

Abstract
Objective. Cartilage damage is frequently observed in advanced destructive gout. The aim of our study was to investigate the effects of monosodium urate monohydrate (MSU) crystals on chondrocyte viability and function.
Methods. The alamarBlue assay and flow cytometry were used to assess the viability of primary human chondrocytes and cartilage explants following culture with MSU crystals. The number of dead chondrocytes in cartilage explants cultured with MSU crystals was quantified. Real-time PCR was used to determine changes in the relative mRNA expression levels of chondrocytic genes. The histological appearance of cartilage in joints affected by gout was also examined.
Results. MSU crystals rapidly reduced primary human chondrocyte and cartilage explant viability in a dose-dependent manner (p < 0.01 for both). Cartilage explants cultured with MSU crystals had a greater percentage of dead chondrocytes at the articular surface compared to untreated cartilage (p = 0.004). Relative mRNA expression of type II collagen and the cartilage matrix proteins aggrecan and versican was decreased in chondrocytes following culture with MSU crystals (p < 0.05 for all). However, expression of the degradative enzymes ADAMTS4 and ADAMTS5 was increased (p < 0.05 for both). In joints affected by gout, normal cartilage architecture was lost, with empty chondrocyte lacunae observed.
Conclusion. MSU crystals have profound inhibitory effects on chondrocyte viability and function. Interactions between MSU crystals and chondrocytes may contribute to cartilage damage in gout through reduction of chondrocyte viability and promotion of a catabolic state.
Cartilage damage is often observed in joints affected by advanced gout1. Ultrasonographic studies have highlighted the close relationship between monosodium urate monohydrate (MSU) crystals and articular cartilage in patients with gout. The “double contour” sign, a hyperechoic band over the superficial margin of articular cartilage, is considered diagnostic of gout, and is thought to represent MSU crystals coating articular cartilage2,3,4. The close relationship between MSU crystals and cartilage is further supported by histopathological observations of MSU crystals deposited radially in the superficial layers of articular cartilage in gouty joints5,6.
Following interaction with MSU crystals, chondrocytes may contribute to cartilage damage in gout through inflammatory mechanisms such as increased production of degradative enzymes and proinflammatory mediators. Liu-Bryan, et al7,8 reported that MSU crystals induce nitric oxide generation and expression of matrix metalloprotease-3 (MMP-3) in articular chondrocytes through Toll-like receptor-2 signaling and upregulation of nuclear factor-κB7,8. Interleukin 1β (IL-1β) plays a central role in the initiation of gouty inflammation9,10. MSU crystals, in the presence of IL-1β, stimulate cyclooxygenase-2 (COX-2) mRNA and prostaglandin E2 (PGE2) protein expression in human chondrocytes11, which may amplify inflammation. The direct effects of MSU crystals on chondrocyte viability and the ability of chondrocytes to maintain cartilage matrix have not yet been explored, and could be of significance to the overall function of cartilage in joints affected by gout.
The aim of our study was to investigate the mechanisms of cartilage damage in gout, through examination of the effects of MSU crystals on chondrocyte viability and function.
MATERIALS AND METHODS
Reagents
Unless specified, all reagents were purchased from Sigma-Aldrich. Culture media, fetal bovine serum (FBS), and antibiotics used in tissue culture were from Invitrogen (Life Technologies NZ Ltd.). Endotoxin-free MSU crystals were prepared by recrystallization from uric acid as described12.
Chondrocyte cell and cartilage organ culture
Cultures of primary human chondrocytes were prepared using macroscopically normal human cartilage obtained from 27 patients, aged 50–70 years old, undergoing knee or hip arthroplasty. Collection of human samples was approved by the Northern X Regional ethics committee. All patients provided written informed consent. For isolation of chondrocytes, macroscopically normal cartilage was removed from the bone and chopped finely. Tissue was incubated at 37°C with 0.8% pronase for 1 h, followed by 0.1% collagenase for up to 18 h until all tissue had been digested. Cells were then seeded in 75 cm2 flasks (Corning) with 10% FBS/Dulbecco modified Eagle’s medium (DMEM) containing 20 μg/ml L-ascorbic acid 2-phosphate (AA2P). Cells were cultured to 90% confluence and passaged once before use. For in vitro experiments, 48-well tissue culture plates (Corning) were coated with poly(2-hydroxyethyl methacrylate)(poly-HEMA) as described7, to prevent cellular attachment and maintain a chondrocytic phenotype under nonadherent conditions13. For cartilage organ culture, full-depth slices of cartilage were removed from the bone and cartilage explants (6 mm diameter) were punched out using a stainless steel biopsy punch. Explants were transferred to poly-HEMA coated 48-well tissue culture plates and were maintained in 10% FBS/DMEM/20 μg/ml AA2P for 2 days. All cultures contained serum and were incubated at 37°C with 5% CO2.
AlamarBlue assay for cell viability
Primary human chondrocytes were seeded into poly-HEMA coated 48-well plates at 300,000 cells/well in 1% FBS/DMEM/20 μg/ml AA2P. Primary human cartilage explants were cultured as described above. Twenty-four h after seeding, media were changed and MSU crystals were added for a further 24 h. Preliminary optimization experiments demonstrated that MSU crystals alone interfered with endpoint values for all assays assessed (data not shown)14. Therefore, 1–2 U/ml uricase was added to all cultures (including control wells) for the final 4 h of MSU crystal incubation plus an additional 18 h. Following dissolution of crystals, 5% alamarBlue was added for 18 h and fluorescence was read (excitation 540 nm; emission 630) using a Synergy 2 multi-detection microplate reader (BioTek Instruments Inc.) as described14. There were 2–4 wells in each treatment group. Explant viability was corrected for using the total tissue wet weight for each individual explant.
MTT assay for cell viability
Primary human chondrocytes were seeded into 24-well plates (Greiner Bio-One) at 37,500 cells/well in 5% FBS/DMEM/20 μg/ml AA2P. Cells were cultured with MSU crystals, uricase, and 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT; Invitrogen), and changes in cell viability were assessed as described14.
Flow cytometry
Primary human chondrocytes were seeded into 24-well plates at 50,000 cells/well in 5% FBS/DMEM/20 μg/ml AA2P. Fresh media and MSU crystals were then added for various times. Cells were treated with 2 U/ml uricase for the final 2 h of MSU culture and then stained with annexin V and propidium iodide (PI) as described14. Samples were analyzed on an LSRII flow cytometer using FACSDiva software (Becton Dickinson).
Quantitative real-time PCR
Primary human chondrocytes were seeded into 48-well poly-HEMA coated plates at 500,000 cells/well in 1% FBS/DMEM/20 μg/ml AA2P. The next day, fresh media and various concentrations of MSU crystals were added for 16 h. There were 3 wells per treatment. Cells were harvested and total cellular RNA was extracted using the TRIzol (Invitrogen) method15. Synthesis of cDNA and multiplex real-time PCR was performed as described16. The ΔΔCt method was used to quantify mRNA gene expression. Target gene expression was normalized to the expression of an endogenous control, 18S rRNA. This corrects for differences in cell number and cDNA input between samples. Target gene expression levels were calculated relative to the control (no MSU crystals).
Collagen deposition assay
Primary human chondrocytes were seeded into 24-well plates at 200,000 cells/well in 5% FBS/DMEM/20 μg/ml AA2P. After 2 days, media were changed and 30 ng/ml transforming growth factor-β was added to all wells for 2 h prior to the addition of MSU crystals for 5 days. Cells were washed and 1 U/ml uricase was added for 18 h to dissolve MSU crystals. Cells were then fixed using Bouin’s solution, washed with distilled water, dried, and stained with 0.1% Sirius red. Cells were washed with 0.01 M hydrochloric acid and Sirius red dye was released with 0.1 M sodium hydroxide before absorbance was read at 570 nm using a Synergy 2 multidetection microplate reader. There were 3–4 wells in each treatment group.
Histology of cartilage explants
Cartilage explants were cultured as described above. MSU crystals were added for 24 h and explants were then fixed in formalin, decalcified in 10% formic acid/5% formaldehyde, paraffin-embedded, sectioned through the sagittal plane at 5 μm intervals and placed on glass slides. Slides were processed and stained with toluidine blue as described14 and analyzed by light microscopy. The percentage of dead chondrocytes was determined by counting all dead and live cells up to a depth of 55 μm from the articular cartilage surface for each section of cartilage. Live chondrocytes were defined as cells within a lacuna that had an intact nucleus, while all empty lacunae and cells without an intact nucleus were counted as dead chondrocytes. All observations were made by 1 scorer (AC), who was blinded to patient data and treatment details. There were 2–3 explants for each experimental point, and up to 9 sections were counted for each explant. The mean number of cells for each explant was used for analysis.
Histology of joint samples from patients with gout
Cartilage samples (2 from first metatarsophalangeal joints and 1 each from a knee, midfoot, and finger distal interphalangeal joint) were obtained from 3 patients with gout undergoing orthopedic surgery and from 2 cadaveric donors with microscopically proven gout. Cadaveric samples were transferred to 70% ethanol at the time of harvest. Paraffin-embedded demineralized slides were prepared as described16. Sections were stained with 1% toluidine blue or with H&E, and examined using polarizing light microscopy. Cartilage morphology and its relationship to tophus were assessed by a musculoskeletal histopathologist (MD), and are presented as descriptive data only.
Statistical analysis
Data were analyzed using GraphPad Prism Software (GraphPad Software) or SAS Software (v9.2 SAS Institute Inc.). For all experiments, data were pooled from 3–6 biological repeats and are presented as mean and the standard error of the mean (SEM). Biological repeats of each assay were done using different patient samples. Data were analyzed using 1-way or 2-way ANOVA with posthoc Dunnett and Bonferroni multiple comparison tests, as appropriate.
RESULTS
MSU crystals reduced chondrocyte viability in a dose-dependent manner without altering levels of apoptosis
MSU crystals reduced human chondrocyte viability after 24 h in a dose-dependent manner in the alamarBlue assay (p < 0.0001; Figure 1A). The effects on chondrocyte viability were confirmed using different batches of MSU crystals synthesized in our laboratory and from another institution (gift of Professor Dorian Haskard, Hammersmith Hospital, UK; data not shown). The effects on viability were also confirmed using the MTT assay (data not shown). Further characterization of the chondrocyte-MSU crystal interaction demonstrated that this effect on viability was specific to the crystalline form of urate, because soluble uric acid had no significant effects on chondrocyte viability at the same concentrations used for MSU crystals (data not shown). The effects on chondrocyte viability did not vary with different MSU crystal lengths and were not inhibited by increasing levels of serum in cultures (data not shown). Flow cytometry showed there was no change in the percentage of early apoptotic cells (PI-negative and annexin V-positive) following culture of primary human chondrocytes with various concentrations of MSU crystals for 24 h (p = 0.588; Figure 1B). Further, there was no significant change in the percentage of early apoptotic cells following culture with 0.3 mg/ml MSU crystals for shorter time periods of culture (Figure 1C). However, there was a steady rise in the percentage of dead cells (PI-positive) and a steady decline in the percentage of live cells (annexin 5-negative and PI-negative) over time with MSU crystals, and over 60% of cells were dead (PI-positive) at the end of the 24-h incubation (Figure 1C).

Characteristics of the interaction between primary human chondrocytes and monosodium urate monohydrate (MSU) crystals. A. MSU crystals reduce chondrocyte cell viability in a dose-dependent manner after 24 h of culture as assessed using the alamarBlue assay. Data shown are pooled from 5 individual patient samples and are presented as mean (SEM) and 1-way ANOVA (p < 0.0001) with posthoc Dunnett test. *p < 0.01, **p < 0.001 vs control (no MSU crystals). B. MSU crystals rapidly induce cell death but not apoptosis in chondrocytes. Flow cytometry was used to determine changes in the percentage of dead [propidium iodide (PI)-positive], live (PI-negative, annexin V–negative), and apoptotic (PI-negative, annexin V–positive) chondrocytes following culture with various concentrations of MSU crystals for 24 h. C. 0.3 mg/ml MSU crystals over the course of 24 h. Data shown are pooled from 4 individual patient samples and are presented as mean (SEM), 1-way ANOVA (p value for each cell state shown in graph legends).
MSU crystals reduced the viability of chondrocytes within their extracellular matrix
In the alamarBlue assay, higher concentrations of MSU crystals (0.3 and 0.5 mg/ml) also reduced the viability of chondrocytes within human cartilage explants following 24 h of culture (p < 0.01; Figure 2A). To confirm these findings, we analyzed the histological appearance of primary human cartilage explants cultured with MSU crystals. This analysis demonstrated an increase in the percentage of dead cells in the superficial layer of cartilage cultured with 0.3 mg/ml MSU crystals for 24 h compared to control cartilage (p = 0.004; Figure 2B). Proteoglycan staining was not depleted compared to control (Figure 2C) and there was no change in the number of fissures or amount of surface fibrillation (data not shown).

Monosodium urate monohydrate (MSU) crystals reduce the viability of primary human chondrocytes within their extracellular matrix. A. The alamarBlue assay was used to assess the viability of primary human cartilage explants following culture with MSU crystals for 24 h. Viability was corrected for using the wet weight of each individual explant. Data shown are pooled from 5 individual patient samples and are presented as mean (SEM) and 1-way ANOVA (p < 0.01) with posthoc Dunnett test. * p < 0.05 vs control (no MSU crystals). B. Primary human cartilage explants were cultured with MSU crystals for 24 h and the number of live and dead cells in the articular superficial layer were counted following staining with toluidine blue. The percentage of dead cells with increasing concentrations of MSU crystals is shown. Data shown are pooled from 3 individual patient samples and are presented as mean SEM and 1-way ANOVA (p = 0.004) with posthoc Dunnett test. *p < 0.05 vs control (no MSU crystals). C. Representative photomicrographs of toluidine blue–stained cartilage cultured with or without 0.3 mg/ml MSU crystals for 24 h. Arrows indicate empty lacunae. Scale bar represents 100 μm.
MSU crystals altered mRNA gene expression levels and reduced collagen protein deposition in chondrocytes
Real-time PCR was used to determine changes in the relative mRNA expression levels of chondrocytic genes. Primary human chondrocytes cultured with 0.3 and 0.5 mg/ml MSU crystals for 16 h had significantly increased expression of the degradative enzymes ADAMTS4 and ADAMTS5 (p < 0.05 for both; Figure 3A–B). There was no change in the relative mRNA expression levels of matrix metalloprotease-13 (MMP-13; data not shown). Culture with MSU crystals decreased expression of the matrix proteins aggrecan and versican and collagen type 2α1 (p < 0.05 for all; Figure 3C–E). Sirius red staining demonstrated reduced collagen deposition by primary human chondrocytes following treatment with 0.3 and 0.5 mg/ml MSU crystals for 5 days (p < 0.0001; Figure 3F).
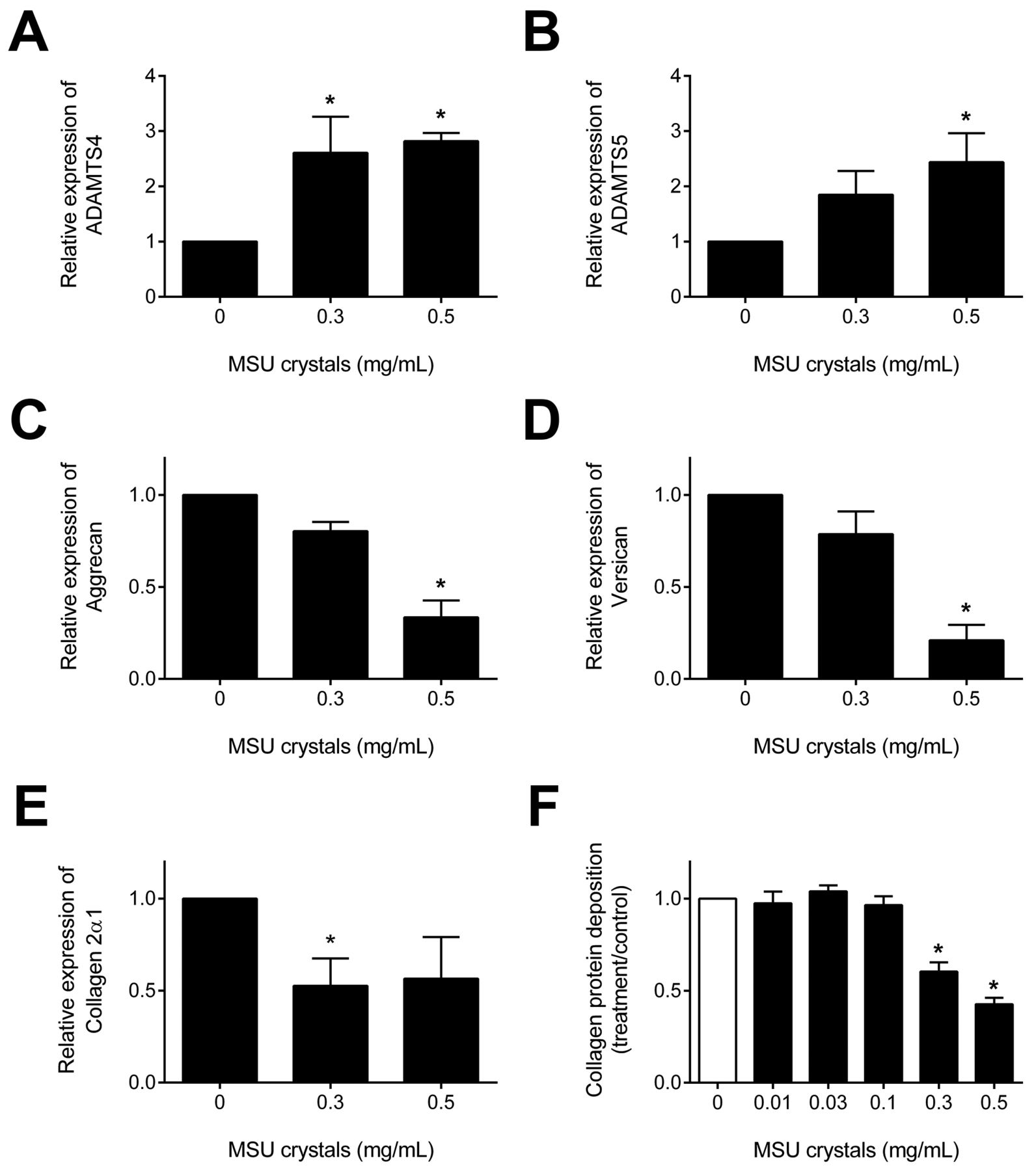
Monosodium urate monohydrate (MSU) crystals alter relative mRNA expression levels of chondrocytic genes and reduce collagen deposition in primary human chondrocytes. Real-time PCR was used to determine changes in the relative mRNA expression levels of the degradative enzymes ADAMTS4 and ADAMTS5 (A–B); matrix proteins aggrecan and versican (C–D), and collagen type 2α1 (E), in chondrocytes cultured with MSU crystals for 16 h. Data shown are presented as mean (SEM) and 1-way ANOVA (p < 0.05) with posthoc Dunnett’s test. *p < 0.05 vs control (no MSU crystals). F. Sirius red staining for collagen was used to determine changes in transforming growth factor-β–induced collagen deposition following treatment with MSU crystals for 5 days. Data shown are pooled from 4 individual patient samples and are presented as mean (SEM) and 1-way ANOVA (p < 0.0001) with posthoc Dunnett’s test. *p < 0.001 vs control (no MSU crystals).
Cartilage in joints affected by gout was highly disordered
The potential clinical relevance of the chondrocyte and cartilage explant assay results was further explored through examination of cartilage morphology in joints affected by gout. In these joints, cartilage adjacent to MSU crystals was highly disordered, with a loss of normal hyaline cartilage architecture, surface discontinuity, and empty chondrocyte lacunae. In particular, in joints with extensive tophi, only small residual pieces of degenerate cartilage were observed, almost entirely surrounded by MSU crystals, and with few or no live chondrocytes within lacunae. Proteoglycan staining was still present in these small fragments of cartilage (Figure 4).

Cartilage adjacent to monosodium urate monohydrate (MSU) crystals is highly disordered in patients with gout. Photomicrographs showing a cartilage fragment with empty chondrocyte lacunae, surrounded by MSU crystals and tophaceous material in the index finger distal interphalangeal joint of a cadaveric donor with microscopically proven gout. H&E–stained samples were viewed using (A) light microscopy and (B) a polarizing filter and red compensator. C. Toluidine blue–stained samples were viewed using light microscopy. Empty chondrocyte lacunae are marked with arrows.
DISCUSSION
Our study shows that MSU crystals interact with human chondrocytes to alter chondrocyte viability and function. The viability findings using isolated chondrocytes were confirmed in ex vivo cartilage explants, demonstrating that MSU crystals can influence chondrocytes within their extra-cellular matrix. The gene expression data suggest that there is an imbalance between the formation of new matrix components and increased activity of degradative enzymes in chondrocytes exposed to MSU crystals, leading to a compromised cartilage matrix. This imbalance, combined with a reduction in chondrocyte viability and overall function, may limit the ability of cartilage to effectively repair itself once damaged.
Previous histological case series of gout have described chondrocyte death within the cartilage of joints affected by gout17, as was observed in our study. The effects of MSU crystals on chondrocyte viability have not been reported, to our knowledge. Research has shown that soluble uric acid has no effect on primary rabbit chondrocyte viability18 and is able to inhibit IL-1-induced cell death in a chondrocytic cell line19. Our in vitro data demonstrating cell death when MSU crystals, but not soluble urate, were added to isolated chondrocytes support the notion that the crystalline form of urate is important in this process. We observed inhibitory effects on chondrocyte viability at very low concentrations of MSU crystals, in both isolated chondrocytes and ex vivo cartilage explant cultures. The mechanism by which MSU crystals cause chondrocyte death is not certain. The observation that annexin V binding in isolated chondrocytes did not change following culture with MSU crystals suggests that the early phase of the apoptotic process was not induced20, but does not provide definitive evidence that programmed cell death does not occur in response to MSU crystals in vivo. The effects of MSU crystals on isolated chondrocyte viability in vitro were rapid, suggesting a necrotic mechanism of cell death, most likely membranolysis following interaction of the crystal with the cell membrane. However, we also observed that culture with MSU crystals induced chondrocyte death in cells that were distant from the articular surface within cartilage explants. It has been postulated that the chondrocyte death observed histologically in gouty samples may be due to impaired cell nutrition, because MSU crystals coating the cartilage surface may adsorb to nutrients and/or hinder their diffusion through the matrix17,21,22. A lack of nutrients may help explain the chondrocyte cell death observed in the superficial layer in our histological analysis of cartilage explants cultured with MSU crystals, although this could not be directly assessed in our ex vivo models. The activation of intracellular signaling pathways leading to the production of soluble mediators such as nitric oxide may also contribute to chondrocyte death in response to MSU crystals8,19. Further work is needed to elucidate the mechanism by which MSU crystals reduce chondrocyte viability, particularly in those chondrocytes embedded within cartilage matrix.
Our study has also provided new insights into the mechanisms of cartilage degradation in gout. Our gene expression data suggest that MSU crystals reduce anabolic function and promote a catabolic state in chondrocytes. Importantly, the gene expression assays corrected for differences in cell number between samples through normalization to an endogenous control. Therefore, loss of cell viability does not explain the reduction in matrix protein gene expression. The reduction in collagen deposition using the Sirius red assay is consistent with the gene expression data, although it should be noted that collagen deposition may have been reduced by both inhibition of protein expression and loss of cell viability; cell number could not be determined as part of this assay. Earlier research from other groups has shown that MMP-3, COX-2, and nitric oxide synthesis are upregulated in chondrocytes cultured with MSU crystals7,8,11; factors that can actively cause matrix breakdown. We have extended these findings to show that the expression of essential cartilage matrix proteins and collagens is also reduced in the presence of MSU crystals, which may impair cartilage repair responses in joints affected by MSU crystal deposition.
A potential limitation in our study was that the human chondrocytes and cartilage used for experiments were taken from osteoarthritic joints. However, this model may be particularly relevant in the study of gout, noting the growing evidence to suggest a link between osteoarthritis (OA) and deposition of MSU crystals. OA and gout frequently coexist23,24 and synovial uric acid levels have been shown to be a marker of OA severity in the knee25. The direction of this association is unclear; that is, whether the presence of osteoarthritic cartilage predisposes to local formation and deposition of MSU crystals, or whether MSU crystals initiate and drive cartilage damage. A number of explanations have been suggested for why MSU crystals preferentially deposit in osteoarthritic cartilage26. The OA joint has increased levels of cartilage degradation products present such as chondroitin sulphate, which lowers urate solubility and promotes nucleation and growth of MSU crystals27,28,29. In addition, chondrocyte death within late-stage osteoarthritic cartilage30 leads to locally elevated concentrations of uric acid in the joint from the degradation of nucleic acids31. Our data suggest that, once present in osteoarthritic joints, MSU crystals promote further chondrocyte death and degradation of the cartilage matrix, possibly through the increased activity of degradative enzymes, which play a role in both arthropathies. ADAMTS4 and ADAMTS5 aggrecanases are highly expressed in osteoarthritic cartilage32,33 and have been shown to be important for driving cartilage damage in osteoarthritic joints34,35. Our results also demonstrated increased gene expression of ADAMTS4 and ADAMTS5 by chondrocytes cultured with MSU crystals, a finding that has not been previously reported, to our knowledge, implying some shared mechanisms of joint damage between OA and gout. This work also highlights the need for an in vivo model of chronic gout, which would enable the changes in cartilage and other joint structures to be observed and assessed over time.
Our study demonstrates that MSU crystals contribute to cartilage damage in gout through a reduction in chondrocyte viability and function, and increased catabolic activity within cartilage. Our data suggest that deposition of MSU crystals onto articular cartilage has important consequences for progressive joint disease in patients with gout. Thus, the “double contour” sign on ultrasonography may have important prognostic implications for progression of cartilage damage in gout. Longitudinal studies examining the relationship between the double contour sign and progression of cartilage loss will be of great interest. It is unknown whether dissolution of MSU crystals through effective urate-lowering therapy (ULT) can alter progression of radiographic damage, including joint space narrowing. These data provide a rationale for clinical trials that address the efficacy of ULT to prevent cartilage damage in patients with gout.
Acknowledgment
We acknowledge Sharita Meharry and Satya Amirapu for assisting with histological sample processing.
Footnotes
-
Supported by a University of Auckland Doctoral Scholarship (AC) and the Auckland Medical Research Foundation.
- Accepted for publication August 29, 2013.








{kind=link}
{kind=link}
{kind=link}
{kind=link}