

Abstract
Objective. The presence of anticitrullinated protein antibodies (ACPA) in rheumatoid arthritis (RA) indicates a breach in immune tolerance. Recent studies indicate that this breach extends to homocitrullination of lysines with the formation of anti-carbamylated protein (anti-CarP) antibodies. We analyzed the clinical and serologic relationships of anti-CarP in 2 RA cohorts.
Methods. Circulating levels of immunoglobulin G anti-CarP antibodies were determined by ELISA in established (Dartmouth-Hitchcock Medical Center) and early (Sherbrooke University Hospital Center) cohorts and evaluated for anticyclic citrullinated peptide antibodies (anti-CCP), specific ACPA, and rheumatoid factor (RF) levels using the Student t test and correlation analysis.
Results. We identified elevated anti-CarP antibodies titers in 47.0% of seropositive patients (Dartmouth, n = 164), with relationships to anti-CCP (p < 0.0001) and IgM-RF (p = 0.001). Similarly, 38.2% of seropositive patients from the Sherbrooke cohort (n = 171) had elevated anti-CarP antibodies; titers correlated to anti-CCP (p = 0.01) but not IgM-RF (p = 0.09). A strong correlation with anti-Sa was observed: 47.9% anti-Sa+ patients were anti-CarP antibodies+ versus only 25.4% anti-Sa− in the Sherbrooke cohort (p = 0.0002), and 62.6% anti-Sa+ patients versus 26.9% anti-Sa− were anti-CarP antibodies+ in Dartmouth (p < 0.0001). We found a more variable response for reactivity to citrullinated fibrinogen or to citrullinated peptides from fibrinogen and α enolase.
Conclusion. In 2 North American RA cohorts, we observed a high prevalence of anti-CarP antibody positivity. We also describe a surprising and unexpected association of anti-CarP with anti-Sa antibodies that could not be explained by cross-reactivity. Further, considerable heterogeneity exists between anti-CarP reactivity and other citrullinated peptide reactivity, raising the question of how the pathogenesis of antibody responses for carbamylated proteins and citrullinated proteins may be linked in vivo.
In addition to the formation of antibodies to citrullinated proteins [anticitrullinated protein antibodies (ACPA)], studies have suggested that the disease-specific breaches in immune tolerance in rheumatoid arthritis (RA) also extend to another posttranslational modification — homocitrullination of lysines. This modification is similar to the citrullination of arginines with the same functional ureido group. Enzymatic catalysis of arginine to citrulline is mediated by peptidylarginine deiminase (PADI)1. In contrast, homocitrullination involves chemical carbamylation of the primary amine group of lysine2 through a reaction with cyanate. Presumably, this process occurs at inflammatory sites by the action of myeloperoxidase3,4, although carbamylation can also occur as a result of the spontaneous reversible dissociation of urea5,6. Since both PADI 4 and myeloperoxidase are found in the azurophilic granules of neutrophils, it seems likely that these posttranslational modifications occur at inflammatory sites. In this regard, fibrinogen has been shown to be a target for both modifications7.
Humoral responses to homocitrullinated proteins [subsequently referred to as anti-carbamylated protein (anti-CarP) antibodies] have been reported in both patients with early and established seropositive RA, as well as by a proportion of seronegative patients with RA7,8,9. Indeed, anti-CarP antibodies such as ACPA can be found in patient sera years before the onset of RA, with a median time of about 5 years from first serologic appearance to the onset of clinical signs and symptoms8,10.
Given the similarity of these posttranslational modifications of basic amino acids, it is not surprising that some, but not all, ACPA and anti-CarP antibodies in patient sera have been reported to exhibit cross-reactivity11,12. It is additionally clear that some ACPA and anti-CarP antibodies demonstrate remarkable fine specificity, being capable of discriminating between citrullinated and homocitrullinated forms of the same protein7,10. This is perhaps best demonstrated by the presence of anti-CarP antibodies in ACPA-negative patients10.
Anti-Sa antibodies represent 1 subfamily of ACPA that specifically targets citrullinated vimentin, with prior research suggesting that they arise following the formation of neutrophil extracellular traps and the subsequent breach of self-tolerance that leads to the development of RA13. Present in a subset of about 40% of patients with RA14, anti-Sa antibodies are notable for their high specificity (> 95%) for RA15,16 and strong correlation with poor disease outcomes including radiographic progression compared with anti-Sa−negative patients17.
In our study, we analyzed the relationships between serum/plasma levels of anti-CarP antibodies and several clinical and serologic variables, including anti-Sa status. We used both an established and an early RA cohort to confirm our findings.
MATERIALS AND METHODS
Study population
Our study was derived from 2 North American cohorts of patients with RA based at the Dartmouth-Hitchcock Medical Center in Lebanon, New Hampshire, USA, and the Sherbrooke University Hospital Center in Sherbrooke, Quebec, Canada18. Shared epitope status and radiograph progression were available as previously described18. A total of 548 patients with RA and 65 healthy controls were involved in the analyses. All patients with RA fulfilled the revised 1987 American College of Rheumatology criteria for the classification of RA19. All participants provided written informed consent, and ethical permission was obtained from the institutional review boards of both institutions.
The Dartmouth cohort was composed of patients with established arthritis followed in the Rheumatology Clinic at the Dartmouth-Hitchcock Medical Center. This cohort included 212 patients with RA (Table 1). Demographic and serologic data are available on all patients. Serologic status was determined prior to enrollment by nephelometry [immunoglobulin M (IgM) rheumatoid factor (RF) > 14 IU/ml using immunoturbidimetric measurement; Roche Diagnostics] and/or IgG anticyclic citrullinated peptide (anti-CCP) > 5.0 U/ml by ELISA (DiaSorin), with seronegativity being defined as negative for both RF and anti-CCP at all measurements. Subsequently, IgM-RF and anti-CCP were measured by ELISA using the serum sample obtained at enrollment within the cohort as outlined below, and reactivity defined as per the manufacturer. This resulted in 3 samples being reclassified as seropositive because of positive RF reactivity.
Features of patients from both RA cohorts. In the Dartmouth seronegative patients, only 28 of 45 had measured values for total IgG, CXCL13, and CXCL10. When percentages are given, but not all patients in that group were tested (i.e., shared epitope), the “n” is displayed as a fraction representing number of positive over number of those tested. Classification by sero status and anti-Sa reactivity was determined using different methods for the 2 cohorts. Values are mean ± SD unless otherwise specified.
The Sherbrooke cohort consisted of a subset of the patients recruited at the Sherbrooke University Hospital Center as part of the longitudinal Early Undifferentiated Polyarthritis (EUPA) cohort. In contrast with the Dartmouth established RA cohort, this cohort represented an early arthritis population, consisting of patients who were largely naive to disease-modifying antirheumatic drugs (DMARD) and corticosteroids at the time of inclusion, and with nearly equivalent numbers of seropositive (n = 171) and seronegative (n = 165) patients (Table 1). Cohort inclusion criteria included disease duration between 1 and 12 months and swollen joint count of 3 or more (based on the 66-joint count). For the study, seropositivity was defined as an IgM-RF titer ≥ 40 IU/ml measured using RapiTex RF (Dade Behring) and IgG anti-CCP > 20 U/ml using QUANTA Lite (Inova Diagnostics) concurrently at least once. Seronegativity was defined as negative RF and anti-CCP at all visits. This subset of patients was chosen randomly from the Sherbrooke EUPA cohort, with samples matched only for serologic status. Information gathered at enrollment included demographic details such as age, sex, and time since onset of arthritis, as well as clinical details such as the Disease Activity Score at 28 joints (DAS28) with C-reactive protein (CRP) score, radiographic Sharp/van der Heijde score of hand and feet films, and RF and ACPA status.
Serum/plasma analysis
Patient serum and plasma were stored at −80°C (Dartmouth cohort) or −20°C (Sherbrooke cohort) until first analysis and then maintained at 4°C while additional assays were performed. Antibodies directed against CarP were detected using carbamylated fetal calf serum (FCS) as previously described10. FCS was diluted 1:250 in water and then incubated with 1M KCN (Sigma-Aldrich) for 12 h at 37°C prior to 72 h dialysis against water at 4°C. For competition studies (Figure 1), this protocol was modified to incubating FCS at a 1:1 ratio with KCN (final 1M concentration) for 12 h at 24°C before dialysis against phosphate buffered saline (PBS). Successful carbamylation of FCS was confirmed by immunoblot using a rabbit anti-carbamyl-lysine polyclonal antibody (Cell Biolabs Inc.).

Competition for anti-Sa reactivity. (A) Immunoblot analysis of native FCS and carbamylated FCS using rabbit anti-carbamyl lysine antibody confirming extensive carbamylation. (B) Five sera were tested for anti-Sa reactivity or (C) CarP reactivity in the absence or presence of varying (1:100 and 1:10) dilutions of carbamylated FCS (0.32–3.2 mg/ml). FCS: fetal calf serum; CarP: carbamylated protein.
Ninety-six well plates (R&D Systems) were coated with carbamylated FCS or non-carbamylated FCS at 4°C overnight and then blocked with 1% bovine serum albumin (Sigma) in PBS. Human serum or plasma was incubated at 4°C overnight. Bound human IgG antibodies were detected after incubation for 3.5 h at 4°C with horseradish peroxidase-conjugated goat anti-human IgG (H+L) F(ab′)2 antibody fragment (KPL, Kirkegaard & Perry Laboratories Inc.) at a 1:10,000 dilution, followed by the addition of tetra-methylbenzidine substrate (R&D Systems) and subsequent administration of stop solution. Absorbance was measured at 450 nm (reference wavelength of 570 nm). Optical density was transformed into arbitrary units (AU/ml) using the titration curve of a strongly anti-CarP antibody+ serum sample, whose anti-CarP antibodies concentration was set at 500 AU/ml. To analyze specific anti-CarP antibody reactivity, the background signal with non-carbamylated FCS was subtracted from the carbamylated FCS signal. The cutoff for a positive response was calculated as the mean (4.2 AU/ml) plus 2 times the SD (9.0 AU/ml) of the specific anti-CarP reactivity of the healthy controls, corresponding to a level of 22.16 AU/ml.
For both the Dartmouth and the Sherbrooke cohorts, serum levels of anti-Sa IgG were measured by ELISA (EUROIMMUN). For both cohorts, ELISA was used to determine IgG levels (Immunology Consultants Laboratory), IgM-RF levels (TheraTest Laboratories), and C-X-C motif chemokine 13 (CXCL13) and CXCL10 levels (R&D Systems). In the Dartmouth cohort, we additionally measured serum levels of IgG ACPA using a human QUANTA Lite CCP3 IgG ELISA kit (Inova Diagnostics).
Competition studies
The cross-reactivity of anti-CarP antibodies for the Sa antigen (citrullinated vimentin) was examined in sera from the Dartmouth cohort. Sera (n = 5) were left untreated or were preincubated [15’ at room temperature (RT)] with carbamylated FCS at a concentration of either 1:10 or 1:100. Samples were then transferred to a 96-well plate coated with Sa antigen or carbamylated FCS, and levels of IgG reactivity were measured by ELISA, as described above.
Multiplex assay
A subset of the Dartmouth cohort was analyzed for reactivity to citrullinated native fibrinogen and to synthetic citrullinated peptides using a bead-based multiplex array platform. Specified peptides were synthesized with C- and N-terminal cysteines to cyclize the peptide that included an N-terminal biotin. Microsphere beads (LumAvidin, Luminex Inc.) were coated with a > 10-fold molar excess of each type of biotinylated antigen (citrullinated, native, or other ligand controls), and then blocked with free biotin. A master mixture of different coated microspheres was made. Serum samples were diluted 1:1000 in SM01 buffer (Surmodics Inc.), and then 200 μl of diluted serum was incubated with 10 μl of the antigen-coated microsphere mixture, representing 2000 beads/analyte. Samples were incubated overnight at 4°C with shaking at 50 rpm, then loaded into individual duplicate wells of a 96-well vacuum filter plate (Millipore). Wells were then washed thrice with PBS-Tween 20 (v/v 0.05%). For detection, 100 μl of PE-conjugated anti-human IgG Fcγ-specific antibody (eBioscience) diluted 1:250 in SM01 buffer was added to each well, and incubated for 1 h at RT, with shaking. After 3 washes, microspheres were resuspended at 150 μl and data collected using a Luminex 200 (Luminex Inc.). Data represent mean median fluorescence intensity values from duplicate wells.
Statistical analysis
Statistical analyses were performed using GraphPad Prism software version 6 (GraphPad Software). Levels of IgG, anti-CCP, CXCL10, CXCL13, high-sensitivity CRP, and anti-Sa were log-transformed to normalize the data. Student t test for independent samples was used to compare the means of 2 sets of data. Pearson correlation was used for analysis of log-transformed anti-CarP antibody and other measures. Chi-square analysis was used for determination of anti-CarP antibody+ relationship to the shared epitope. All p values were 2-tailed, with values < 0.05 considered significant.
RESULTS
Elevated anti-CarP antibody titers in seropositive patients with RA
The Dartmouth RA cohort (n = 212) is an established RA cohort, including patients with a broad range of disease duration (Table 1). We analyzed serum/plasma anti-CarP antibody levels in seropositive and seronegative patients with RA relative to a control group of healthy individuals (n = 65) to establish baseline values. Among the healthy cohort, the mean anti-CarP antibody level was measured at 4.2 AU/ml, with over 50% of donors having no detectable signal. We established the cutoff for anti-CarP antibody positivity at 22.16 AU/ml, corresponding to the mean plus 2 times the SD (9.0 AU/ml). Three healthy donors (4.6%) had values above this level; this is similar to levels (6.3%) previously described8.
Among seropositive patients with RA (n = 167), 46.1% exhibited elevated levels of anti-CarP antibodies (Figure 2A), with a median of 18.9 AU/ml, mean of 70.2 AU/ml, and an interquartile range (IQR) of 6.8–58.7 AU/ml. The distribution of anti-CarP antibody concentrations was strongly positively skewed, with observed levels ranging as high as 1550 AU/ml. Because of the non-normal pattern of the distribution, anti-CarP antibody levels were log-transformed for analysis. Among seronegative patients (n = 45), 8.9% were anti-CarP antibody+, and anti-CarP antibody levels fell within a more narrow range, with an IQR of 5.9–11.0 AU/ml. An anti-CarP antibody titer greater than 40 AU/ml was observed in only 1 patient (2.2%), while this value was exceeded in nearly one-third (31.1%) of seropositive patients. This patient had been seronegative for ACPA and RF on multiple determinations over > 10 years. Overall, the mean anti-CarP antibody level in seropositive patients with RA was significantly higher than in otherwise seronegative patients (70.2 AU/ml vs 7.6 AU/ml, p < 0.0001; Figure 2A).
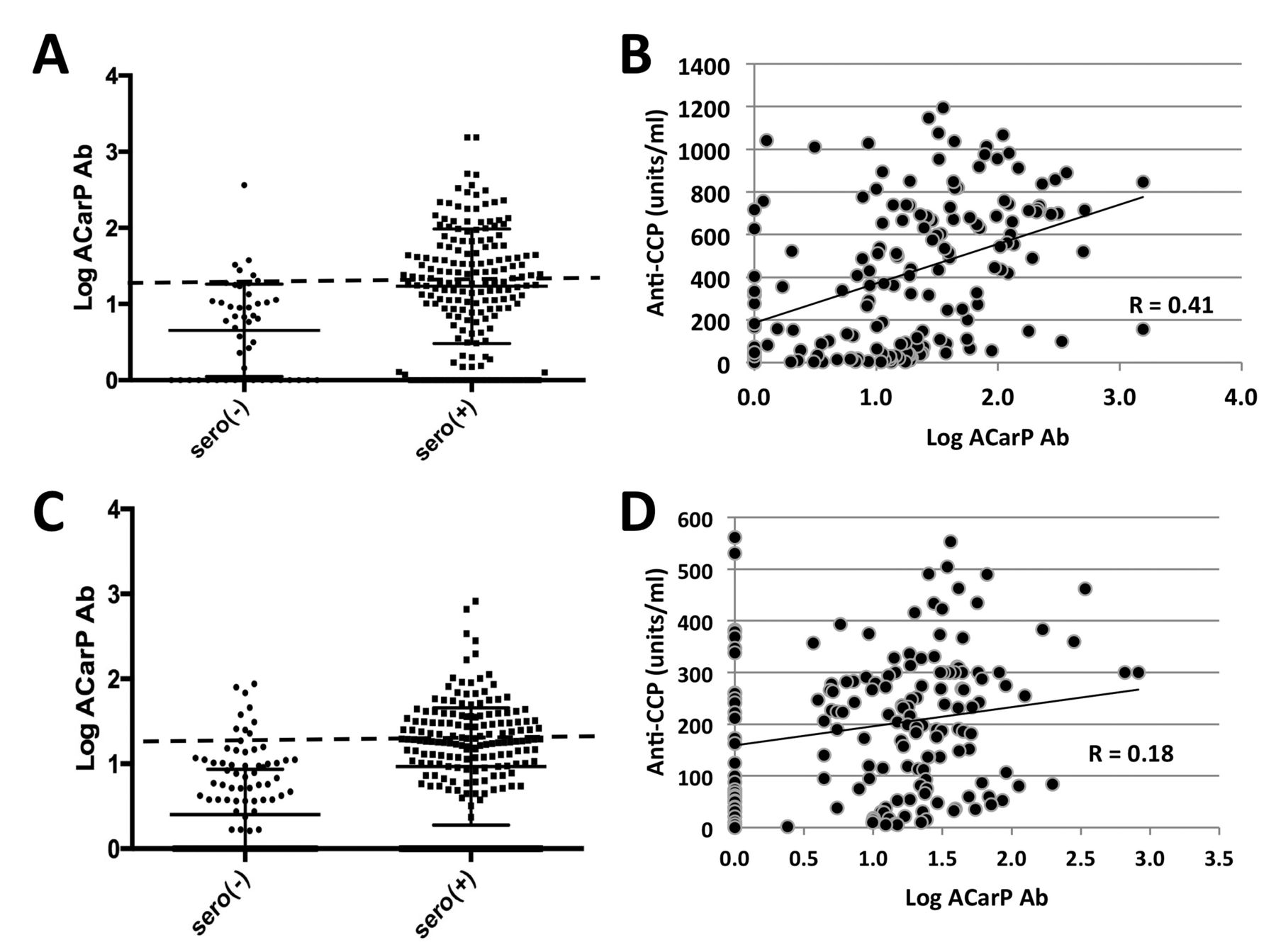
Relationships of anti-CarP antibody titer to seropositivity and anti-CCP. (A) and (B) represent the Dartmouth established RA cohort, and (C) and (D) represent the Sherbrooke EUPA cohort. (A and C) Log-transformed anti-CarP antibody is higher in seropositive than in seronegative patients with RA in both the Dartmouth cohort [seronegative: n = 45, mean (SEM) = 0.66 (0.08); seropositive: n = 167, mean (SEM) = 1.23 (0.05), p < 0.0001] and the Sherbrooke cohort patients [seronegative: n = 165, mean (SEM) = 0.40 (0.05); seropositive: n = 171, mean (SEM) = 0.97 (0.05), p < 0.0001]. (B and D) Among seropositive samples, a positive correlation is observed between anti-CarP antibody titer and anti-CCP in both cohorts (p < 0.0001, p = 0.01, respectively). Diagonal lines represent lines of best fit. Anti-CarP: anti-carbamylated protein; anti-CCP: anticyclic citrullinated peptide antibody; RA: rheumatoid arthritis; SEM: standard error of the mean; EUPA: Early Undifferentiated Polyarthritis.
Anti-CarP antibody relationships with other serologic and clinical features
Among seropositive patients with RA in this cohort, we observed a significant positive correlation between levels of anti-CarP antibodies and anti-CCP (R = 0.41, p < 0.0001; Figure 2B). Among this group, anti-CarP antibody+ patients had a median ACPA of 660 units/ml (IQR 373–788) versus a level of 143 units/ml (IQR 27–450) among anti-CarP antibody– patients (p < 0.0001). Anti-CarP antibody titers among seropositive patients also demonstrated a direct but weaker relationship with IgM-RF (R = 0.25, p = 0.001; Supplementary Figure 1A, available online at jrheum.org). Additional analyses of this cohort demonstrated a lack of relationship between anti-CarP antibody titer and total serum IgG levels in the seropositive patients (R = 0.15, p = 0.06; Supplementary Figure 1B, available online at jrheum.org).
We have previously identified a strong relationship to the B cell chemokine CXCL13 and autoantibody status18. We evaluated the relationship of anti-CarP titer with CXCL13 as well as CXCL10, another chemokine strongly associated with RA. We did identify correlations in seropositive patients with RA between anti-CarP antibody titer and the levels of the chemokines CXCL13 (R = 0.21, p = 0.007; Supplementary Figure 1C, available online at jrheum.org) and CXCL10 (R = 0.18, p = 0.03; Supplementary Figure 1D, available online at jrheum.org), but no relationships were identified with age (R = −0.01, p = 0.87; Supplementary Figure 1E, available online at jrheum.org) or sex (p = 0.08; Supplementary Figure 1F, available online at jrheum.org). Similarly, anti-CarP positivity was not more commonly associated with inheritance of those MHC II alleles associated with the shared epitope that are the strongest set of genetic susceptibility factors for RA (p = 0.61, n = 107; data not shown).
Relationships between anti-CarP antibody and antibody levels in an early RA cohort
We confirmed these results in validation studies of the Sherbrooke EUPA cohort, a cohort of predominantly DMARD- and corticosteroid-naive patients with early RA (Table 1). Seropositive patients in this cohort (n = 171) displayed trends similar to their Dartmouth counterparts with established RA; 38.2% had elevated anti-CarP antibody levels (p < 0.0001; Figure 2C). Once again, the anti-CarP antibody concentrations were not normally distributed, with a median of 17.0 AU/ml, mean of 34.9 AU/ml, IQR of 5.0–31.3 AU/ml, and a maximal value of 821 AU/ml. As with the Dartmouth cohort, the anti-CarP antibody levels had a positive correlation with anti-CCP levels (R = 0.2, p = 0.01; Figure 2D); however, no relationship was seen with IgM-RF (R = 0.13, p = 0.09) or serum total IgG (R = 0.097, p = 0.2; Supplementary Figures 2A and 2B, available online at jrheum.org). When results from seropositive patients from the Dartmouth and Sherbrooke cohorts were pooled, anti-CarP antibody levels maintained a direct relationship with ACPA levels (R = 0.35, p < 0.0001; Supplementary Figure 4A, available online at jrheum.org) with a weaker relationship to IgM-RF (R = 0.16, p = 0.004; Supplementary Figure 4B, available online at jrheum.org).
Among seronegative patients (n = 165), 6.1% were anti-CarP antibody+, and the distribution of anti-CarP antibody levels were within a relatively narrow range, with an IQR of 0.0–7.3 AU/ml. An anti-CarP antibody titer greater than 40 AU/ml was observed in 4 patients (2.4%) compared with 31 of 173 (18%) seropositive patients. Further, akin to the Dartmouth cohort, the mean anti-CarP antibody level in seropositive patients with RA was significantly higher than in seronegative patients (34.7 vs 5.8 AU/ml, p < 0.0001).
As with the Dartmouth cohort, no relationships between anti-CarP antibody levels and age or sex were observed among seropositive Sherbrooke patients (Supplementary Figures 2E and 2F, available online at jrheum.org). In this cohort, we also have clinical variables such as DAS28-CRP, CRP levels, and erosions, yet none exhibited any relationships to anti-CarP antibody levels (Supplementary Figures 3A–C, available online at jrheum.org). We were able to evaluate anti-CarP reactivity in relation to being a nonsmoker or to being a current or former smoker, and found that there was a relationship, with a mean anti-CarP antibody level of 24.1 and 40.3 AU/ml, respectively (p = 0.02; Supplementary Figure 3D available online at jrheum.org). There were no correlations with CXCL13 (R = 0.08, p = 0.31) or CXCL10 (R = 0.09, p = 0.26; Supplementary Figures 2C and 2D, available online at jrheum.org), and we saw no relationship with the shared epitope (p = 0.3, n = 141; data not shown).
Relationship between anti-CarP antibody and anti-Sa antibody levels
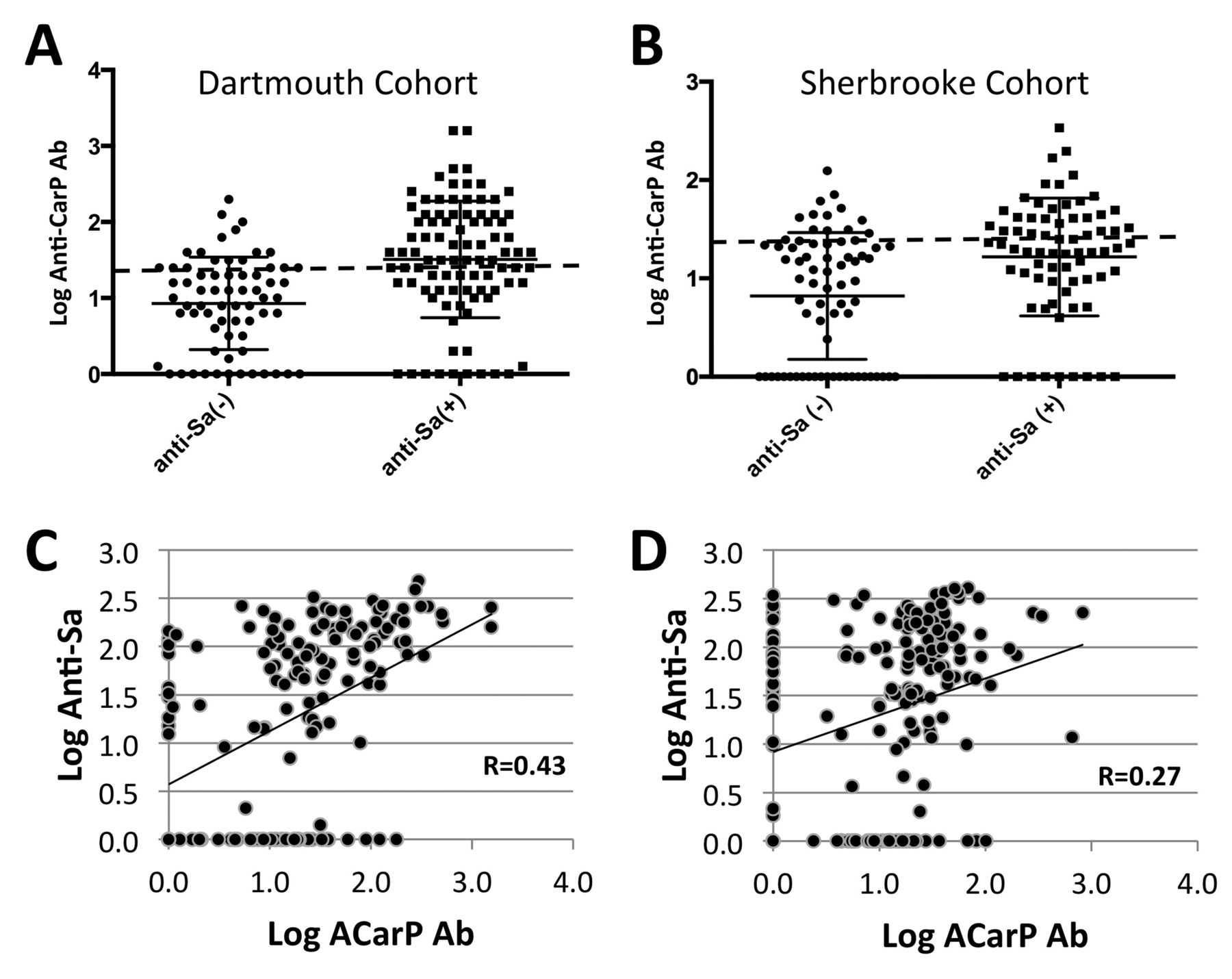
The Sherbrooke cohort has previously been evaluated for anti-Sa status, which has been shown to be a biomarker for RA disease and severity20,21. In the current studies, we found that anti-CarP positivity was more highly represented in the anti-Sa+ patients (47.9%) compared with only 25.4% percent of anti-Sa− patients (p = 0.0002; Figure 3B). Moreover, anti-Sa+ patients in this cohort had higher anti-CarP antibody levels than anti-Sa− patients (37.6 AU/ml and 14.6 AU/ml, respectively). Intrigued by this association, we tested seropositive samples from the Dartmouth cohort for anti-Sa antibody level (n = 158). Once again, anti-Sa status had a strong relationship with anti-CarP antibody status (p < 0.0001), with 62.6% of anti-Sa+ patients also being anti-CarP+ versus only 26.9% of anti-Sa− patients (Figure 3A). Similarly, the mean anti-CarP antibody titer was higher among anti-Sa+ patients (112.5 AU/ml) relative to the anti-Sa−, wherein the mean titer (19.1 AU/ml) was below the threshold for positivity. Correlation analysis of anti-CarP antibody titers and anti-Sa titers also yielded a significant relationship in both the Dartmouth cohort (R = 0.43, p < 0.0001; Figure 3C) and the Sherbrooke cohort (R = 0.27, p = 0.0002; Figure 3D). There were also 133/165 seronegative patients from the Sherbrooke cohort and 4/45 from the Dartmouth cohort tested for their anti-Sa status. Among these samples, 14 donors were anti-CarP antibody+, 4 were anti-Sa antibody+, and only 1 was positive for both anti-CarP and anti-Sa antibodies.

Anti-CarP antibody titer robustly correlates with anti-Sa antibody levels among seropositive patients with RA in both the Dartmouth and Sherbrooke cohorts. (A) Anti-Sa+ among seropositive patients in the Dartmouth cohort also exhibit a significantly higher mean anti-CarP antibody level than anti-Sa– among seropositive patients [anti-Sa positive: n = 91, mean (SEM) = 1.51 (0.08); anti-Sa negative: n = 67, mean (SEM) = 0.93 (0.07), p < 0.0001]. (B) Anti-Sa+ among seropositive patients in the Sherbrooke cohort have a significantly higher mean anti-CarP antibody level than anti-Sa– among seropositive patients [anti-Sa positive: n = 72, mean (95% CI) = 1.22 (1.15–1.29); anti-Sa negative: n = 71, mean (95% CI) = 0.82 (0.74–0.90), p = 0.0002]. (C) Within the Dartmouth cohort, anti-CarP antibody is significantly correlated with anti-Sa antibody titer (n = 158, p < 0.0001). (D) A similar relationship is seen with seropositive patients with RA from the Sherbrooke cohort (n = 169, p = 0.0002). Anti-CarP: anti-carbamylated protein; RA: rheumatoid arthritis; SEM: standard error of the mean.
The cross-reactivity of anti-CarP antibodies with anti-Sa reactivity was evaluated using carbamylated FCS (Figure 1A). Preincubation with carbamylated FCS blocked anti-CarP reactivity, but had no effect on anti-Sa levels (Figures 1B and 1C). These data are consistent with the interpretation that the association of anti-Sa and anti-CarP reactivity arises from coexpression rather than cross-reactivity.
To further evaluate for interactions of anti-CarP antibodies with anticitrullinated proteins, we analyzed 31 seropositive serum samples for antibody reactivity to CarP (13 anti-CarP+, 18 anti-CarP−), Sa antigen, and human citrullinated (cit-) full-length fibrinogen. Anti-CarP positivity was significantly associated with higher levels of reactivity with Sa (as described above) as well as with cit-fibrinogen relative to anti-CarP–negative sera (p < 0.0001). In contrast, the presence of anti-cit-fibrinogen antibodies does not seem linked to the presence of anti-Sa (Figures 4A and 4B). When specific citrullinated peptide reactivity was tested, a great deal of variability was seen in anti-CarP+ sera, although there was less heterogeneity seen with reactivity to cit-vimentin 59–78 and cit-fibrinogen-β 36–52 (Figure 4C).

Variability in reactivity to CarP, Sa antigen, cit-fibrinogen, and cit-peptides in the Dartmouth Cohort. (A) Anti-CarP+ samples have a stronger relationship to cit-fibrinogen reactivity than in (B) anti-CarP– samples (p < 0.0001). There is no relationship between anti-Sa and cit-fibrinogen values in either the anti-CarP+ samples (R = −0.08, p = 0.79, n = 13) or in the anti-CarP– samples (R = −0.028, p = 0.91, n = 18). (C) Using a multiplex fluorescent bead array, antibody reactivity to cit-vimentin 59–78, cit-fibrinogen-α 616–635, cit-fibrinogen-α 39–50, cit-fibrinogen-β 36–52, and cit-α-enolase 5–21 was measured in anti-CarP antibody+ (n = 14) and anti-CarP antibody– (n = 30) samples. Values displayed represent AU/ml for CarP reactivity while the values for the peptides represent mean MFI of reactivity to the citrullinated peptide minus the value obtained from native peptide (corrected value). CarP: carbamylated protein; cit-: citrullinated; MFI: median fluorescence intensity.
DISCUSSION
In the current studies, we observed that levels of anti-CarP antibodies are elevated in about half (46.1%) of established seropositive patients with RA, while we found a somewhat lower frequency (38.2%) of anti-CarP antibodies in seropositive patients from an early RA cohort. These data are compatible with prior reports of frequencies of 49–73% anti-CarP antibody positivity among seropositive patients with RA in early RA cohorts9,10 and 40–55% in established RA cohorts7. We also found anti-CarP antibodies in only a small percentage (6.1–8.9%) of seronegative patients with RA, and our results suggest that most seronegative patients with RA with anti-CarP positivity are low-titer positive, rarely having levels more than 1–2 SD above the determined cutoff for seropositivity. The comparable frequency and titers of the anti-CarP antibodies in these 2 cohorts may suggest that anti-CarP antibody levels remain relatively stable over time. However, additional research is needed to evaluate the stability of anti-CarP titer over the course of RA treatment. Additionally, although these clinical centers used 2 different methods for determining seropositivity, similar results were documented across both cohorts, which further strengthens the general relevance of these findings.
We also confirmed an association between anti-CCP and anti-CarP antibody titers in both cohorts, as previously reported by Shi, et al10. Various associations (i.e., with IgM-RF, CXCL13, and CXCL10) were observed in the Dartmouth cohort, but not found in the Sherbrooke cohort. The association between anti-CarP antibody titer and anti-CCP, but absence of an association with the shared epitope, supports findings by Jiang, et al, and provides further evidence that this immunoreactivity may arise through different pathogenic pathways9.
An unexpected finding was the high concordance between anti-Sa antibody status and anti-CarP autoantibody positivity and levels. This association was at least as strong as the association of anti-CarP and anti-CCP reactivity in the Dartmouth cohort. Anti-Sa represents a type of antibody reactivity with citrullinated vimentin, an intermediate filament cytoplasmic protein. In contrast, anti-CarP autoantibodies are detected using carbamylated FCS as the source of antigenic ligand, and frequently react with carbamylated fibrinogen7,10.
Several lines of evidence suggest anti-CarP and anti-Sa constitute distinct antibody specificities rather than reflect broadly cross-reactive antibodies. Figure 4A shows that specific anti-CarP positivity using carbamylated FCS was better correlated with reactivity with Sa relative to citrullinated forms of fibrinogen, which is a major target of carbamylation in FCS. Second, anti-Sa reactivity was found in anti-CarP–negative sera (Figure 4B). Finally, competition experiments in patients who contained both anti-CarP and anti-Sa reactivity did not suggest cross-reactivity (Figure 1). Clarifying this relationship would be enhanced by mapping the neoepitopes in fibrinogen generated by carbamylation that are being recognized in patients with RA.
This finding is potentially interesting because of the reports that anti-CarP antibody positivity is associated with radiographic progression10. The same relationship has been reported with anti-Sa17,22,23. Our data raise the possibility that the coexpression of anti-CarP and anti-Sa may be confounding these reports. Alternatively, the presence of both anti-Sa and anti-CarP might be associated with an improved ability to predict severe erosive RA. Additional evaluation is needed to find out whether anti-CarP has added value to anti-Sa in the determination of disease activity and prognosis. The absence of cross-reactivity between these assays, in contrast to that seen with ACPA and anti-CarP, may make these tests important adjuncts to ACPA testing in predicting disease outcome.
ONLINE SUPPLEMENT
Supplementary data for this article are available online at jrheum.org.
Footnotes
Supported by the National Institutes of Health grant R21 AR-061643 from the National Institute of Arthritis and Musculoskeletal and Skin Diseases (to WFCR) and the Rheumatology Research Foundation (to WFCR and a separate award to GJS), by the Hitchcock Foundation (to JDJ), and by grants 00/201 and RG06/108 from The Arthritis Society and Canadian Institutes for Health Research grant MOP-110959 (to GB). AJB-F, AM, and GB are members of the Centre de Recherche Clinique Etienne-LeBel at the Centre Hospitalier Universitaire de Sherbrooke, which received a team grant from the Health Research Funds of Quebec. Since 2007, the Sherbrooke Early Undifferentiated Polyarthritis cohort has received financial support from the Canadian ArTHritis CoHort (CATCH) study, designed and implemented by investigators and financially supported initially by Amgen Canada and Pfizer Canada by an unrestricted research grant. As of 2011, further support was provided by Hoffmann-La Roche, Chemicals of Belgium (UCB) Canada, Bristol-Myers Squibb Canada, Abbott Laboratories, and Janssen Biotech (a wholly owned subsidiary of Johnson & Johnson).
- Accepted for publication October 7, 2015.








{kind=link}
{kind=link}
{kind=link}
{kind=link}