for ß1i was detected in 5 out of 6 patients with SS (Figure 6B).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Detection of the inducible ß1i subunit in PBMC of SS patients (P) and healthy controls (C). A. Immunoblotting revealed reduction of the ß1i signal using K463 polyclonal antibody, while no differences were observed using the polyclonal anti-ß1i antibodies PW8345 and K464. B. 2-D electrophoresis and immunoblotting using K463 revealed significantly stronger signal for ß1i compared to ß1 in controls as well as in patients with SS. Staining signal for ß1i using K463 was reduced in 5 out of 6 patients with SS compared to controls.
DISCUSSION
We analyzed expression of the 3 inducible proteasomal subunits ß1i, ß2i, and ß5i in minor salivary gland biopsy samples from patients with sicca symptoms. Patients with SS showed a characteristic dysregulation of the proteasomal immunosubunit ß1i in the targeted tissues, i.e., in the epithelium layer of acinus cells, compared to patients with sicca symptoms without autoimmune sialadenitis. The altered expression of ß1i in SS was visualized using polyclonal anti-ß1i antibodies, showing a significantly reduced staining signal in minor salivary glands. Interestingly, the reduced ß1i signal was negatively correlated to the intensity of lymphocytic infiltrates according to Chisholm-Mason grading. According to CD20 and CD27 expression24, the phenotype and distribution of B and T lymphocytes were similar in the salivary glands of patients with SS regardless of staining signals for ß1i. In contrast to the reduced staining signal for ß1i using K463, the proteasome subunit ß1i was significantly upregulated at the mRNA level in the tissues of patients with SS. In accord with the observation of altered ß1i expression in the inflammatory glandular infiltrates, the staining signal for ß1i was also reduced in PBMC of patients with SS compared to healthy controls using the same polyclonal anti-ß1i antibody. These findings suggest that the proinflammatory signal of the infiltrating cells does upregulate ß1i expression in the targeted tissue of patients with SS at the mRNA level. Despite the fact that mRNA and protein expression do not necessarily correlate, the reduced staining intensity using a polyclonal antibody indicates the altered antigenic properties of ß1i in immune cells of SS patients compared to controls.
Of note, in our cohort 3 out of 4 patients undergoing immunomodulating therapy and with disease quiescence expressed detectable levels of ß1i in the salivary glands as well as in the infiltrating lymphocytes. In contrast, all but one untreated patient with SS had infiltrating lymphocytes that showed altered expression of the ß1i protein. Although it is not possible to draw conclusions from such a limited number of patients, this could potentially indicate that expression of ß1i at the inflammatory site is influenced by treatment and perhaps represents a marker of pathogenic cells. Interestingly, induction of the constitutive alpha-type C3 subunit was observed after exposure to glucocorticoid therapy25. However, it remains to be confirmed whether expression of ß1i or other proteasomal subunits is also influenced by antiinflammatory drugs. By interference with signaling pathways of transcriptional factors like nuclear factor-κB such medications might be immune modulating of the ß1i-deficient cells. Investigations in larger cohorts are necessary to clarify whether this expression profile could be useful for diagnostic purposes or whether altered expression of ß1i characterizes a distinct subset of patients with SS.
A molecular defect in the bidirectional promoter region of the ß1i gene has been described in the non-obese diabetic (NOD) mouse, a model of type I diabetes and SS26. In consequence, reduced levels of ß1i were reported at the mRNA and protein levels, with implications for development of autoimmune disease in that particular animal model of autoimmunity. In SS, mRNA levels of ß1i were found to be upregulated in PBMC, but no genetic defect was found in the promoter region (Krause, et al19 and unpublished data). Therefore, observation of altered ß1i expression in the human disease remains obscure and could be explained by posttranscriptional alterations of ß1i turnover. Another possibity is a posttranslational modification of the ß1i protein, with a potential influence on antigenic properties, leading to altered antibody recognition. Thus, functional studies are required to confirm alterations in the proteasome system. Longterm observations of SS patients would be necessary to clarify the effects of ß1i dysregulation or modification on the disease course or outcome.
A polymorphism of ß1i that may be associated with different autoimmune diseases has been described27. This polymorphism in codon 60 (R/H) also resulted in reduced recognition of the protein by antibodies, but the explanation for this remains unclear.
An intact proteasome system is crucial for cellular homeostasis, activation of several transcriptional factors, and control of the immune response28⇓–30. In this regard, incorporation of immunoproteasome subunits into the catalytic complex is known to have a marked influence on the quality and quantity of the antigenic MHC I epitopes produced, including maintenance of a normal selection of self-peptides14,31⇓⇓–34. In this context, immunoproteasomes are also involved in effective clearance of, for example, viral infections13,35. However, altered expression of one singular proteasomal subunit could interfere with the assembly of the entire catalytic complex and alter the composition of certain immunoproteasome isoforms or hybrid proteasome complexes36. Given the influence of proteasomal subunit composition on the function of the proteasome system, our results suggest a possible role for the ß1i subunit in the pathogenesis of SS. Dysregulation of ß1i could potentially alter apoptosis of autoreactive T cells and lead to perpetuation of inflammatory responses. Further, proteasomes were also shown to be involved in the generation of intracellular peptides for inflammatory cell signaling37. Therefore, such altered proteasome expression could influence the initiation or maintenance of the inflammatory autoimmune response. Taken together, our data strongly encourage further investigations of the proteasome system in SS. Identifying tissue-or cell-specific markers for SS is crucial for our understanding of this complex disease, and could support early diagnosis and novel treatment strategies in clinical trials with targeted interventions of only disease-causing cells. The dysregulation of ß1i appears to be characteristic for SS, since it was rarely seen in patients with sicca symptoms who did not have autoimmune sialadenitis. The common dysregulation of ß1i in subsets of lymphoid cells of both SS-prone NOD mice and SS patients suggests that altered expression of ß1i protein could be a marker of pathogenetic cells38,39.
Acknowledgment
We thank Gabriele Fernahl, Institute of Pathology, Charité-Universitätsmedizin Berlin, for technical assistance.
Footnotes
Supported by the Deutsche Forschungsgemeinschaft (FE 470/3-1).
- Accepted for publication July 20, 2009.
REFERENCES
- 1.
- 2.
- 3.
- 4.
- 5.
- 6.
- 7.
- 8.
- 9.
- 10.
- 11.
- 12.
- 13.
- 14.
- 15.
- 16.
- 17.
- 18.
- 19.
- 20.
- 21.
- 22.
- 23.
- 24.
- 25.
- 26.
- 27.
- 28.
- 29.
- 30.
- 31.
- 32.
- 33.
- 34.
- 35.
- 36.
- 37.
- 38.
- 39.